KcsA калиевый канал
| KcsA калиевый канал | |||
|---|---|---|---|
 Четыре субъединицы, образующие канал, нарисованы разными цветами. Они окружают центральную пору, охраняемую селективным фильтром, состоящим из P-петлей каждой из субъединиц. Синие и красные точки обозначают границы липидного бислоя . | |||
| Идентификаторы | |||
| Символ | ? | ||
| Пфам | PF07885 | ||
| ИнтерПро | ИПР013099 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1бл8 / СКОПе / СУПФАМ | ||
| Суперсемейство OPM | 8 | ||
| белок OPM | 1r3j | ||
| |||
KcsA (K-канал стрептомицетов А) представляет собой прокариотический калиевый канал почвенной бактерии Streptomyces lividans , который широко изучался в ионных каналов исследованиях . Белок, активируемый рН, имеет два трансмембранных сегмента и высокоселективную область пор, ответственную за шлюзование и передвижение K. + ионы выходят из клетки. [1] [2] Аминокислотная последовательность, обнаруженная в селективном фильтре KcsA, высоко консервативна как среди прокариотических, так и среди эукариотических K. + каналы напряжения; [1] [3] в результате исследование KcsA предоставило важное структурное и механистическое понимание молекулярной основы K. + отбор и проводимость ионов. Будучи одним из наиболее изученных ионных каналов по сей день, KcsA является образцом для исследования K + Функция канала и ее выясненная структура лежат в основе компьютерного моделирования динамики каналов как для прокариотических, так и для эукариотических видов. [4]
История
[ редактировать ]KcsA был первым каналом ионов калия, который был охарактеризован с помощью рентгеновской кристаллографии Родериком Маккинноном и его коллегами в 1998 году. За годы, предшествовавшие этому, исследования структуры K + каналы были сосредоточены на использовании связывания небольших токсинов для выявления местоположения пор и селективного фильтра среди остатков каналов. Группа Маккиннона выдвинула теорию тетрамерного расположения трансмембранных сегментов и даже предположила наличие порообразующих «петлей» в области фильтра, состоящих из коротких сегментов аминокислот, которые взаимодействовали с K. + ионы, проходящие через канал [5] Открытие сильной гомологии последовательностей между KcsA и другими каналами семейства Kv, включая белок Shaker , привлекло внимание научного сообщества, особенно потому, что K + последовательность сигнатур каналов начала появляться в других прокариотических генах. Простота двух трансмембранных спиралей в KcsA, в отличие от шести во многих ионных каналах эукариот , также позволила понять механизмы KcsA. + направляет проводимость на более элементарном уровне, тем самым обеспечивая еще больший стимул для изучения KcsA.
Кристаллическая структура KcsA была расшифрована группой Маккиннона в 1998 году после открытия, что удаление C-концевого цитоплазматического домена нативного белка (остатки 126–158) увеличивает стабильность кристаллизованных образцов. Была создана модель KcsA с разрешением 3,2А, которая подтвердила тетрамерное расположение белка вокруг центральной поры, при этом одна спираль каждой субъединицы обращена к внутренней оси, а другая - наружу. [6] Три года спустя Мораис-Кабрал и Чжоу создали модель с более высоким разрешением после того, как моноклональные фрагменты Fab были прикреплены к кристаллам KcsA для дальнейшей стабилизации канала. [7] В начале 2000-х годов появились свидетельства занятия селективного фильтра двумя K + атом в процессе транспортировки возник на основе энергетических и электростатических расчетов, выполненных для моделирования области пор. Продолжающееся исследование различных открытых и закрытых, неактивных и активных конформаций KcsA с помощью других методов визуализации, таких как осс-ЯМР и ЭПР, с тех пор позволило еще лучше понять структуру каналов и силы, вызывающие переключение от инактивации канала к проводимости.
В 2007 году Риек и др. показали, что открытие канала, возникающее в результате титрования ионного канала от pH 7 до pH 4, соответствует конформационным изменениям в двух областях: переходу в ионообменное состояние селективного фильтра и открытию расположения ТМ2 при С -конечная точка . [8] Эта модель объясняет способность KcsA одновременно выбирать K + ионы, а также регулируют электропроводность. В 2011 году кристаллическая структура полноразмерного KcsA была определена и показала, что препятствия со стороны ранее усеченных остатков позволяют только прямое расширение области прохождения межклеточных ионов белка. Это исследование позволяет более детально рассмотреть движение отдельных областей каналов во время ионной проводимости. [9] В настоящее время исследования KcsA сосредоточены на использовании прокариотического канала в качестве модели динамики каналов более крупных эукариотических K. + каналы, включая hERG .
