Эукариот
| Эукариоты Временной диапазон: статериан – настоящее время, | |
|---|---|
| Научная классификация | |
| Домен: | Эукариоты ( Чаттон , 1925) Уиттакер и Маргулис , 1978 |
| Супергруппы и королевства [2] | |
| Синонимы | |




Эукариоты ə ( / j uː ˈ k æri oʊ t s , t - s əts / ) yoo- KARR -ee-ohts , - [5] составляют область эукариот которых или эукариот , организмов , клетки имеют мембраносвязанное ядро . Все животные , растения , грибы и многие одноклеточные организмы являются эукариотами. Они составляют основную группу форм жизни наряду с двумя группами прокариот : бактериями и археями . Эукариоты составляют небольшое меньшинство среди всех организмов, но, учитывая их обычно гораздо больший размер, их коллективная глобальная биомасса намного больше, чем у прокариот.
Эукариоты, по-видимому, возникли в составе архей Асгарда и тесно связаны с Heimdallarchaeia . [6] Это означает, что существует только две области жизни : бактерии и археи, а эукариоты включены в состав архей. Эукариоты впервые появились в палеопротерозое , вероятно, в виде жгутиковых клеток. Ведущая эволюционная теория утверждает, что они были созданы в результате симбиогенеза между анаэробными археями Асгарда и аэробными протеобактериями , которые образовали митохондрии . Второй эпизод симбиогенеза с цианобактерией создал растения с хлоропластами .
Эукариотические клетки содержат мембраносвязанные органеллы, такие как ядро , эндоплазматическая сеть и аппарат Гольджи . Эукариоты могут быть как одноклеточными , так и многоклеточными . Для сравнения, прокариоты обычно одноклеточные. Одноклеточных эукариот иногда называют протистами . Эукариоты могут размножаться как бесполым путем посредством митоза , так и половым путем посредством мейоза и гамет слияния ( оплодотворения ).
Разнообразие [ править ]
Эукариоты — это организмы , которые варьируются от микроскопических одиночных клеток , таких как пикозойи размером менее 3 микрометров в поперечнике, [7] таким животным, как синий кит , весом до 190 тонн и длиной до 33,6 метров (110 футов), [8] или такие растения , как прибрежное красное дерево , высотой до 120 метров (390 футов). [9] Многие эукариоты одноклеточные; неофициальная группа, называемая протистами, включает в себя многие из них, а также некоторые многоклеточные формы, такие как гигантские водоросли, длиной до 200 футов (61 м). [10] К многоклеточным эукариотам относятся животные, растения и грибы , но опять же, эти группы также содержат много одноклеточных видов . [11] Эукариотические клетки обычно намного крупнее, чем клетки прокариотов ( бактерий и архей) , их объем примерно в 10 000 раз больше. [12] [13] Эукариоты представляют собой небольшое меньшинство среди всех организмов , но, поскольку многие из них намного крупнее, их коллективная глобальная биомасса (468 гигатонн) намного больше, чем у прокариотов (77 гигатонн), причем одни только растения составляют более 81% общая биомасса Земли . [14]
- Размеры эукариот варьируются от одиночных клеток до организмов весом во многие тонны.






Эукариоты представляют собой разнообразную линию, состоящую в основном из микроскопических организмов . [15] Многоклеточность в той или иной форме развивалась независимо у эукариот по крайней мере 25 раз. [16] [17] Сложные многоклеточные организмы, не считая агрегации амеб с образованием слизевиков , развились лишь в пределах шести эукариотических линий: животных , симбиомикотовых грибов , бурых водорослей , красных водорослей , зеленых водорослей и наземных растений . [18] Эукариоты группируются по сходству генома, поэтому у групп часто отсутствуют видимые общие характеристики. [15]
Отличительные особенности [ править ]
Ядро [ править ]
Отличительной особенностью эукариот является то, что их клетки имеют ядра . Это дало им название от греческого εὖ ( eu , «хорошо» или «хорошо») и κάρυον ( karyon , «орех» или «ядро», что здесь означает «ядро»). [19] Эукариотические клетки имеют множество внутренних мембраносвязанных структур, называемых органеллами , и цитоскелет , который определяет организацию и форму клетки. клетки Ядро хранит ДНК , которая разделена на линейные пучки, называемые хромосомами ; [20] они разделяются на два совпадающих набора с помощью микротрубчатого веретена во время деления ядра в характерном для эукариотов процессе митоза . [21]
Биохимия [ править ]
Эукариоты отличаются от прокариотов во многих отношениях, с уникальными биохимическими путями, такими как синтез стерана . [22] Сигнатурные белки эукариот не имеют гомологии с белками из других областей жизни, но, по-видимому, универсальны среди эукариот. Они включают белки цитоскелета, сложную транскрипционную машину, системы мембранной сортировки, ядерную пору и некоторые ферменты биохимических путей. [23]
Внутренние мембраны [ править ]

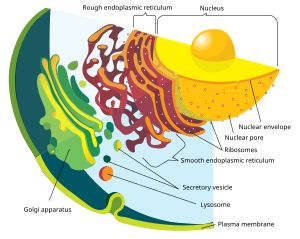
Клетки эукариот включают в себя множество мембраносвязанных структур, вместе образующих эндомембранную систему . [24] Простые отсеки, называемые везикулами и вакуолями , могут образовываться путем отпочкования от других мембран. Многие клетки поглощают пищу и другие материалы посредством процесса эндоцитоза , при котором внешняя мембрана инвагинируется , а затем отщипывается, образуя везикулу. [25] Некоторые клеточные продукты могут покидать везикулу посредством экзоцитоза . [26]
Ядро окружено двойной мембраной, известной как ядерная оболочка , с ядерными порами , которые позволяют веществу проникать внутрь и наружу. [27] Различные трубчатые и листовидные отростки ядерной мембраны образуют эндоплазматический ретикулум , который участвует в транспорте и созревании белков . Он включает шероховатый эндоплазматический ретикулум, покрытый рибосомами , синтезирующими белки; они попадают во внутреннее пространство или просвет. Впоследствии они обычно попадают в везикулы, которые отпочковываются от гладкой эндоплазматической сети. [28] У большинства эукариот эти несущие белок везикулы высвобождаются и далее модифицируются в стопки уплощенных везикул ( цистерны ), аппарат Гольджи . [29]
Везикулы могут быть специализированными; например, лизосомы содержат пищеварительные ферменты , которые расщепляют биомолекулы в цитоплазме. [30]
Митохондрии [ править ]

Митохондрии – это органеллы эукариотических клеток. Митохондрии обычно называют «электростанцией клетки». [31] за свою функцию по обеспечению энергией путем окисления сахаров или жиров с образованием запасающей энергию молекулы АТФ . [32] [33] Митохондрии имеют две окружающие мембраны , каждая из которых представляет собой бислой фосфолипидов ; внутренняя часть которых сложена в впячивания, называемые кристами , где аэробное дыхание . происходит [34]
