Гибридный геном эукариот
| Глоссарий |
|---|
Гибридные геномы эукариот возникают в результате межвидовой гибридизации , при которой близкородственные виды спариваются и производят потомство со смешанными геномами . Появление крупномасштабного геномного секвенирования показало, что гибридизация широко распространена и может представлять собой важный источник новых вариаций . Хотя большинство межвидовых гибридов бесплодны или менее приспособлены, чем их родители, некоторые из них могут выживать и размножаться, обеспечивая перенос адаптивных вариантов через границу вида и даже приводя к образованию новых эволюционных линий . Существует два основных варианта геномов гибридных видов: аллополиплоидные , имеющие по одному полному набору хромосом от каждого родительского вида, и гомоплоидные , представляющие собой мозаику геномов родительских видов без увеличения числа хромосом.
Создание гибридных видов требует развития репродуктивной изоляции от родительских видов. Аллополиплоидные виды часто имеют сильные внутренние репродуктивные барьеры из-за различий в числе хромосом , а гомоплоидные гибриды могут оказаться репродуктивно изолированными от родительских видов из-за ряда генетической несовместимости. Однако оба типа гибридов могут стать еще более репродуктивно изолированными, приобретая внешние изоляционные барьеры за счет использования новых экологических ниш по отношению к своим родителям. Гибриды представляют собой слияние расходящихся геномов и, таким образом, сталкиваются с проблемами, возникающими из-за несовместимых комбинаций генов. Таким образом, гибридные геномы очень динамичны и могут подвергаться быстрым эволюционным изменениям, включая стабилизацию генома , при которой отбор против несовместимых комбинаций приводит к фиксации совместимых комбинаций блоков предков внутри гибридных видов. Потенциал быстрой адаптации или видообразования делает гибридные геномы особенно интересной темой для изучения. эволюционная биология . В статье резюмируется, как могут образовываться интрогрессивные аллели или гибридные виды и как развиваются полученные гибридные геномы.
Фон
[ редактировать ]Генетический обмен между видами может препятствовать эволюции биоразнообразия, поскольку поток генов между расходящимися видами противодействует их дифференциации, а гибридизация между недавно разошедшимися видами может привести к потере генетических адаптаций или слиянию видов. [1] Традиционно зоологи рассматривали межвидовую гибридизацию как неадаптивное поведение. [2] что может привести к разрушению коадаптированных генных комплексов . [3] Напротив, биологи растений сразу же осознали, что гибридизация иногда может быть важной эволюционной силой, способствующей увеличению биоразнообразия. [4] [5] В последнее время накапливаются данные, показывающие, что гибридизация также является важным эволюционным процессом у животных. [1] [6] [7] Межвидовая гибридизация может обогатить генетическое разнообразие интрогрессированного таксона, привести к интрогрессии полезных генетических вариаций или даже создать новые гибридные виды. [1]
Сейчас также известно, что гибридизация способствует развитию эволюционного потенциала в нескольких хрестоматийных примерах адаптивной радиации , включая галапагосских вьюрков Geospiza , [8] Африканские цихлиды , [9] Геликониусы бабочки [10] [11] [12] и гавайские мадинские тарвиды и серебряные мечи. [13] В данной статье рассмотрены эволюционные результаты межвидовой гибридизации и свойства геномов гибридных геномов. Многие из обсуждаемых тем также применимы к гибридизации между разными подвидами или популяциями одного и того же вида, но в этой статье основное внимание уделяется межвидовой гибридизации (в этом обзоре называемой гибридизацией).