Структура
[ редактировать ]
Структура KcsA представляет собой перевернутый конус с центральной порой, идущей по центру и состоящей из двух трансмембранных спиралей (внешняя спираль M1 и внутренняя спираль M2), которые охватывают липидный бислой . Сам канал представляет собой тетрамер, состоящий из четырех идентичных однодоменных субъединиц (каждая с двумя α-спиралями), расположенных так, что одна спираль M2 обращена к центральной поре, а другая спираль M1 обращена к липидной мембране . Внутренние спирали наклонены примерно на 25° по отношению к липидной мембране и слегка изломаны, открываясь наружу, как цветок. [6] Эти две спирали ТМ связаны возвратной петлей, распределенной симметрично вокруг общей оси, соответствующей центральной поре . Область пор охватывает примерно 30 аминокислотных остатков и может быть разделена на три части: селективный фильтр вблизи внеклеточной стороны, расширенную, заполненную водой полость в центре и закрытые ворота вблизи цитоплазматической стороны, образованные четырьмя упакованными спиралями М2. [6] Обнаружено, что эта архитектура высококонсервативна в семействе калиевых каналов. [10] [11] как у эукариот , так и у прокариот.
Общая длина поры составляет 45 Å, а ее диаметр значительно варьируется в пределах отдельных участков внутреннего туннеля. Двигаясь из внутриклеточной области наружу (на рисунке снизу вверх), пора начинается с воротной области, образованной спиралями М2 диаметром 18 Å, а затем открывается в широкую полость (~ 10 Å в поперечнике) вблизи середины мембраны. . [6] В этих регионах К. + ионы контактируют с окружающими молекулами воды, но когда они попадают в канал из селективного фильтра вверху, полость настолько узкая, что K + ионы должны выделять гидратирующую воду, чтобы проникнуть в клетку. [6] Что касается аминокислотного состава остатков, выстилающих поры в KcsA, боковые цепи, выстилающие внутреннюю пору и полость, преимущественно гидрофобны , но в селективном фильтре присутствуют полярные аминокислоты, которые контактируют с обезвоженным K. + ионы.
Селективный фильтр
[ редактировать ]Более широкий конец конуса соответствует внеклеточному устью канала, состоящего из спиралей пор, а также селективного фильтра , образованного последовательностью TVGYG (треонин, валин, глицин, тирозин, глицин), характерной для калиевых каналов. [12] В этой области координация между аминокислотами TVGYG и поступающими K + ионы обеспечивают проводимость ионов через канал. Селективный фильтр KcsA содержит четыре сайта связывания ионов, хотя предполагается, что только два из этих четырех положений заняты одновременно. Селективный фильтр имеет диаметр около 3 Å. [13] хотя моделирование молекулярной динамики предполагает, что фильтр является гибким. [14] Присутствие TVGYG в области фильтра KcsA сохраняется даже в более сложных эукариотических каналах, что делает KcsA оптимальной системой для изучения KcsA. + проводимость канала у разных видов.
Функция
[ редактировать ]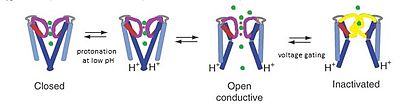
Канал KcsA считается модельным каналом , поскольку структура KcsA обеспечивает основу для понимания KcsA. + канальная проводимость, которая состоит из трех частей: по калию селективность , блокирование каналов за счет чувствительности к pH и потенциалзависимая инактивация каналов. К + ионов Проникновение происходит в верхней области селективного фильтра поры, в то время как pH-регулирование повышается в результате протонирования трансмембранных спиралей в конце поры. При низком pH спираль М2 протонируется, переводя ионный канал из закрытой в открытую конформацию. [15] Считается, что когда ионы проходят через канал, механизмы ограничения напряжения вызывают взаимодействия между Glu71 и Asp80 в селективном фильтре, которые дестабилизируют проводящую конформацию и облегчают переход в долгоживущее непроводящее состояние, напоминающее инактивацию напряжения C-типа . зависимые каналы . [16]
В непроводящей конформации KcsA при pH 7 K + тесно связан с координирующими атомами кислорода селективного фильтра, а четыре спирали TM2 сходятся возле цитоплазматического соединения, блокируя прохождение любых ионов калия. [8] Однако при pH 4 KcsA претерпевает конформационные обмены в миллисекундном масштабе, фильтруя проникающие и непроницаемые состояния, а также между открытой и закрытой конформациями спиралей M2. [8] Хотя эти отдельные конформационные изменения происходят в отдельных областях канала, молекулярное поведение каждой области связано как электростатическими взаимодействиями , так и аллостерией . [8] Динамика этого обмена стереохимических конфигураций в фильтре обеспечивает физическую основу для одновременного K + проводимость и стробирование.