Митохондрии содержат собственную ДНК , которая имеет близкое структурное сходство с бактериальной ДНК , из которой она произошла, и которая кодирует гены рРНК и тРНК, продуцирующие РНК, которая по структуре ближе к бактериальной РНК, чем к РНК эукариот. [35]
Некоторые эукариоты, такие как метамонады Giardia и Trichomonas , а также амебозои Pelomyxa , по-видимому, лишены митохондрий, но все они содержат органеллы, полученные из митохондрий, такие как гидрогеносомы или митосомы , вторично потерявшие свои митохондрии. [36] Они получают энергию путем ферментативного действия в цитоплазме. [37] [36]
Пластиды [ править ]
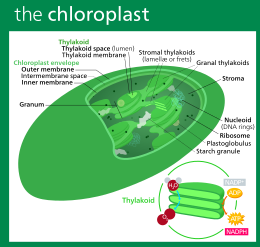
Растения и различные группы водорослей имеют пластиды не только митохондрии, но и . Пластиды, как и митохондрии, имеют собственную ДНК и развиваются из эндосимбионтов , в данном случае цианобактерий . Обычно они имеют форму хлоропластов , которые, как и цианобактерии, содержат хлорофилл и производят органические соединения (например, глюкозу ) посредством фотосинтеза . Другие занимаются хранением продуктов питания. Хотя пластиды, вероятно, имели единое происхождение, не все группы, содержащие пластиды, тесно связаны между собой. Вместо этого некоторые эукариоты получили их от других посредством вторичного эндосимбиоза или проглатывания. [38] Захват и секвестрация фотосинтезирующих клеток и хлоропластов, клептопластика , происходит у многих типов современных эукариотических организмов. [39] [40]
Цитоскелетные структуры [ править ]

Цитоскелет обеспечивает структуру жесткости и точки прикрепление к двигательным структурам, которые позволяют клетке двигаться, изменять форму или транспортировать материалы. Двигательные структуры представляют собой микрофиламенты актина , и актин-связывающие белки , в том числе α- актинин , фимбрин и филамин присутствующие в подмембранозных корковых слоях и пучках. Моторные белки микротрубочек динеин и кинезин , а также миозин актиновых филаментов обеспечивают динамический характер сети. [41] [42]
Многие эукариоты имеют длинные тонкие подвижные цитоплазматические выступы, называемые жгутиками , или несколько более коротких структур, называемых ресничками . Эти органеллы по-разному участвуют в движении, питании и ощущениях. Они состоят в основном из тубулина и полностью отличаются от жгутиков прокариот. Они поддерживаются пучком микротрубочек, возникающих из центриоли , обычно расположенных в виде девяти дублетов, окружающих два синглета. Жгутики могут иметь волоски ( мастигонемы ), как у многих Stramenopiles . клетки Их внутренняя часть непрерывна с цитоплазмой . [43] [44]
Центриоли часто присутствуют даже в клетках и группах, не имеющих жгутиков, а у хвойных и цветковых растений их нет. Обычно они встречаются группами, дающими начало различным микротрубчатым корням. Они образуют основной компонент цитоскелета и часто собираются в ходе нескольких клеточных делений, при этом один жгутик сохраняется от родителя, а другой происходит от него. Центриоли производят веретено во время ядерного деления. [45]
Клеточная стенка [ править ]
Клетки растений, водорослей, грибов и большинства хромальвеолатов , но не животных, окружены клеточной стенкой. Это слой вне клеточной мембраны , обеспечивающий клетке структурную поддержку, защиту и механизм фильтрации. Клеточная стенка также предотвращает чрезмерное расширение при попадании воды в клетку. [46]
Основными полисахаридами, составляющими первичную клеточную стенку наземных растений, являются целлюлоза , гемицеллюлоза и пектин . целлюлозы Микрофибриллы связаны между собой гемицеллюлозой, заключенной в пектиновую матрицу. Наиболее распространенной гемицеллюлозой первичной клеточной стенки является ксилоглюкан . [47]
Половое размножение [ править ]

Эукариоты имеют жизненный цикл, включающий половое размножение , чередующееся между гаплоидной фазой, когда в каждой клетке присутствует только одна копия каждой хромосомы, и диплоидной фазой, когда в каждой клетке имеется две копии каждой хромосомы. Диплоидная фаза образуется путем слияния двух гаплоидных гамет, таких как яйцеклетка и сперматозоиды , с образованием зиготы ; из него может вырасти тело, клетки которого делятся митозом и на каком-то этапе производят гаплоидные гаметы посредством мейоза — деления, которое уменьшает количество хромосом и создает генетическую изменчивость . [48] Эта модель имеет значительные вариации. Растения имеют как гаплоидную, так и диплоидную многоклеточную фазу . [49] Эукариоты имеют более низкую скорость метаболизма и более длительное время генерации, чем прокариоты, поскольку они крупнее и, следовательно, имеют меньшее соотношение площади поверхности к объему. [50]
Эволюция полового размножения может быть изначальной характеристикой эукариот. На основании филогенетического анализа Дакс и Роджер предположили, что у общего предка группы присутствовал факультативный пол. [51] Основной набор генов, которые функционируют в мейозе, присутствует как у Trichomonas vaginalis, так и у Giardia интестиналис , двух организмов, которые ранее считались бесполыми. [52] [53] Поскольку эти два вида являются потомками линий, которые рано отошли от эволюционного древа эукариот, основные мейотические гены и, следовательно, пол, вероятно, присутствовали у общего предка эукариот. [52] [53] Виды, которые когда-то считались бесполыми, такие как паразиты Leishmania , имеют половой цикл. [54] Амебы, ранее считавшиеся бесполыми, в древности были половыми; современные асексуальные группы, вероятно, возникли недавно. [55]
Эволюция [ править ]
История классификации [ править ]
В древности две линии животных и растений были признаны Аристотелем и Теофрастом . присвоил таксономический ранг Королевства этим линиям Линней в 18 веке . Хотя он с некоторыми оговорками включил грибы в состав растений, позже выяснилось, что они совершенно различны и заслуживают отдельного царства. [57] Различные одноклеточные эукариоты первоначально были помещены в растения или животные, когда они стали известны. В 1818 году немецкий биолог Георг А. Гольдфус ввёл слово «простейшие» для обозначения таких организмов , как инфузории , [58] и эта группа расширялась до тех пор, пока Эрнст Геккель не превратил ее в царство, охватывающее всех одноклеточных эукариотов, Protista , в 1866 году. [59] [60] [61] Таким образом, эукариоты стали рассматриваться как четыре царства:
В то время протистов считали «примитивными формами» и, таким образом, эволюционной ступенью , объединенной их примитивной одноклеточной природой. [60] Понимание древнейших разветвлений древа жизни существенно развилось только благодаря секвенированию ДНК , что привело к созданию системы доменов, а не царств, как ранга высшего уровня, выдвинутого Карлом Вёзе , Отто Кандлером и Марком Уиллисом в 1990 году, объединяющей всех эукариот. королевства в области «Эукариа», однако заявляя, что « эукариоты будут продолжать оставаться приемлемым общим синонимом». [3] [62] В 1996 году биолог-эволюционист Линн Маргулис предложила заменить Королевства и Домены «инклюзивными» названиями, чтобы создать «филогению, основанную на симбиозе», дав описание «Эукария (ядерные организмы, полученные из симбиоза)». [4]
Филогения [ править ]
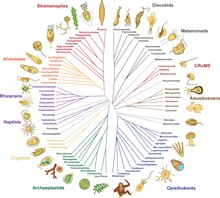
К 2014 году в результате филогеномных исследований, проведенных за предыдущие два десятилетия, начал формироваться грубый консенсус. [11] [63] Большинство эукариот можно отнести к одной из двух крупных клад, названных Amorphea (по составу схожей с гипотезой униконта ) и Diphoda (ранее биконты), которая включает растения и большинство линий водорослей. Третья основная группа, Excavata , была оставлена как формальная группа, поскольку она является парафилетической . [2] Предлагаемая ниже филогения включает только одну группу раскопок ( Discoba ), [64] и включает предположение 2021 года о том, что пикозои являются близкими родственниками родофитов. [65] Провора . — группа микробных хищников, обнаруженная в 2022 году [1]
| Эукариоты |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2200 млн лет назад |
Один взгляд на великие королевства и их основные группы. [64] [66] [67] [15] Метамонаду Дискобы трудно определить, поскольку она является сестрой, возможно, , возможно, Малавимонады . [15]
Происхождение эукариот [ править ]

Возникновение эукариотической клетки, или эукариогенез , является важной вехой в эволюции жизни, поскольку к эукариотам относятся все сложные клетки и почти все многоклеточные организмы. Последний общий предок эукариот (LECA) является гипотетическим источником всех ныне живущих эукариот. [69] и, скорее всего, представлял собой биологическую популяцию , а не отдельную особь. [70] Считается, что LECA представлял собой простейшего с ядром, по крайней мере, одной центриолью и жгутиком , факультативно аэробными митохондриями, полом ( мейоз и сингамия ), спящей кистой с клеточной стенкой из хитина или целлюлозы и пероксисомами . [71] [72] [73]
Эндосимбиотический союз между подвижными анаэробными архей и аэробными альфапротеобактериями дал начало LECA и всем эукариотам с митохондриями . Второй, гораздо более поздний эндосимбиоз с цианобактерией дал начало предку растений с хлоропластами . [68]
Наличие эукариотических биомаркеров у архей указывает на архейное происхождение. Геномы архей Асгарда содержат множество генов сигнатурных белков эукариот , которые играют решающую роль в развитии цитоскелета и сложных клеточных структур, характерных для эукариот. В 2022 году криоэлектронная томография продемонстрировала, что археи Асгарда имеют сложный цитоскелет на основе актина , что стало первым прямым визуальным свидетельством архейного происхождения эукариот. [74]
Окаменелости [ править ]
Время возникновения эукариот определить трудно, но открытие Qingshania magnificia , самого раннего многоклеточного эукариота из Северного Китая, жившего 1,635 миллиарда лет назад, позволяет предположить, что эукариоты коронной группы произошли из позднего палеопротерозоя ( статерианского периода ); В Северном Китае также обнаружены самые ранние одноклеточные эукариоты, жившие примерно 1,65 миллиарда лет назад: Tappania plana , Shuiyousphaeridium macroreticulatum , Dictyosphaera macroreticulata , Germinosphaera alveolata и Valeria lophostriata . [75]
Некоторые акритархи известны как минимум 1,65 миллиарда лет назад, а возраст ископаемого Grypania , который может быть водорослью, составляет целых 2,1 миллиарда лет. [76] [77] « Проблематичный » [78] ископаемая Дискагма была обнаружена в палеопочвах возрастом 2,2 миллиарда лет. [78]

Структуры, которые, как предполагается, представляют собой «крупные колониальные организмы», были обнаружены в черных сланцах палеопротерозоя , таких как формация Франсвилиан B в Габоне , получившая название « биота Франсвилья », возраст которой составляет 2,1 миллиарда лет. [79] [80] Однако статус этих структур как окаменелостей оспаривается, и другие авторы предполагают, что они могут представлять собой псевдоокаменелости . [81] Самые старые окаменелости, которые можно однозначно отнести к эукариотам, относятся к группе Руян в Китае и датируются примерно 1,8–1,6 миллиарда лет назад. [82] Окаменелости, которые явно связаны с современными группами, начали появляться примерно 1,2 миллиарда лет назад в виде красных водорослей , хотя недавние исследования предполагают существование окаменелых нитчатых водорослей в бассейне Виндхья , датируемых, возможно, 1,6–1,7 миллиарда лет назад. [83]
Присутствие стеранов , специфичных для эукариот биомаркеров , в австралийских сланцах ранее указывало на то, что эукариоты присутствовали в этих породах возрастом 2,7 миллиарда лет. [22] [84] но эти архейские биомаркеры были опровергнуты как более поздние загрязнители. [85] Возраст самых старых действительных записей биомаркеров составляет всего около 800 миллионов лет. [86] Напротив, анализ молекулярных часов предполагает появление биосинтеза стеринов уже 2,3 миллиарда лет назад. [87] Природа стеранов как биомаркеров эукариот еще больше усложняется продукцией стеринов некоторыми бактериями. [88] [89]
Когда бы ни возникли эукариоты, они, возможно, стали экологически доминирующими гораздо позже; Массовое увеличение состава цинка в морских отложениях 800 миллионов лет назад было связано с появлением значительных популяций эукариот, которые преимущественно потребляют и включают цинк по сравнению с прокариотами, примерно через миллиард лет после их происхождения (самое позднее). [90]
См. также [ править ]
- Гибридный геном эукариот
- Список секвенированных геномов эукариот
- Паракарион миоджиненсис
- Хранилище (органелла)
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б Тихоненков Д.В., Михайлов КВ, Гаврилюк Р.М. и др. (декабрь 2022 г.). «Микробные хищники образуют новую супергруппу эукариот». Природа . 612 (7941): 714–719. Бибкод : 2022Natur.612..714T . дои : 10.1038/s41586-022-05511-5 . ПМИД 36477531 . S2CID 254436650 .
