Тело Барра

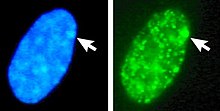
Тело Барра (названо в честь первооткрывателя Мюррея Барра ) [1] или Х-хроматин представляет собой неактивную Х-хромосому . У видов с определением пола XY (включая человека) самки обычно имеют две Х-хромосомы. [2] и человек становится неактивным в процессе, называемом лионизацией . Ошибки в разделении хромосом также могут привести к появлению у людей мужского и женского пола дополнительных Х-хромосом. Гипотеза Лиона утверждает, что в клетках с несколькими Х-хромосомами все, кроме одной, инактивированы на ранних стадиях эмбрионального развития млекопитающих . [3] [4] Х-хромосомы, которые становятся инактивированными, выбираются случайным образом, за исключением сумчатых и некоторых внеэмбриональных тканей некоторых плацентарных млекопитающих, у которых Х-хромосома из сперматозоидов всегда деактивирована. [5]
У людей с эуплоидией генотипическая ) женщина (46, XX кариотип имеет одно тельце Барра на ядро соматической клетки , а генотипический мужчина (46, XY) не имеет ни одного. Тельце Барра можно увидеть в интерфазном ядре как небольшую темную массу, контактирующую с мембраной ядра. Тельца Барра можно увидеть в нейтрофилах по краю ядра.
У людей с более чем одной Х-хромосомой количество телец Барра, видимых в интерфазе, всегда на единицу меньше общего числа Х-хромосом. Например, люди с синдромом Клайнфельтера (47, XXY) имеют одно тельце Барра, а люди с кариотипом 47, XXX — два тельца Барра.
Механизм
[ редактировать ]У человека с двумя Х-хромосомами (например, у большинства женщин приходится только одно тельце Барра ) на соматическую клетку , а у человека с одной Х-хромосомой (например, у большинства мужчин) его нет.
млекопитающих Инактивация Х-хромосомы инициируется из центра инактивации X или Xic , обычно расположенного вблизи центромеры . [6] Центр содержит двенадцать генов , семь из которых кодируют белки , пять — нетранслируемые РНК , из которых, как известно, только два играют активную роль в процессе инактивации X, Xist и Tsix . [6] Центр также, по-видимому, важен для подсчета хромосом: он гарантирует, что случайная инактивация происходит только тогда, когда присутствуют две или более Х-хромосомы. Предоставление дополнительного искусственного Xic на раннем эмбриогенезе может индуцировать инактивацию единственного X, обнаруженного в мужских клетках. [6]
Роли Ксиста и Тсикса кажутся антагонистическими. Потеря экспрессии Tsix на будущей неактивной Х-хромосоме приводит к увеличению уровней Xist вокруг Xic . Между тем, в дальнейшем активные уровни X Tsix сохраняются; таким образом, уровни Xist остаются низкими. [7] Этот сдвиг позволяет Xist начать покрывать будущую неактивную хромосому, распространяясь от Xic . [2] При неслучайной инактивации этот выбор, по-видимому, фиксирован, и современные данные позволяют предположить, что ген, унаследованный от матери, может быть импринтирован . [3] Сообщалось об изменениях частоты Xi в зависимости от возраста, беременности, использования пероральных контрацептивов, колебаний менструального цикла и неоплазии. [8]
Считается, что это представляет собой механизм выбора и позволяет последующим процессам установить компактное состояние тела Барра. Эти изменения включают модификации гистонов , такие как метилирование гистона H3 (т.е. H3K27me3 с помощью PRC2, который рекрутируется с помощью Xist ). [9] и убиквитинирование гистона H2A , [10] а также прямая модификация самой ДНК посредством метилирования сайтов CpG . [11] Эти изменения помогают инактивировать экспрессию генов на неактивной Х-хромосоме и вызвать ее уплотнение с образованием телец Барра.
Возможна также реактивация телец Барра, которая наблюдалась у больных раком молочной железы. [12] Одно исследование показало, что частота телец Барра при карциноме молочной железы была значительно ниже, чем у здоровых людей, что указывает на реактивацию этих когда-то инактивированных Х-хромосом. [12]
См. также
[ редактировать ]- Х-инактивация
- Система определения пола
- Ядерный секс — метод генетического определения пола.
- Деметилирование
- Ацетилирование
- Xist
- Тсикс (ген)
Ссылки
[ редактировать ]Ссылки на полные тексты статей предоставляются там, где доступ свободен, в остальных случаях дается ссылка только на аннотацию.