К + избирательность
[ редактировать ]Последовательность TVGYG особенно важна для поддержания калиевой специфичности KcsA. Глицины в этой последовательности селективного фильтра имеют двугранные углы, которые позволяют карбонильным атомам кислорода в белковой основной цепи фильтра указывать в одном направлении, к ионам вдоль поры. [5] Глицины и треонин координируются с K + ион, в то время как боковые цепи валина и тирозина направлены в ядро белка, чтобы наложить геометрические ограничения на фильтр. В результате тетрамер KcsA содержит четыре равноотстоящих друг от друга K + места связывания, каждая сторона которого состоит из клетки, образованной восемью атомами кислорода, расположенными в вершинах куба. Атомы кислорода, окружающие K + ионы в фильтре располагаются подобно молекулам воды, окружающим гидратированный K + ионы в полости канала; это говорит о том, что центры координации и связывания кислорода в селективном фильтре оплачивают энергетические затраты K + обезвоживание. [5] Поскольку ион Na+ слишком мал для этих K + Поскольку места связывания имеют малый размер, энергия дегидратации не компенсируется и, таким образом, фильтр отбирает другие посторонние ионы. [5] Кроме того, канал KcsA блокируется Cs. + ионы и стробирование требуют присутствия Mg 2+ ионы. [1]
Чувствительность к pH
[ редактировать ]Зависимая от pH проводимость KcsA указывает на то, что открытие ионного канала происходит, когда белок подвергается воздействию более кислой среды. Исследования ЯМР, проведенные группой Riek, показывают, что чувствительность к pH возникает как в C-концевой области TM2 белка, так и в остатках Tyr78 и Gly79 в селективном фильтре. Есть данные, позволяющие предположить, что основной сенсор pH находится в цитоплазматическом домене. Замена отрицательно заряженных аминокислот на нейтральные делала канал KcsA нечувствительным к pH, хотя аминокислотных изменений в трансмембранной области не было. [17] [18] Кроме того, при pH от 6 до 7 гистидин является одной из немногих титруемых боковых цепей гистидинов; они отсутствуют в трансмембранных и внеклеточных сегментах TM2, но присутствуют на C-конце KcsA. Это подчеркивает возможный механизм медленного открытия KcsA, который особенно чувствителен к pH, особенно потому, что конформационное распространение сигнала открытия канала от С-конца к селективному фильтру может быть важным для координации структурных изменений, необходимых для проводимости вдоль всей поры. .
Исследования ЯМР также показывают, что сложная сеть водородных связей между Tyr78, Gly79, Glu71 и Asp80 существует в области фильтра KcsA и, кроме того, действует как pH-чувствительный триггер проводимости. Мутация ключевых остатков в этом регионе, включая E71A, приводит к большим энергетическим затратам – 4 ккал/моль. −1 , что эквивалентно потере водородной связи между Glu71 и Tyr78 и опосредованной водой водородной связи между Glu71 и Asp80 в KcsA(E71A). Эти исследования дополнительно подчеркивают роль шлюзования pH в функции каналов KcsA.
