Фи Х 174
| Вирус эшерихии ΦX174 | |
|---|---|
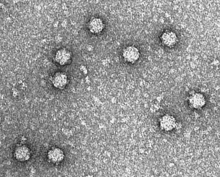 | |
| Электронная микрофотография фага ΦX174. | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Моноднавирия |
| Королевство: | Сангервиры |
| Тип: | Фиксвирикота |
| Сорт: | Мальграндавирицетес |
| Заказ: | Петитвирусы |
| Семья: | Микровирусиды |
| Род: | Вирус Зинсхаймера |
| Разновидность: | Вирус эшерихии ΦX174 |

Бактериофаг phi X 174 (или ΦX174 ) ) , представляет собой вирус с одноцепочечной ДНК ( оцДНК который инфицирует Escherichia coli . Этот вирус был выделен в 1935 году Николаем Булгаковым. [1] в Феликса д'Эреля лаборатории в Институте Пастера из образцов, собранных в канализации Парижа. Его характеристика и изучение механизма репликации проводились с 1950-х годов. Это был первый геном на основе ДНК , который удалось секвенировать. Эта работа была завершена Фредом Сэнгером и его командой в 1977 году. [2] В 1962 году Уолтер Фирс и Роберт Синшеймер уже продемонстрировали физическую, ковалентно замкнутую кольцевость ДНК ΦX174. [3] Лауреат Нобелевской премии Артур Корнберг использовал ΦX174 в качестве модели, чтобы впервые доказать, что ДНК, синтезированная в пробирке с помощью очищенных ферментов, может обладать всеми свойствами природного вируса, открывая эпоху синтетической биологии . [4] [5] В 1972–1974 годах Джерард Гурвиц , Сью Викнер и Рид Викнер с соавторами идентифицировали гены, необходимые для производства ферментов, катализирующих превращение одноцепочечной формы вируса в двухцепочечную репликативную форму. [6] сообщила В 2003 году группа Крейга Вентера , что геном ΦX174 был первым, который был полностью собран in vitro из синтезированных олигонуклеотидов. [7] Вирусная частица ΦX174 также была успешно собрана in vitro . [8] В 2012 году было показано, как его сильно перекрывающийся геном может быть полностью распакован и при этом оставаться функциональным. [9]
Геном
[ редактировать ]
Этот бактериофаг имеет [+] смысловой кольцевой одноцепочечный ДНК- геном длиной 5386 нуклеотидов . [10] в геноме Содержание GC составляет 44% и 95% нуклеотидов принадлежат кодирующим генам. Из-за сбалансированного характера оснований генома она используется в качестве контрольной ДНК для секвенаторов Illumina. [ нужна ссылка ]
Гены
[ редактировать ]ΦX174 кодирует 11 генов, названных последовательными буквами алфавита в том порядке, в котором они были обнаружены, за исключением A*, который является альтернативным стартовым кодоном в больших генах A. Только гены А* и К считаются несущественными, хотя относительно А* есть некоторые сомнения, поскольку его стартовый кодон можно заменить на АТТ, но не на какую-либо другую последовательность. [11] Теперь известно, что АТТ, вероятно, все еще способна производить белок. [12] внутри E. coli , и, следовательно, этот ген на самом деле может быть важным.
Первая половина генома ΦX174 характеризуется высоким уровнем перекрывания генов. [13] при этом восемь из 11 генов перекрываются хотя бы на один нуклеотид. [2] Было показано, что эти совпадения не являются существенными. [9] хотя реорганизованный фаг с удаленными всеми перекрытиями генов имел меньшую приспособленность по сравнению с диким типом. [14]
Фаг ΦX174 использовался, чтобы попытаться установить отсутствие неоткрытой генетической информации с помощью подхода «доказательство путем синтеза». [15]
Транскриптом
[ редактировать ]В 2020 году был создан транскриптом ΦX174. [16] Примечательными особенностями транскриптома ΦX174 является серия из четырех относительно слабых промоторов последовательно с четырьмя Rho-независимыми (внутренними) терминаторами и одним Rho-зависимым терминатором. [ нужна ссылка ]
Белки
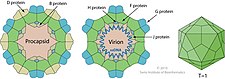
[ редактировать ]ΦX174 кодирует 11 белков .