Эволюционные результаты
[ редактировать ]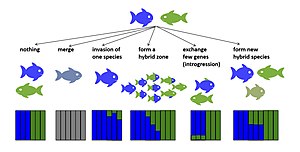
Есть несколько потенциальных эволюционных результатов гибридизации. Если гибриды раннего поколения нежизнеспособны или стерильны, гибридизация может снизить репродуктивный успех родительского вида. [14] [15] Потенциально это может привести к подкреплению , отбору для усиления изоляции перед сматыванием. [16] или если этот вид не сможет развить изоляцию перед спариванием, это может увеличить риск их исчезновения из-за напрасных репродуктивных усилий. [14] Если приспособленность гибридов раннего поколения отлична от нуля, а у некоторых гибридов более поздних поколений приспособленность так же высока или даже выше, чем приспособленность одного или обоих родительских таксонов, гибриды могут вытеснить родительские таксоны, и гибридизующиеся таксоны могут слиться (обратное видообразование). [17] [18] ). Если приспособленность гибридов ранних поколений снижена, но не равна нулю, в зоне контакта таксонов могут возникнуть гибридные зоны. [19] Если гибриды плодовиты, гибридизация может внести вклад в новые вариации за счет обратного скрещивания редких гибридов с родительскими видами. Такая интрогрессивная гибридизация может позволить нейтральным или селективно полезным аллелям переноситься через границы видов даже в парах видов, которые остаются различными, несмотря на случайный поток генов . [20] [21] Пригодность гибрида может меняться в зависимости от времени расхождения между гибридизующимися таксонами. Эта закономерность была продемонстрирована для множества таксонов, включая Drosophila , [22] птицы [23] и рыба. [24] Гибридная приспособленность также может различаться в зависимости от поперечного направления. [25] между гибридами первого и более поздних поколений, [26] и среди людей внутри поколений одного и того же перекрестного типа. [27] [28] В некоторых случаях гибриды могут эволюционировать в новые гибридные виды с репродуктивной изоляцией от обоих родительских таксонов. [29] [30] Ниже описаны эволюционные результаты гибридизации, приводящие к появлению устойчивых гибридных геномов.
Адаптивная интрогрессия
[ редактировать ]При обратном скрещивании редких гибридов с родительскими видами аллели, кодирующие признаки, полезные для обоих родительских видов, могут передаваться через границы видов, даже если родительские виды остаются отдельными таксонами. Этот процесс называется адаптивной интрогрессией (несколько вводящий в заблуждение термин, поскольку само по себе обратное скрещивание может и не быть адаптивным, но некоторые из интрогрессивных вариантов могут быть полезными). [1] ). Моделирование показывает, что адаптивная интрогрессия возможна, если гибридная приспособленность не будет существенно снижена. [31] [32] или адаптивные локусы тесно связаны с вредными. [33] Примеры адаптивных признаков, которые были переданы посредством интрогрессии, включают ген устойчивости к инсектицидам, который был перенесен от Anopheles gambiae к A. coluzzii. [21] и признак красной предупреждающей окраски крыльев у бабочек Heliconius , который находится в процессе естественного отбора от хищников и был интрогрессирован, например, от H. melpomene к H. timareta. [34] и другие виды Heliconius . [20] У растения Arabidopsis arenosa некоторые аллели, отвечающие за адаптацию к засухе и фитотоксичным уровням металлов, были интрогрессированы от A. lyrata . [35] Даже у людей имеются доказательства адаптивной интрогрессии, например, аллелей иммунитета, аллелей пигментации кожи и аллелей, обеспечивающих адаптацию к высокогорной среде обитания у неандертальцев и денисовцев. [36] Если признаки, важные для распознавания видов или репродуктивной изоляции, проникают в популяцию другого вида, интрогрессированная популяция может стать репродуктивно изолированной от других популяций того же вида. Примеры этого включают бабочек Heliconius , у которых происходит избирательная интрогрессия генов рисунка крыльев между разошедшимися линиями. [37] а рисунок крыльев способствует репродуктивной изоляции у некоторых пар видов с низким (например, между H. t. florencia и H. t. linaresi ) и промежуточным уровнем (например, H. c. galanthus / H. pachinus ) дивергенции. [38]
Обнаружение и изучение с помощью геномных инструментов