- ↑ Перейти обратно: Перейти обратно: а б Адл С.М., Басс Д., Лейн CE и др. (январь 2019 г.). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ↑ Перейти обратно: Перейти обратно: а б Вёзе Ч.Р. , Кандлер О. , Уилис М.Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ↑ Перейти обратно: Перейти обратно: а б Маргулис Л. (6 февраля 1996 г.). «Архейно-эубактериальные слияния в происхождении эукариев: филогенетическая классификация жизни» . Труды Национальной академии наук . 93 (3): 1071–1076. Бибкод : 1996PNAS...93.1071M . дои : 10.1073/pnas.93.3.1071 . ПМЦ 40032 . ПМИД 8577716 .
- ^ «эукариот» . Словарь Merriam-Webster.com . Проверено 12 мая 2024 г.
- ^ Эме, Лаура; Тамарит, Дэниел; Касерес, Ева Ф.; Лестница, Кортни В.; Де Анда, Валери; Шён, Макс Э.; Зейтц, Кили В.; Домбровский, Нина; Льюис, Уильям Х.; Хома, Феликс; Пила, Джимми Х.; Ломбард, Джонатан; Нунура, Такуро; Ли, Вэнь-Цзюнь; Хуа, Чжэн-Шуан; Чен, Линь-Син; Банфилд, Джиллиан Ф.; Джон, Эмили Ст; Рейзенбах, Анна-Луиза; Стотт, Мэтью Б.; Шрамм, Андреас; Кьельдсен, Каспер У.; Теске, Андреас П.; Бейкер, Бретт Дж.; Эттема, Тейс Дж.Г. (29 июня 2023 г.). «Вывод и реконструкция хеймдаллархейного происхождения эукариот» . Природа . 618 (7967): 992–999. Бибкод : 2023Natur.618..992E . дои : 10.1038/s41586-023-06186-2 . ISSN 1476-4687 . ПМЦ 10307638 . ПМИД 37316666 .
- ^ Сенивасан Р., Саусен Н., Медлин Л.К., Мелконян М. (26 марта 2013 г.). «Picomonas judraskeda Gen. Et Sp. Nov.: первый идентифицированный член семейства Picozoa Nov., широко распространенной группы пикоукариот, ранее известной как «пикобилифиты » . ПЛОС ОДИН . 8 (3): e59565. Бибкод : 2013PLoSO...859565S . дои : 10.1371/journal.pone.0059565 . ПМК 3608682 . ПМИД 23555709 .
- ^ Вуд Дж. (1983). Книга рекордов Гиннеса по фактам и подвигам животных . Энфилд, Миддлсекс: превосходная степень Книги Гиннеса. ISBN 978-0-85112-235-9 .
- ^ Эрл CJ, изд. (2017). «Секвойя вечнозеленая» . База данных голосеменных растений . Архивировано из оригинала 1 апреля 2016 года . Проверено 15 сентября 2017 г.
- ^ ван ден Хук С., Манн Д., Янс Х. (1995). Водоросли. Введение в психологию . Кембридж: Издательство Кембриджского университета. ISBN 0-521-30419-9 . Архивировано из оригинала 10 февраля 2023 года . Проверено 7 апреля 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Бурки Ф (май 2014 г.). «Эукариотическое древо жизни с глобальной филогеномной точки зрения» . Перспективы Колд-Спринг-Харбор в биологии . 6 (5): а016147. doi : 10.1101/cshperspect.a016147 . ПМЦ 3996474 . ПМИД 24789819 .
- ^ ДеРенно Б (2001). «Эукариоты, происхождение». Энциклопедия биоразнообразия . Том. 2. Эльзевир. стр. 329–332. дои : 10.1016/b978-0-12-384719-5.00174-x . ISBN 9780123847201 .
- ^ Ямагучи М., Уорман, Колорадо (2014). «Глубоководные микроорганизмы и происхождение эукариотической клетки» (PDF) . Японский журнал протозоологии . 47 (1, 2): 29–48. Архивировано из оригинала (PDF) 9 августа 2017 года.
- ^ Бар-Он, Инон М.; Филлипс, Роб; Майло, Рон (17 мая 2018 г.). «Распределение биомассы на Земле» . Труды Национальной академии наук . 115 (25): 6506–6511. Бибкод : 2018PNAS..115.6506B . дои : 10.1073/pnas.1711842115 . ISSN 0027-8424 . ПМК 6016768 . ПМИД 29784790 .
- ↑ Перейти обратно: Перейти обратно: а б с д Бурки Ф., Роджер Эй.Дж., Браун М.В., Симпсон АГ (2020). «Новое дерево эукариотов» . Тенденции в экологии и эволюции . 35 (1). Эльзевир Б.В.: 43–55. дои : 10.1016/j.tree.2019.08.008 . ISSN 0169-5347 . ПМИД 31606140 . S2CID 204545629 .
- ^ Гросберг Р.К., Стратманн Р.Р. (2007). «Эволюция многоклеточности: незначительный крупный переход?» (PDF) . Annu Rev Ecol Evol Syst . 38 : 621–654. doi : 10.1146/annurev.ecolsys.36.102403.114735 . Архивировано (PDF) из оригинала 14 марта 2023 года . Проверено 8 апреля 2023 г.