| Белок | Копии | Функция [17] |
|---|---|---|
| А | — | Никсы RF ДНК для инициации репликации по катящемуся кругу ; лигирует концы линейной фаговой ДНК с образованием одноцепочечной кольцевой ДНК |
| А* | — | Ингибирует репликацию ДНК клетки-хозяина; блокирует суперинфицирующий фаг; не обязательно |
| Б | 60 в прокапсиде | Внутренний каркасный белок, участвующий в сборке прокапсида |
| С | — | Упаковка ДНК |
| Д | 240 в прокапсиде | Внешний каркасный белок, участвующий в сборке прокапсида |
| И | — | клеток-хозяев Лизис |
| Ф | 60 в вирионе | Основной капсидный белок |
| Г | 60 в вирионе | Главный спайковый белок |
| ЧАС | 12 в вирионе | Пилотный белок ДНК (или белок с второстепенным шипом) |
| Дж | 60 в вирионе | Связывается с новой одноцепочечной ДНК фага; сопровождает ДНК фага в прокапсид |
| К | — | Оптимизирует размер пакета; не обязательно |
Протеом
[ редактировать ]Недавно сообщалось об идентификации всех белков ΦX174 с помощью масс-спектрометрии. [14]
Инфекционный цикл
[ редактировать ]Инфекция начинается, когда G-белок связывается с липополисахаридами на поверхности бактериальной клетки-хозяина. Белок H (или пилотный белок ДНК) проводит вирусный геном через бактериальную мембрану E.coli . бактерий [18] скорее всего, через предсказанную спираль N-концевого трансмембранного домена . [19] Однако стало очевидно, что белок H является многофункциональным белком. [20] Это единственный вирусный капсидный белок ΦX174, у которого отсутствует кристаллическая структура по нескольким причинам. Он имеет низкое содержание ароматических соединений и высокое содержание глицина , что делает структуру белка очень гибкой, и, кроме того, отдельные атомы водорода (группа R для глицинов) трудно обнаружить при кристаллографии белка. Кроме того, белок H индуцирует лизис бактериального хозяина в высоких концентрациях, поскольку предсказанная N-концевая трансмембранная спираль легко протыкает дыры в бактериальной стенке. По данным биоинформатики , этот белок содержит четыре предсказанных спиральных домена, которые имеют значительную гомологию с известными факторами транскрипции. Кроме того, было установлено, что белок H de novo необходим для оптимального синтеза других вирусных белков. [21] Мутации белка H, которые препятствуют внедрению вируса, можно преодолеть, если ввести избыточное количество белка B, внутреннего каркасного белка. [ нужна ссылка ]
ДНК выбрасывается через гидрофильный канал в 5-кратной вершине. [22] Понятно, что белок H находится в этой области, но экспериментальные данные не подтвердили его точное местоположение. Попав внутрь бактерии-хозяина, репликация генома [+] оцДНК происходит через промежуточную ДНК с отрицательным смыслом . Это происходит по мере того, как фаговый геном скручивается, а вторичная структура, образованная в результате такой сверхспирализации, притягивает примосомный белковый комплекс. Он один раз транслоцируется по геному и синтезирует [-] оцДНК из положительного исходного генома. [+] Геномы оцДНК для упаковки в вирусы создаются на основе механизма вращающегося круга. Это механизм, с помощью которого двухцепочечный сверхспиральный геном разрывается на положительной цепи кодируемым вирусом белком А, что также привлекает бактериальную ДНК-полимеразу (ДНАП) к месту расщепления. DNAP использует отрицательную цепь в качестве матрицы для придания ДНК положительного смысла. Транслоцируясь по геному, он вытесняет внешнюю цепь уже синтезированной ДНК, которая немедленно покрывается белками SSBP . Белок А расщепляет весь геном каждый раз, когда распознает исходную последовательность. [ нужна ссылка ]