стробирование напряжения
[ редактировать ]В 2006 году группа Перозо предложила механистическое объяснение влияния полей напряжения на затвор KcsA. После подачи деполяризующего тока в канал происходит переориентация Glu71 в сторону внутриклеточной поры, тем самым разрушая карбоксил-карбоксилатную пару Glu71-Asp80, первоначально стабилизирующую селективный фильтр. Коллапс области фильтра предотвращает вход в инактивированное состояние или облегчает выход из него. [16] Glu71, ключевая часть сигнатурной последовательности фильтра селективности, которая консервативна среди K + ионные каналы играют ключевую роль в стробировании, поскольку его способность переориентироваться в направлении трансмембранного поля напряжения может дать объяснение событиям стробирования напряжения при KcsA. Ориентация аминокислот в области фильтра может играть важную физиологическую роль в модуляции потоков калия у эукариот и прокариот в стационарных условиях. [16]
Исследовать
[ редактировать ]Функция
[ редактировать ]Точный механизм селективности калиевых каналов продолжает изучаться и обсуждаться , и для описания различных аспектов селективности используется множество моделей. Модели, объясняющие селективность, на основе концепции напряженности поля, разработанной Джорджем Эйзенманом. [19] основанные на законе Кулона, были применены к KcsA. [14] [20] Альтернативное объяснение селективности KcsA основано на модели близкого соответствия (также известной как модель плотного прилегания), разработанной Франсиско Безанильей и Армстронгом . [21] Атомы кислорода основной карбонильной группы , составляющие селективный фильтр, удерживаются в точном положении, которое позволяет им замещать молекулы воды в гидратной оболочке иона калия , но они находятся слишком далеко от иона натрия . В дальнейшей работе были изучены термодинамические различия в связывании ионов. [22] топологические соображения, [23] [24] и количество непрерывных мест связывания ионов. [25]
Кроме того, еще предстоит обсудить главное ограничение в изучении и моделировании кристаллической структуры : лучше всего разрешенная и наиболее применяемая кристаллическая структура KcsA, по-видимому, представляет собой «закрытую» форму канала. Это разумно, поскольку закрытому состоянию канала благоприятствует нейтральный pH , при котором кристаллическая структура была определена с помощью рентгеновской кристаллографии . Однако динамическое поведение KcsA затрудняет анализ канала, поскольку кристаллическая структура неизбежно обеспечивает статическое, усредненное по пространству и времени изображение канала. Чтобы преодолеть разрыв между молекулярной структурой и физиологическим поведением, необходимо понимание динамики атомного разрешения калиевых каналов.
Приложения
[ редактировать ]Из-за высокого сходства последовательностей пор KcsA и других эукариотических K + белков ионных каналов, KcsA предоставил важную информацию о поведении других важных белков, проводящих напряжение, таких как дрозофиллы полученный из шейкер, человека , и калиевый канал hERG . KcsA использовался в исследованиях мутагенеза для моделирования взаимодействия hERG с различными лекарственными соединениями. Такие тесты могут выявить взаимодействие лекарственного средства с каналами hERG, вызывающее приобретенный синдром удлиненного интервала QT , и необходимы для определения сердечной безопасности новых лекарств. [26] Кроме того, с помощью вычислений были созданы модели гомологии, основанные на кристаллической структуре KcsA в закрытом состоянии, для построения представления сердечного K hERG в нескольких состояниях. + канал. Такие модели раскрывают гибкость канала hERG и могут последовательно предсказывать аффинность связывания набора разнообразных лигандов, взаимодействующих с ионными каналами. Анализ сложных структур лиганд-hERG можно использовать для управления синтезом аналогов лекарств с пониженной ответственностью hERG на основе структуры лекарства и потенциала докинга. [27]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Шремпф Х., Шмидт О., Кюммерлен Р., Хинна С., Мюллер Д., Бетцлер М., Стейнкамп Т., Вагнер Р. (ноябрь 1995 г.). «Прокариотический калиевый ионный канал с двумя предсказанными трансмембранными сегментами Streptomyces lividans» . Журнал ЭМБО . 14 (21): 5170–8. дои : 10.1002/j.1460-2075.1995.tb00201.x . ПМЦ 394625 . ПМИД 7489706 .
- ^ Мейзер Д., Сплитт Х., Вагнер Р., Шремпф Х. (1999). «Исследование открытой поры калиевого канала Streptomyces lividans » . Письма ФЭБС . 462 (3): 447–452. Бибкод : 1999FEBSL.462..447M . дои : 10.1016/S0014-5793(99)01579-3 . ПМИД 10622743 . S2CID 6231397 .
- ^ Ю Ф.Х., Яров-Яровой В., Гутман Г.А., Каттералл В.А. (декабрь 2005 г.). «Обзор молекулярных взаимоотношений в суперсемействе потенциал-управляемых ионных каналов». Фармакологические обзоры . 57 (4): 387–95. дои : 10.1124/пр.57.4.13 . ПМИД 16382097 . S2CID 2643413 .