- ^ Парфри Л., Лар Д. (2013). «Многоклеточность возникала несколько раз в эволюции эукариот» (PDF) . Биоэссе . 35 (4): 339–347. дои : 10.1002/bies.201200143 . ПМИД 23315654 . S2CID 13872783 . Архивировано (PDF) из оригинала 25 июля 2014 года . Проверено 8 апреля 2023 г.
- ^ Поппер З.А., Мишель Г., Эрве С., Домозич Д.С., Уиллатс В.Г., Туохи М.Г., Клоарег Б., Стенгель Д.Б. (2011). «Эволюция и разнообразие стенок растительных клеток: от водорослей до цветковых растений». Ежегодный обзор биологии растений . 62 : 567–590. doi : 10.1146/annurev-arplant-042110-103809 . hdl : 10379/6762 . ПМИД 21351878 . S2CID 11961888 .
- ^ Харпер, Дуглас. «эукариотический» . Интернет-словарь этимологии .
- ^ Бонев Б., Кавалли Г (14 октября 2016 г.). «Организация и функции 3D-генома». Обзоры природы Генетика . 17 (11): 661–678. дои : 10.1038/nrg.2016.112 . hdl : 2027.42/151884 . ПМИД 27739532 . S2CID 31259189 .
- ^ О'Коннор, Клэр (2008). «Сегрегация хромосом: роль центромер» . Природное образование . Проверено 18 февраля 2024 г.
эукар
- ↑ Перейти обратно: Перейти обратно: а б Брокс Джей-Джей, Логан Г.А., Бьюик Р., Саммонс Р.Э. (август 1999 г.). «Архейские молекулярные окаменелости и раннее появление эукариотов». Наука . 285 (5430): 1033–1036. Бибкод : 1999Sci...285.1033B . CiteSeerX 10.1.1.516.9123 . дои : 10.1126/science.285.5430.1033 . ПМИД 10446042 .
- ^ Хартман Х, Федоров А (февраль 2002 г.). «Происхождение эукариотической клетки: геномное исследование» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (3): 1420–5. Бибкод : 2002PNAS...99.1420H . дои : 10.1073/pnas.032658599 . ПМК 122206 . ПМИД 11805300 .
- ^ Линка М., Вебер А.П. (2011). «Эволюционная интеграция метаболизма хлоропластов с метаболическими сетями клеток» . В Burnap RL, Vermaas WF (ред.). Функциональная геномика и эволюция фотосинтетических систем . Спрингер. п. 215. ИСБН 978-94-007-1533-2 . Архивировано из оригинала 29 мая 2016 года . Проверено 27 октября 2015 г.
- ^ Марш М. (2001). Эндоцитоз . Издательство Оксфордского университета. п. VII. ISBN 978-0-19-963851-2 .
- ^ Сталдер Д., Гершлик, округ Колумбия (ноябрь 2020 г.). «Прямые пути переноса от аппарата Гольджи к плазматической мембране» . Семинары по клеточной биологии и биологии развития . 107 : 112–125. дои : 10.1016/j.semcdb.2020.04.001 . ПМК 7152905 . ПМИД 32317144 .
- ^ Hetzer MW (март 2010 г.). «Ядерная оболочка» . Перспективы Колд-Спринг-Харбор в биологии . 2 (3): а000539. doi : 10.1101/cshperspect.a000539 . ПМК 2829960 . ПМИД 20300205 .
- ^ «Эндоплазматическая сеть (шероховатая и гладкая)» . Британское общество клеточной биологии. Архивировано из оригинала 24 марта 2019 года . Проверено 12 ноября 2017 г.
- ^ «Аппарат Гольджи» . Британское общество клеточной биологии. Архивировано из оригинала 13 ноября 2017 года . Проверено 12 ноября 2017 г.
- ^ «Лизосома» . Британское общество клеточной биологии. Архивировано из оригинала 13 ноября 2017 года . Проверено 12 ноября 2017 г.
- ^ Сайгин Д., Табиб Т., Биттар Х.Э. и др. (июль 1957 г.). «Транскрипционное профилирование популяций клеток легких при идиопатической легочной артериальной гипертензии» . Легочное кровообращение . 10 (1): 131–144. Бибкод : 1957SciAm.197a.131S . doi : 10.1038/scientificamerican0757-131 . ПМК 7052475 . ПМИД 32166015 .
- ^ Voet D, Voet JC, Пратт CW (2006). Основы биохимии (2-е изд.). Джон Уайли и сыновья. стр. 547, 556 . ISBN 978-0471214953 .
- ^ Мак С. (1 мая 2006 г.). «Re: Существуют ли эукариотические клетки без митохондрий?» . madsci.org . Архивировано из оригинала 24 апреля 2014 года . Проверено 24 апреля 2014 г.
- ^ Зик М., Рабл Р., Райхерт А.С. (январь 2009 г.). «Ультраструктура и функция митохондрий, связывающая образование крист». Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1793 (1): 5–19. дои : 10.1016/j.bbamcr.2008.06.013 . ПМИД 18620004 .
- ^ Уотсон Дж., Хопкинс Н., Робертс Дж., Стейц Дж.А., Вайнер А. (1988). «28: Истоки жизни» . Молекулярная биология гена (Четвертое изд.). Менло-Парк, Калифорния: Издательство Benjamin/Cummings Publishing Company, Inc., с. 1154 . ISBN 978-0-8053-9614-0 .
- ↑ Перейти обратно: Перейти обратно: а б Карнковска А, Вацек В, Зубакова З и др. (май 2016 г.). «Эукариот без митохондриальной органеллы» . Современная биология . 26 (10): 1274–1284. Бибкод : 2016CBio...26.1274K . дои : 10.1016/j.cub.2016.03.053 . ПМИД 27185558 .
- ^ Дэвис Дж.Л. (13 мая 2016 г.). «Ученые потрясены, обнаружив эукариот без митохондрий» . ИФЛ Наука . Архивировано из оригинала 17 февраля 2019 года . Проверено 13 мая 2016 г.
- ^ Сато Н. (2006). «Происхождение и эволюция пластид: геномный взгляд на объединение и разнообразие пластид». В Wise RR, Хубер Дж.К. (ред.). Строение и функции пластид . Достижения в области фотосинтеза и дыхания. Том. 23. Спрингер Нидерланды. стр. 75–102. дои : 10.1007/978-1-4020-4061-0_4 . ISBN 978-1-4020-4060-3 .