Поскольку белок D является наиболее распространенным транскриптом гена, его больше всего в вирусном прокапсиде. Аналогично, транскрипты генов F, J и G более распространены, чем H, поскольку стехиометрия этих структурных белков составляет 5:5:5:1. Примосомы представляют собой белковые комплексы, которые прикрепляют/связывают фермент геликазу с матрицей. Примосомы дают цепям РНК праймеры для синтеза ДНК. [ нужна ссылка ]
Филогенетика и разнообразие
[ редактировать ]ΦX174 тесно связан с другими микровирусами , особенно с фагом NC (например, NC1, NC7, NC11, NC16, NC37, NC5, NC41, NC56, NC51 и т. д.) и более отдаленно связан с G4-подобными фагами и еще более отдаленно связан с к α3-подобному фагу. Рокита и др. В 2006 г. было представлено филогенетическое древо их взаимоотношений. [23]
Использование
[ редактировать ]Экспериментальная эволюция
[ редактировать ]ΦX174 использовался в качестве модельного организма во многих эволюционных экспериментах. [24]
Биотехнология
[ редактировать ]ФХ174 регулярно используется в качестве положительного контроля при секвенировании ДНК из-за его относительно небольшого размера генома по сравнению с другими организмами, относительно сбалансированного содержания нуклеотидов — около 23% G, 22% C, 24% A и 31% T, т.е. 45% G+C и 55% A+T, см. инвентарь NC_001422.1. [10] из-за его последовательности длиной 5386 нуклеотидов. В приборах для секвенирования Illumina в качестве положительного контроля используется ΦX174. [25] и один цикл секвенирования Illumina может охватить геном ΦX174 несколько миллионов раз, что делает этот геном, весьма вероятно, наиболее тщательно секвенированным в истории. [ нужна ссылка ]
ΦX174 также используется для проверки устойчивости средств индивидуальной защиты к вирусам, передающимся через кровь. [26]
ΦX174 также был модифицирован для обеспечения пептидного дисплея (фагового дисплея) из белка вирусного капсида G. [27]
Синтетическая биология
[ редактировать ]Геном ΦX174 был первым фагом, клонированным в дрожжах. [9] что обеспечивает удобный сухой док для модификаций генома. [28] ΦX174 также был первым геномом, который был полностью распакован, и все перекрытия генов были удалены. [13] Эффект этих изменений привел к значительному снижению прикрепления к хозяину, нарушению регуляции экспрессии белка и чувствительности к теплу. [14]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Лакович, Здравко; Толян, Карло (20 декабря 2020 г.). «Владимир Сертич: забытый пионер вирусологии и бактериофаговой терапии» . Примечания и записи: Журнал Королевского общества истории науки . 74 (4): 567–578. дои : 10.1098/rsnr.2019.0010 . ISSN 0035-9149 . ПМЦ 7653334 . ПМИД 33177747 .
- ^ Jump up to: а б Сэнгер Ф., Air GM, Баррелл Б.Г., Браун Н.Л., Коулсон А.Р., Фиддес К.А. и др. (февраль 1977 г.). «Нуклеотидная последовательность ДНК бактериофага phi X174». Природа . 265 (5596): 687–95. Бибкод : 1977Natur.265..687S . дои : 10.1038/265687a0 . ПМИД 870828 . S2CID 4206886 .
- ^ Фирс В., Зиншаймер Р.Л. (октябрь 1962 г.). «Структура ДНК бактериофага фи-X174. III. Ультрацентрифужное доказательство кольцевой структуры». Журнал молекулярной биологии . 5 (4): 424–34. дои : 10.1016/S0022-2836(62)80031-X . ПМИД 13945085 .
- ^ Национальная библиотека медицинских профилей в области науки. Документы Артура Корнберга. «Создание жизни в пробирке», 1959–1970 гг. связь [ нужен неосновной источник ]
- ^ Гулиан М., Корнберг А., Зиншаймер Р.Л. (декабрь 1967 г.). «Ферментативный синтез ДНК XXIV. Синтез ДНК инфекционного фага фи-Х174» . Труды Национальной академии наук Соединенных Штатов Америки . 58 (6): 2321–8. Бибкод : 1967PNAS...58.2321G . дои : 10.1073/pnas.58.6.2321 . JSTOR 58720 . ПМК 223838 . ПМИД 4873588 .