- ^ Ру Б (2005). «Ионная проводимость и селективность в K (+) каналах». Ежегодный обзор биофизики и биомолекулярной структуры . 34 : 153–71. doi : 10.1146/annurev.biophys.34.040204.144655 . ПМИД 15869387 .
- ^ Jump up to: а б с д Родерик Маккиннон. «Нобелевская лекция: Калиевые каналы и атомная основа избирательной ионной проводимости» . Нобелевская премия . Нобель Медиа АБ.
- ^ Jump up to: а б с д и Дойл Д.А., Мораис Кабрал Дж., Пфюцнер Р.А., Куо А., Гулбис Дж.М., Коэн С.Л., Хаит Б.Т., Маккиннон Р. (апрель 1998 г.). «Строение калиевого канала: молекулярная основа К + проводимость и селективность». Science . 280 (5360): 69–77. Bibcode : 1998Sci...280...69D . doi : 10.1126/science.280.5360.69 . PMID 9525859 .
- ^ Чжоу Ю., Мораис-Кабрал Дж. Х., Кауфман А., Маккиннон Р. (ноябрь 2001 г.). «Химия координации и гидратации ионов, выявленная K + Комплекс канал-Fab с разрешением 2,0 А". Nature . 414 (6859): 43–8. : 2001Natur.414 ...43Z . doi : 10.1038/35102009 . PMID 11689936. S2CID Bibcode 205022645 .
- ^ Jump up to: а б с д Бейкер К.А., Цицилонис С., Квятковски В., Чой С., Рик Р. (ноябрь 2007 г.). «Конформационная динамика калиевого канала KcsA определяет воротные свойства» . Структурная и молекулярная биология природы . 14 (11): 1089–95. дои : 10.1038/nsmb1311 . ПМЦ 3525321 . ПМИД 17922011 .
- ^ Уйсал С., Куэлло Л.Г., Кортес Д.М., Койде С., Косьяков А.А., Перозо Э. (июль 2011 г.). «Механизм активационного стробирования в полноразмерном KcsA K + канал» . Sciences of the United States of America . 108 (29): 11896–9. Bibcode : 2011PNAS..10811896U . doi : 10.1073/pnas.1105112108 . PMC 3141920. . PMID 21730186 Proceedings of the National Academy of
- ^ Лу З., Клем А.М., Раму Ю. (октябрь 2001 г.). «Пора ионной проводимости сохраняется среди калиевых каналов». Природа . 413 (6858): 809–13. Бибкод : 2001Natur.413..809L . дои : 10.1038/35101535 . ПМИД 11677598 . S2CID 4364245 .
- ^ Чой С. (февраль 2002 г.). «Структуры калиевых каналов». Обзоры природы. Нейронаука . 3 (2): 115–21. дои : 10.1038/nrn727 . ПМИД 11836519 . S2CID 825973 .
- ^ Хилле Б., Армстронг К.М., Маккиннон Р. (октябрь 1999 г.). «Ионные каналы: от идеи к реальности». Природная медицина . 5 (10): 1105–9. дои : 10.1038/13415 . ПМИД 10502800 . S2CID 5216271 .
- ^ Хилле Б. (июнь 1973 г.). «Калиевые каналы в миелинизированном нерве. Избирательная проницаемость для малых катионов» . Журнал общей физиологии . 61 (6): 669–86. дои : 10.1085/jgp.61.6.669 . ПМК 2203488 . ПМИД 4541077 .
- ^ Jump up to: а б Носков С.Ю., Ру Б. (декабрь 2006 г.). «Ионная селективность в калиевых каналах». Биофизическая химия . 124 (3): 279–91. дои : 10.1016/j.bpc.2006.05.033 . ПМИД 16843584 .
- ^ Томпсон А.Н., Поссон DJ, Парса П.В., Nimigean CM (май 2008 г.). «Молекулярный механизм чувствительности к pH в калиевых каналах KcsA» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (19): 6900–5. Бибкод : 2008PNAS..105.6900T . дои : 10.1073/pnas.0800873105 . ПМК 2383984 . ПМИД 18443286 .
- ^ Jump up to: а б с Кордеро-Моралес Х.Ф., Куэлло Л.Г., Чжао Й., Джогини В., Кортес Д.М., Ру Б., Перозо Э. (апрель 2006 г.). «Молекулярные детерминанты стробирования селективного фильтра калиевых каналов». Структурная и молекулярная биология природы . 13 (4): 311–8. дои : 10.1038/nsmb1069 . ПМИД 16532009 . S2CID 20765018 .