- ^ Миннхаген С., Карвалью В.Ф., Саломон П.С., Янсон С. (сентябрь 2008 г.). «Содержание ДНК хлоропластов у Dinophys (Dinophyceae) на разных стадиях клеточного цикла соответствует клептопластике». Окружающая среда. Микробиол . 10 (9): 2411–7. Бибкод : 2008EnvMi..10.2411M . дои : 10.1111/j.1462-2920.2008.01666.x . ПМИД 18518896 .
- ^ Бодил А (февраль 2018 г.). «Развились ли некоторые пластиды, полученные из красных водорослей, посредством клептопластидии? Гипотеза». Биологические обзоры Кембриджского философского общества . 93 (1): 201–222. дои : 10.1111/brv.12340 . ПМИД 28544184 . S2CID 24613863 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (1 января 2002 г.). «Молекулярные моторы». Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . Архивировано из оригинала 8 марта 2019 года . Проверено 6 апреля 2023 г.
- ^ Суини Х.Л., Хольцбаур Э.Л. (май 2018 г.). «Моторные белки» . Перспективы Колд-Спринг-Харбор в биологии . 10 (5): а021931. doi : 10.1101/cshperspect.a021931 . ПМЦ 5932582 . ПМИД 29716949 .
- ^ Барди С.Л., Нг С.И., Джаррелл К.Ф. (февраль 2003 г.). «Структуры подвижности прокариот» . Микробиология . 149 (Часть 2): 295–304. дои : 10.1099/mic.0.25948-0 . ПМИД 12624192 .
- ^ Компакт-диск Silflow, Лефевр, Пенсильвания (декабрь 2001 г.). «Сборка и подвижность эукариотических ресничек и жгутиков. Уроки Chlamydomonas Reinhardtii» . Физиология растений . 127 (4): 1500–7. дои : 10.1104/стр.010807 . ПМК 1540183 . ПМИД 11743094 .
- ^ Воробьёв И.А., Надеждина Е.С. (1987). Центросома и ее роль в организации микротрубочек . Международный обзор цитологии. Том. 106. С. 227–293. дои : 10.1016/S0074-7696(08)61714-3 . ISBN 978-0-12-364506-7 . ПМИД 3294718 .
- ^ Хауленд Дж.Л. (2000). Удивительные археи: открытие еще одной области жизни . Оксфорд: Издательство Оксфордского университета. стр. 69–71. ISBN 978-0-19-511183-5 .
- ^ Фрай СК (1989). «Структура и функции ксилоглюкана». Журнал экспериментальной ботаники . 40 (1): 1–11. дои : 10.1093/jxb/40.1.1 .
- ^ Гамильтон МБ (2009). Популяционная генетика . Уайли-Блэквелл . п. 55 . ISBN 978-1-4051-3277-0 .
- ^ Тейлор Т.Н., Керп Х., Хасс Х. (2005). «Биология истории жизни ранних наземных растений: расшифровка фазы гаметофита» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (16): 5892–5897. дои : 10.1073/pnas.0501985102 . ПМК 556298 . ПМИД 15809414 .
- ^ Переулок N (июнь 2011 г.). «Энергетика и генетика между прокариотами и эукариотами» . Биология Директ . 6 (1): 35. дои : 10.1186/1745-6150-6-35 . ПМЦ 3152533 . ПМИД 21714941 .
- ^ Дакс Дж., Роджер А.Дж. (июнь 1999 г.). «Первая половая линия и актуальность факультативного секса». Журнал молекулярной эволюции . 48 (6): 779–783. Бибкод : 1999JMolE..48..779D . дои : 10.1007/PL00013156 . ПМИД 10229582 . S2CID 9441768 .
- ↑ Перейти обратно: Перейти обратно: а б Рамеш М.А., Малик С.Б., Логсдон Дж.М. (январь 2005 г.). «Филогеномный перечень мейотических генов; доказательства пола у лямблий и раннего эукариотического происхождения мейоза» . Современная биология . 15 (2): 185–191. Бибкод : 2005CBio...15..185R . дои : 10.1016/j.cub.2005.01.003 . ПМИД 15668177 . S2CID 17013247 .
- ↑ Перейти обратно: Перейти обратно: а б Малик С.Б., Пайтлинг А.В., Стефаниак Л.М., Шурко А.М., Логсдон Дж.М. (август 2007 г.). Хан М.В. (ред.). «Расширенный список консервативных мейотических генов доказывает наличие пола у Trichomonas vaginalis» . ПЛОС ОДИН . 3 (8): e2879. Бибкод : 2008PLoSO...3.2879M . дои : 10.1371/journal.pone.0002879 . ПМЦ 2488364 . ПМИД 18663385 .
- ^ Акопьянц Н.С., Кимблин Н., Секундино Н., Патрик Р., Питерс Н., Юрист П., Добсон Д.Е., Беверли С.М., Сакс Д.Л. (апрель 2009 г.). «Демонстрация генетического обмена во время циклического развития Leishmania у переносчика москитов» . Наука . 324 (5924): 265–268. Бибкод : 2009Sci...324..265A . дои : 10.1126/science.1169464 . ПМК 2729066 . ПМИД 19359589 .
- ^ Лар DJ, Парфри Л.В., Митчелл Э.А., Кац Л.А., Лара Э (июль 2011 г.). «Целомудрие амеб: переоценка доказательств пола у амебоидных организмов» . Труды: Биологические науки . 278 (1715): 2081–2090. дои : 10.1098/rspb.2011.0289 . ПМК 3107637 . ПМИД 21429931 .
- ^ Патрик Дж. Килинг ; Яна Эглит (21 ноября 2023 г.). «Открыто доступные иллюстрации как инструменты для описания эукариотического микробного разнообразия» . ПЛОС Биология . 21 (11): e3002395. doi : 10.1371/JOURNAL.PBIO.3002395 . ISSN 1544-9173 . ПМЦ 10662721 . ПМИД 37988341 . Викиданные Q123558544 .
- ^ Мур RT (1980). «Таксономические предложения по классификации морских дрожжей и других дрожжеподобных грибов, включая головни». Ботаника Марина . 23 : 361–373.
- ^ Гольдфус (1818 г.). «Ueber die Classification der Zoophyten» [К классификации зоофитов]. Исида, Одер, Encyclopädische Zeitung von Oken (на немецком языке). 2 (6): 1008–1019. Архивировано из оригинала 24 марта 2019 года . Проверено 15 марта 2019 г. Из стр. 1008: «Эрсте Класс. Уртьер. Простейшие». (Первый класс. Первобытные животные. Простейшие.) [Примечание: каждая графа на каждой странице этого журнала пронумерована; на странице есть две колонки.]