- ^ Барр, МЛ; Бертрам, Э.Г. (1949). «Морфологическое различие между нейронами мужского и женского пола и поведение ядрышкового сателлита во время ускоренного синтеза нуклеопротеинов». Природа . 163 (4148): 676–677. Бибкод : 1949Natur.163..676B . дои : 10.1038/163676a0 . ПМИД 18120749 . S2CID 4093883 .
- ^ Перейти обратно: а б Лион, МФ (2003). «Лион и гипотеза LINE». Семинары по клеточной биологии и биологии развития . 14 (6): 313–318. дои : 10.1016/j.semcdb.2003.09.015 . ПМИД 15015738 .
- ^ Перейти обратно: а б Браун, С.Дж., Робинсон, В.П. (1997), Экспрессия XIST и инактивация Х-хромосомы в предимплантационных эмбрионах человека Am. Дж. Хум. Жене. 61, 5–8 ( Полный текст в формате PDF )
- ^ Лион, МФ (1961). «Действие гена в Х -хромосоме мыши ( Mus musculus L.)». Природа . 190 (4773): 372–373. Бибкод : 1961Natur.190..372L . дои : 10.1038/190372a0 . ПМИД 13764598 . S2CID 4146768 .
- ^ Ли, Джей Ти (2003). «Инактивация Х-хромосомы: междисциплинарный подход». Семинары по клеточной биологии и биологии развития . 14 (6): 311–312. дои : 10.1016/j.semcdb.2003.09.025 . ПМИД 15015737 .
- ^ Перейти обратно: а б с Ружёль, К.; Авнер, П. (2003). «Контроль Х-инактивации у млекопитающих: что находится в центре?». Семинары по клеточной биологии и биологии развития . 14 (6): 331–340. дои : 10.1016/j.semcdb.2003.09.014 . ПМИД 15015740 .
- ^ Ли, Джей Ти; Давидов, Л.С.; Варшавский, Д. (1999). «Tisx, ген, антисмысловой по отношению к Xist в центре X-инактивации». Нат. Жене . 21 (4): 400–404. дои : 10.1038/7734 . ПМИД 10192391 . S2CID 30636065 .
- ^ Шарма, Дипти (10 января 2018 г.). «Расшифровка роли тельца Барра в злокачественных новообразованиях» . Медицинский журнал Университета Султана Кабуса . 17 (4): 389–397. дои : 10.18295/squmj.2017.17.04.003 . ПМК 5766293 . ПМИД 29372079 .
- ^ Херд, Э.; Ружёль, К.; Арно, Д.; Авнер, П.; Эллис, компакт-диск (2001). «Метилирование гистона H3 по Lys-9 является ранним признаком X-хромосомы во время инактивации X» . Клетка . 107 (6): 727–738. дои : 10.1016/S0092-8674(01)00598-0 . ПМИД 11747809 . S2CID 10124177 .
- ^ де Наполес, М.; Мермуд, JE; Вакао, Р.; Тан, Ю.А.; Эндо, М.; Аппана, Р.; Нестерова, Т.Б.; Сильва, Дж.; Отте, AP; Видаль, М.; Косеки, Х.; Брокдорф, Н. (2004). «Белки группы Polycomb связывают убиквитилирование гистона H2A Ring1A/B с наследственным подавлением генов и инактивацией X» . Дев. Клетка . 7 (5): 663–676. дои : 10.1016/j.devcel.2004.10.005 . ПМИД 15525528 .
- ^ Чедвик, BP; Уиллард, Х.Ф. (2003). «Запрет экспрессии генов после XIST: поддержание факультативного гетерохроматина на неактивном X». Семинары по клеточной биологии и биологии развития . 14 (6): 359–367. дои : 10.1016/j.semcdb.2003.09.016 . ПМИД 15015743 .
- ^ Перейти обратно: а б Натекар, Прашант Э.; ДеСуза, Фатима М. (2008). «Реактивация неактивной Х-хромосомы в буккальном мазке рака молочной железы» . Индийский журнал генетики человека . 14 (1): 7–8. дои : 10.4103/0971-6866.42320 . ISSN 0971-6866 . ПМК 2840782 . ПМИД 20300284 .
Дальнейшее чтение
[ редактировать ]- Альбертс, Б.; Джонсон, А.; Льюис, Дж.; Рафф, М.; Робертс, К.; Уолтер, П. (2002). Молекулярная биология клетки, четвертое издание . Гирляндная наука. стр. 428–429. ISBN 978-0-8153-4072-0 . (Веб-версия, бесплатный доступ)
- Тернпенни и Эллард: Элементы медицинской генетики Эмери 13E ( http://www.studentconsult.com/content/default.cfm?ISBN=9780702029172&ID=HC006029 . Архивировано 13 апреля 2020 г. в Wayback Machine ).