- ^ Хирано М., Ониши Ю., Янагида Т., Иде Т. (ноябрь 2011 г.). «Роль цитоплазматического домена канала KcsA в рН-зависимом стробировании» . Биофизический журнал . 101 (9): 2157–62. Бибкод : 2011BpJ...101.2157H . дои : 10.1016/j.bpj.2011.09.024 . ПМК 3207171 . ПМИД 22067153 .
- ^ Ючи З., вице-президент Пау, Ян Д.С. (декабрь 2008 г.). «GCN4 повышает стабильность порового домена калиевого канала KcsA» . Журнал ФЭБС . 275 (24): 6228–36. дои : 10.1111/j.1742-4658.2008.06747.x . ПМИД 19016844 .
- ^ Эйзенман Г. (март 1962 г.). «Катионселективные стеклянные электроды и принцип их работы» . Биофизический журнал . 2 (2, часть 2): 259–323. Бибкод : 1962BpJ.....2..259E . дои : 10.1016/S0006-3495(62)86959-8 . ПМЦ 1366487 . ПМИД 13889686 .
- ^ Носков С.Ю., Бернеш С., Ру Б. (октябрь 2004 г.). «Контроль селективности ионов в калиевых каналах с помощью электростатических и динамических свойств карбонильных лигандов». Природа . 431 (7010): 830–4. Бибкод : 2004Natur.431..830N . дои : 10.1038/nature02943 . ПМИД 15483608 . S2CID 4414885 .
- ^ Безанилья Ф., Армстронг CM (ноябрь 1972 г.). «Отрицательная проводимость, вызванная попаданием ионов натрия и цезия в калиевые каналы аксонов кальмара» . Журнал общей физиологии . 60 (5): 588–608. дои : 10.1085/jgp.60.5.588 . ПМК 2226091 . ПМИД 4644327 .
- ^ Варма С., Ремпе С.Б. (август 2007 г.). «Настройка архитектуры координации ионов для обеспечения избирательного разделения» . Биофизический журнал . 93 (4): 1093–9. arXiv : физика/0608180 . Бибкод : 2007BpJ....93.1093V . дои : 10.1529/biophysj.107.107482 . ЧВК 1929028 . ПМИД 17513348 .
- ^ Томас М., Джаятилака Д., Корри Б. (октябрь 2007 г.). «Преобладающая роль координационного числа в селективности калиевых каналов» . Биофизический журнал . 93 (8): 2635–43. Бибкод : 2007BpJ....93.2635T . дои : 10.1529/biophysj.107.108167 . ЧВК 1989715 . ПМИД 17573427 .
- ^ Бостик Д.Л., Брукс К.Л. (май 2007 г.). «Селективность по К + каналов происходит из-за топологического контроля координированного состояния проникающих ионов» . Proceedings of the National Academy of Sciences of the United States of America . 104 (22): 9260–5. Бибкод : 2007PNAS..104.9260B . doi : 10.1073/pnas .0700554104 . ПМК 1890482 .
- ^ Деребе М.Г., Зауэр Д.Б., Цзэн В., Алам А., Ши Н., Цзян Й. (январь 2011 г.). «Настройка ионной селективности каналов тетрамерных катионов путем изменения количества мест связывания ионов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (2): 598–602. Бибкод : 2011PNAS..108..598D . дои : 10.1073/pnas.1013636108 . ПМК 3021048 . ПМИД 21187421 .
- ^ Сангинетти MC, Митчесон Дж.С. (март 2005 г.). «Прогнозирование взаимодействия препарата с каналом hERG, вызывающего приобретенный синдром удлиненного интервала QT». Тенденции в фармакологических науках . 26 (3): 119–24. дои : 10.1016/j.tips.2005.01.003 . ПМИД 15749156 .
- ^ Раджамани Р., Тунге Б.А., Ли Дж., Рейнольдс Ч. (март 2005 г.). «Модель гомологии hERG K с двумя состояниями + канал: применение для связывания лиганда». Bioorganic & Medicinal Chemistry Letters . 15 (6): 1737–41. doi : 10.1016/j.bmcl.2005.01.008 . PMID 15745831 .