- ^ Скамарделла Дж. М. (1999). «Не растения и не животные: краткая история происхождения королевств Protozoa, Protista и Protoctista» (PDF) . Международная микробиология . 2 (4): 207–221. ПМИД 10943416 . Архивировано из оригинала (PDF) 14 июня 2011 года.
- ↑ Перейти обратно: Перейти обратно: а б Ротшильд ЖЖ (1989). «Простейшие, Протиста, Протоктиста: что в имени?» . Журнал истории биологии . 22 (2): 277–305. дои : 10.1007/BF00139515 . ПМИД 11542176 . S2CID 32462158 . Архивировано из оригинала 4 февраля 2020 года . Проверено 4 февраля 2020 г.
- ^ Уиттакер Р.Х. (январь 1969 г.). «Новые концепции царств или организмов. Эволюционные отношения лучше представлены новыми классификациями, чем традиционными двумя царствами». Наука . 163 (3863): 150–60. Бибкод : 1969Sci...163..150W . CiteSeerX 10.1.1.403.5430 . дои : 10.1126/science.163.3863.150 . ПМИД 5762760 .
- ^ Нолл АХ (1992). «Ранняя эволюция эукариотов: геологическая перспектива». Наука . 256 (5057): 622–627. Бибкод : 1992Sci...256..622K . дои : 10.1126/science.1585174 . ПМИД 1585174 .
Эукарии, или эукариоты
- ^ Бурки Ф., Каплан М., Тихоненков Д.В. и др. (январь 2016 г.). «Распутывание ранней диверсификации эукариот: филогеномное исследование эволюционного происхождения Centrohelida, Haptophyta и Cryptista» . Труды: Биологические науки . 283 (1823): 20152802. doi : 10.1098/rspb.2015.2802 . ПМК 4795036 . ПМИД 26817772 .
- ↑ Перейти обратно: Перейти обратно: а б Браун М.В., Хейсс А.А., Камикава Р., Инагаки Ю., Ябуки А., Тайс А.К., Ширатори Т., Исида К.И., Хасимото Т., Симпсон А., Роджер А. (19 января 2018 г.). «Филогеномика помещает сиротские линии протистанов в новую супергруппу эукариот» . Геномная биология и эволюция . 10 (2): 427–433. дои : 10.1093/gbe/evy014 . ПМЦ 5793813 . ПМИД 29360967 .
- ^ Шён М.Е., Златогурский В.В., Сингх Р.П. и др. (2021). «Пикозоа — это архепластиды без пластид» . Природные коммуникации . 12 (1): 6651. bioRxiv 10.1101/2021.04.14.439778 . дои : 10.1038/s41467-021-26918-0 . ПМК 8599508 . ПМИД 34789758 . S2CID 233328713 . Архивировано из оригинала 2 февраля 2024 года . Проверено 20 декабря 2021 г.
- ^ Шён М.Е., Златогурский В.В., Сингх Р.П. и др. (2021). «Пикозоа — это архепластиды без пластид» . Природные коммуникации . 12 (1): 6651. bioRxiv 10.1101/2021.04.14.439778 . дои : 10.1038/s41467-021-26918-0 . ПМК 8599508 . ПМИД 34789758 . S2CID 233328713 .
- ^ Тихоненков Д.В., Михайлов КВ, Гаврилюк Р.М. и др. (декабрь 2022 г.). «Микробные хищники образуют новую супергруппу эукариот». Природа . 612 (7941): 714–719. дои : 10.1038/s41586-022-05511-5 . ПМИД 36477531 . S2CID 254436650 .
- ↑ Перейти обратно: Перейти обратно: а б Латорре А., Дурбан А., Моя А., Перето Дж. (2011). «Роль симбиоза в эволюции эукариот» . В Гарго М., Лопес-Гарсия П., Мартин Х. (ред.). Происхождение и эволюция жизни: астробиологическая перспектива . Кембридж: Издательство Кембриджского университета. стр. 326–339. ISBN 978-0-521-76131-4 . Архивировано из оригинала 24 марта 2019 года . Проверено 27 августа 2017 г.
- ^ Габальдон Т. (октябрь 2021 г.). «Происхождение и ранняя эволюция эукариотической клетки». Ежегодный обзор микробиологии . 75 (1): 631–647. doi : 10.1146/annurev-micro-090817-062213 . ПМИД 34343017 . S2CID 236916203 .
- ^ О'Мэлли М.А., Леже М.М., Уайдман Дж.Г., Руис-Трилло I (март 2019 г.). «Представления о последнем общем предке эукариот». Экология и эволюция природы . 3 (3): 338–344. Бибкод : 2019NatEE...3..338O . дои : 10.1038/s41559-019-0796-3 . hdl : 10261/201794 . ПМИД 30778187 . S2CID 67790751 .
- ^ Леандер Б.С. (май 2020 г.). «Хищные протисты» . Современная биология . 30 (10): 510–516 р. Бибкод : 2020CBio...30.R510L . дои : 10.1016/j.cub.2020.03.052 . ПМИД 32428491 . S2CID 218710816 .
- ^ Штрассерт Дж. Ф., Ирисарри I, Уильямс Т. А., Бурки Ф (март 2021 г.). «Молекулярная временная шкала эволюции эукариот с последствиями для происхождения пластид, полученных из красных водорослей» . Природные коммуникации . 12 (1): 1879. Бибкод : 2021NatCo..12.1879S . дои : 10.1038/s41467-021-22044-z . ПМЦ 7994803 . ПМИД 33767194 .
- ^ Куманду В.Л., Уикстед Б., Джинджер М.Л., ван дер Гизен М., Дакс Дж.Б., Филд MC (2013). «Молекулярная палеонтология и сложность последнего общего предка эукариот» . Критические обзоры по биохимии и молекулярной биологии . 48 (4): 373–396. дои : 10.3109/10409238.2013.821444 . ПМЦ 3791482 . ПМИД 23895660 .
- ^ Родригес-Оливейра Т., Волвебер Ф., Понсе-Толедо Р.И. и др. (2023). «Актиновый цитоскелет и сложная клеточная архитектура у архей Асгарда» . Природа . 613 (7943): 332–339. Бибкод : 2023Natur.613..332R . дои : 10.1038/s41586-022-05550-y . hdl : 20.500.11850/589210 . ПМЦ 9834061 . ПМИД 36544020 .