- ^ Викнер С., Гурвиц Дж. (октябрь 1974 г.). «Преобразование вирусной ДНК phiX174 в двухцепочечную форму очищенными белками Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (10): 4120–4. дои : 10.1073/pnas.71.10.4120 . ПМК 434340 . ПМИД 4610569 .
- ^ Смит Х.О., Хатчисон К.А., Пфанкох К., Вентер Дж.К. (декабрь 2003 г.). «Создание синтетического генома путем сборки всего генома: бактериофаг phiX174 из синтетических олигонуклеотидов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (26): 15440–5. Бибкод : 2003PNAS..10015440S . дои : 10.1073/pnas.2237126100 . JSTOR 3149024 . ПМК 307586 . ПМИД 14657399 .
- ^ Черва Дж.Э., Органтини Л.Дж., Эшли Р.Э., Хафенштейн С.Л., Фейн Б.А. (сентябрь 2011 г.). «СБОРКА прокапсида øX174 in VITRO из олигомеров внешнего каркасного белка и ранних промежуточных соединений пентамерной сборки». Журнал молекулярной биологии . 412 (3): 387–96. дои : 10.1016/j.jmb.2011.07.070 . ПМИД 21840317 .
- ^ Jump up to: а б с Яшке П.Р., Либерман Э.К., Родригес Дж., Сьерра А., Энди Д. (декабрь 2012 г.). «Полностью декомпрессированный синтетический геном бактериофага øX174, собранный и заархивированный в дрожжах» . Вирусология . 434 (2): 278–84. дои : 10.1016/j.virol.2012.09.020 . ПМИД 23079106 .
- ^ Jump up to: а б с Фаг энтеробактерий phiX174 sensu lato , полный геном. «Полный геном: доступ NC_001422» , Национальный центр биотехнологической информации . Проверено 30 января 2016 г.
- ^ Баас П.Д., Ливеринк Х., ван Теффелен Х.А., ван Мансфельд А.Д., ван Бум Дж.Х., Янс Х.С. (июнь 1987 г.). «Изменение стартового кодона ATG белка A бактериофага phi X174 на кодон АТТ дает жизнеспособный фаг, что указывает на то, что белок A не важен для размножения phi X174» . Письма ФЭБС . 218 (1): 119–25. дои : 10.1016/0014-5793(87)81030-x . ПМИД 2954853 . S2CID 24174007 .
- ^ Хехт А., Глазго Дж., Яшке П.Р., Бавазер Л.А., Мансон М.С., Кокран Дж.Р. и др. (апрель 2017 г.). «Измерения инициации трансляции всех 64 кодонов E. coli» . Исследования нуклеиновых кислот . 45 (7): 3615–3626. дои : 10.1093/nar/gkx070 . ПМК 5397182 . ПМИД 28334756 .
- ^ Jump up to: а б Райт, Брэдли В.; Моллой, Марк П.; Яшке, Пол Р. (5 октября 2021 г.). «Перекрывающиеся гены в природных и искусственно созданных геномах» . Обзоры природы Генетика . 23 (3): 154–168. дои : 10.1038/s41576-021-00417-w . ISSN 1471-0064 . ПМЦ 8490965 . ПМИД 34611352 .
- ^ Jump up to: а б с Райт Б.В., Руан Дж., Моллой, член парламента, Яшке П.Р. (ноябрь 2020 г.). «Модуляризация генома показывает, что перекрывающаяся топология генов необходима для эффективного воспроизводства вирусов». ACS Синтетическая биология . 9 (11): 3079–3090. doi : 10.1021/acsynbio.0c00323 . ПМИД 33044064 . S2CID 222300240 .
- ^ Яшке П.Р., Дотсон Г.А., Хунг К.С., Лю Д., Энди Д. (ноябрь 2019 г.). «Окончательная демонстрация путем синтеза полноты аннотации генома» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (48): 24206–24213. дои : 10.1073/pnas.1905990116 . ПМК 6883844 . ПМИД 31719208 .
- ^ Логель Д.Ю., Яшке П.Р. (август 2020 г.). «Карта транскрипции бактериофага φX174 высокого разрешения» . Вирусология . 547 : 47–56. дои : 10.1016/j.virol.2020.05.008 . ПМИД 32560904 . S2CID 219459208 .
- ^ Фейн Б.А., Брентлингер К.Л., Берч А.Д., Чен М., Хафенштейн С., Мур Э., Новак Ч.Р., Утияма А. (2006). «ɸX174 и др., Микровирусы ». В Календаре R (ред.). Бактериофаги (2-е изд.). Нью-Йорк: Оксфордский университет. Нажимать. п. 130. ИСБН 978-0195148503 .
- ^ Язвинский С.М., Линдберг А.А., Корнберг А. (июль 1975 г.). «Липополисахаридный рецептор бактериофага phiX174 и S13». Вирусология . 66 (1): 268–82. дои : 10.1016/0042-6822(75)90197-х . ПМИД 1094681 .
- ^ Туснади Г.Е., Симон I (сентябрь 2001 г.). «Сервер прогнозирования трансмембранной топологии HMMTOP» . Биоинформатика . 17 (9): 849–50. дои : 10.1093/биоинформатика/17.9.849 . ПМИД 11590105 .
- ^ Черва Дж. Э., Янг Л. Н., Фейн Б. А. (март 2011 г.). «Развязка функций многофункционального белка: выделение мутанта пилотного белка ДНК, влияющего на морфогенез частиц» . Вирусология . 411 (1): 9–14. дои : 10.1016/j.virol.2010.12.026 . ПМИД 21227478 .
- ^ Рубоянес М.В., Чен М., Дубрава М.С., Черва Дж.Э., Фейн Б.А. (октябрь 2009 г.). «Экспрессия пилотных белков ДНК с делецией N-конца ингибирует ранние стадии репликации phiX174» . Журнал вирусологии . 83 (19): 9952–6. дои : 10.1128/JVI.01077-09 . ПМК 2748053 . ПМИД 19640994 .
- ^ Маккенна Р., Ся Д., Виллингманн П., Илаг Л.Л., Кришнасвами С., Россманн М.Г. и др. (январь 1992 г.). «Атомная структура одноцепочечной ДНК бактериофага phi X174 и ее функциональное значение» . Природа . 355 (6356): 137–43. Бибкод : 1992Natur.355..137M . дои : 10.1038/355137a0 . ПМК 4167681 . ПМИД 1370343 .
- ^ Рокита Д.Р., Берч К.Л., Кодл С.Б., Вичман Х.А. (февраль 2006 г.). «Горизонтальный перенос генов и эволюция геномов колифагов микровиридов» . Журнал бактериологии . 188 (3): 1134–42. дои : 10.1128/JB.188.3.1134-1142.2006 . ПМЦ 1347346 . ПМИД 16428417 .
- ^ Вичман Х.А., Браун С.Дж. (август 2010 г.). «Экспериментальная эволюция вирусов: микровирусы как модельная система» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 365 (1552): 2495–501. дои : 10.1098/rstb.2010.0053 . ПМЦ 2935103 . ПМИД 20643739 .
- ^ «Использование элемента управления PhiX для циклов секвенирования HiSeq®» . Иллюмина. Архивировано из оригинала 9 января 2019 года . Проверено 8 января 2019 г.
- ^ «СИЗ-Информация – Стандартные сведения» . wwwn.cdc.gov . Проверено 8 февраля 2019 г.
- ^ Кристакос К.Дж., Чепмен Дж.А., Фейн Б.А., Кампос С.К. (январь 2016 г.). «PhiXing-it, отображение чужеродных пептидов на бактериофаге ΦX174» . Вирусология . 488 : 242–8. дои : 10.1016/j.virol.2015.11.021 . ПМК 6191337 . ПМИД 26655242 .
- ^ Андо Х., Лемир С., Пирес Д.П., Лу Т.К. (сентябрь 2015 г.). «Разработка модульных вирусных каркасов для целевого редактирования бактериальной популяции» . Клеточные системы . 1 (3): 187–196. дои : 10.1016/j.cels.2015.08.013 . ПМЦ 4785837 . ПМИД 26973885 .
Внешние ссылки
[ редактировать ]- Гудселл Д (февраль 2000 г.). «Бактериофаг phiX174» . Молекула месяца . РЦББ-ПДБ.