- ^ Мяо, Л.; Инь, З.; Нолл, АХ; Цюй, Ю.; Чжу, М. (2024). «Многоклеточные эукариоты возрастом 1,63 миллиарда лет из формации Чуанлингоу в Северном Китае» . Достижения науки . 10 (4): eadk3208. Бибкод : 2024SciA...10K3208M . дои : 10.1126/sciadv.adk3208 . ПМЦ 10807817 . ПМИД 38266082 .
- ^ Хан Т.М., Раннегар Б. (июль 1992 г.). «Мегаскопические эукариотические водоросли из железоформации Негауни возрастом 2,1 миллиарда лет, Мичиган». Наука . 257 (5067): 232–5. Бибкод : 1992Sci...257..232H . дои : 10.1126/science.1631544 . ПМИД 1631544 .
- ^ Нолл А.Х., Яво Э.Дж., Хьюитт Д., Коэн П. (июнь 2006 г.). «Эукариотические организмы в протерозойских океанах» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 361 (1470): 1023–1038. дои : 10.1098/rstb.2006.1843 . ПМЦ 1578724 . ПМИД 16754612 .
- ↑ Перейти обратно: Перейти обратно: а б с Реталлак Г.Дж., Крулл Э.С., Текрей Г.Д., Паркинсон Д.Х. (2013). «Проблемные окаменелости в форме урны из палеопротерозойской (2,2 млрд лет) палеопочвы в Южной Африке». Докембрийские исследования . 235 : 71–87. Бибкод : 2013PreR..235...71R . doi : 10.1016/j.precamres.2013.05.015 .
- ^ Эль Альбани А., Бенгтсон С., Кэнфилд Д.Э. и др. (июль 2010 г.). «Крупные колониальные организмы с скоординированным ростом в насыщенной кислородом среде 2,1 миллиарда лет назад». Природа . 466 (7302): 100–104. Бибкод : 2010Natur.466..100A . дои : 10.1038/nature09166 . ПМИД 20596019 . S2CID 4331375 .
- ^ Эль Альбани, Абдерразак (2023). «Поиск жизни в палеопротерозойских морских отложениях с использованием изотопов Zn и геохимии» (PDF) . Письма о Земле и планетологии . 623 : 118169. Бибкод : 2023E&PSL.61218169E . дои : 10.1016/j.epsl.2023.118169 . S2CID 258360867 .
- ^ Осса Осса, Франц; Понс, Мари-Лора; Беккер, Андрей; Хофманн, Аксель; Поултон, Саймон В.; и др. (2023). «Обогащение цинком и изотопное фракционирование в морской среде обитания Францевильской группы около 2,1 млрд лет назад: признак использования цинка эукариотами?» (PDF) . Письма о Земле и планетологии . 611 : 118147. Бибкод : 2023E&PSL.61118147O . дои : 10.1016/j.epsl.2023.118147 .
- ^ Фахри, Моджтаба; Тархан Лидия Георгиевна; Рейнхард, Кристофер Т.; Кроу, Шон А.; Лайонс, Тимоти В.; Планавски, Ной Дж. (май 2023 г.). «Окисление поверхности Земли и возникновение эукариотической жизни: новый взгляд на связь с экскурсией положительных изотопов углерода в Ломагунди» . Обзоры наук о Земле . 240 : 104398. Бибкод : 2023ESRv..24004398F . doi : 10.1016/j.earscirev.2023.104398 . S2CID 257761993 .
- ^ Бенгтсон С., Беливанова В., Расмуссен Б., Уайтхаус М. (май 2009 г.). «Спорные «кембрийские» окаменелости виндхийцев реальны, но старше их более чем на миллиард лет» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (19): 7729–7734. Бибкод : 2009PNAS..106.7729B . дои : 10.1073/pnas.0812460106 . ПМЦ 2683128 . ПМИД 19416859 .
- ^ Уорд П. (9 февраля 2008 г.). «Массовые вымирания: микробы наносят ответный удар» . Новый учёный . стр. 40–43. Архивировано из оригинала 8 июля 2008 года . Проверено 27 августа 2017 г.
- ^ Френч К.Л., Холлманн С., Хоуп Дж.М., Шун П.Л., Зумберге Дж.А., Хосино Ю., Питерс К.А., Джордж С.К., Лав Г.Д., Брокс Дж.Дж., Бьюик Р., Саммонс Р.Э. (май 2015 г.). «Переоценка углеводородных биомаркеров в архейских породах» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (19): 5915–5920. Бибкод : 2015PNAS..112.5915F . дои : 10.1073/pnas.1419563112 . ПМЦ 4434754 . ПМИД 25918387 .
- ^ Брокс Джей Джей, Джарретт Эй Джей, Сирантуан Э, Холлманн С, Хосино Ю, Лиянаге Т (август 2017 г.). «Рост водорослей в криогенных океанах и появление животных». Природа . 548 (7669): 578–581. Бибкод : 2017Natur.548..578B . дои : 10.1038/nature23457 . ПМИД 28813409 . S2CID 205258987 .
- ^ Голд DA, Кэрон А., Фурнье GP, Summons RE (март 2017 г.). «Палеопротерозойский биосинтез стеринов и появление кислорода» . Природа . 543 (7645): 420–423. Бибкод : 2017Natur.543..420G . дои : 10.1038/nature21412 . hdl : 1721.1/128450 . ПМИД 28264195 . S2CID 205254122 .
- ^ Вэй Дж. Х., Инь X, Веландер П. В. (24 июня 2016 г.). «Синтез стерола в различных бактериях» . Границы микробиологии . 7 : 990. дои : 10.3389/fmicb.2016.00990 . ПМЦ 4919349 . ПМИД 27446030 .
- ^ Хосино Ю., Гоше Э.А. (июнь 2021 г.). «Эволюция бактериального биосинтеза стероидов и его влияние на эукариогенез» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (25): e2101276118. Бибкод : 2021PNAS..11801276H . дои : 10.1073/pnas.2101276118 . ПМЦ 8237579 . ПМИД 34131078 .
- ^ Иссон Т.Т., Лав Г.Д., Дюпон К.Л. и др. (июнь 2018 г.). «Отслеживание достижения эукариотами экологического доминирования с помощью изотопов цинка» . Геобиология . 16 (4): 341–352. Бибкод : 2018Gbio...16..341I . дои : 10.1111/gbi.12289 . ПМИД 29869832 .