[ редактировать ]Многие эмпирические тематические исследования начинаются с поискового обнаружения предполагаемых гибридных таксонов или особей с помощью подходов геномной кластеризации, таких как те, которые используются в программном обеспечении STRUCTURE. [39] ПРИМЕСЬ [40] или тонкаяСТРУКТУРА. [41] Эти методы выводят из данных указанное пользователем количество генетических групп и относят каждого человека к одной или нескольким из этих групп. Их можно применять к близкородственным таксонам без необходимости предварительного отнесения особей к таксонам и, таким образом, они могут быть особенно полезны при изучении близкородственных таксонов или комплексов видов. Однако неравномерная выборка родительских таксонов или различная степень дрейфа включенных таксонов могут привести к ошибочным выводам о доказательствах гибридизации. [42]
Если доступны геномные данные нескольких видов, филогенетические методы могут лучше подойти для выявления интрогрессии. Интрогрессивная гибридизация приводит к образованию генных деревьев, которые не согласуются с деревом видов, в результате чего интрогрессивные особи филогенетически ближе к источнику интрогрессии, чем к своим неинтрогрессивным сородичам. Такие противоречивые генные деревья могут также возникнуть случайно из-за неполной сортировки линий, особенно если сравниваемые виды еще молоды. Следовательно, несогласованные генные деревья являются свидетельством интрогрессии только в том случае, если генное дерево, полученное в результате избыточного совместного использования аллелей между гибридизующимися таксонами, сильно перепредставлено по сравнению с альтернативными несогласованными генными деревьями. Для обнаружения такого избыточного обмена аллелями между гибридизующимися таксонами был разработан целый набор методов, включая статистику D Паттерсона или тесты ABBA-BABA. [43] [44] [45] или f-статистика. [46] [47] Модифицированные версии этих тестов можно использовать для определения интрогрессивных областей генома. [48] направление потока генов [49] [50] или количество потока генов. [47]
Для наборов данных с большим количеством таксонов может быть сложно вычислить все возможные тесты гибридизации. В таких случаях лучше подойдут методы построения графов. [51] [52] [53] Эти методы реконструируют сложные филогенетические модели с помощью гибридизации, которые лучше всего соответствуют генетическим связям между выбранными таксонами и дают оценки дрейфа и интрогрессии. Другие методы филогенетической сети, учитывающие неполную сортировку и гибридизацию линий, также могут помочь. [54] [55] Методы, основанные на распаде неравновесия по сцеплению, или методы, предполагающие наличие наследственных путей, могут использоваться для датировки недавних событий примеси или интрогрессии, поскольку с течением времени родовые пути постоянно разрушаются в результате рекомбинации. [52] [56] [57] [58] [59] По мере усиления стабилизации генома люди должны меньше различаться по местному происхождению. Таким образом, уровни стабилизации генома можно оценить путем расчета пропорций предков (например, с помощью fd) в геномных окнах и проверки, коррелируют ли они между людьми. Кроме того, если гибридизация все еще продолжается, пропорции предков должны различаться у разных людей и в космосе.
Другой подход заключается в использовании демографического моделирования для упрощения эволюционной истории изучаемых таксонов. [60] Демографическое моделирование следует применять только к небольшим наборам таксонов, поскольку с увеличением количества таксонов сложность модели увеличивается, а количество параметров модели, таких как время, количество и направление потока генов, а также размеры популяций и время разделения, может быстро стать слишком большим. Соответствие демографических моделей данным можно оценить с помощью частотного спектра сайта. [61] [62] или со сводной статистикой в рамках приближенных байесовских вычислений. [63] Также возможно получить больше мощности, объединив информацию о закономерностях распада неравновесия по сцеплению и частотном спектре аллелей. [64]
Определение гибридных видов
[ редактировать ]Одним из потенциальных эволюционных результатов гибридизации является создание новой, репродуктивно изолированной линии, т. е. гибридного видообразования. [1] [29] Гибридный вид имеет смешанный геном и образует стабильные генетически различные популяции. [29] Некоторые исследователи утверждают, что доказательства наличия гибридизационной основы репродуктивной изоляции должны быть дополнительным определяющим критерием гибридного видообразования. [65] но см. Moharrek et al. [66] Это более строгое определение включает таксоны полиплоидных гибридов, но охватывает лишь несколько хорошо изученных случаев гомоплоидного гибридного видообразования, например Heliconius heurippa , [10] [11] [12] воробей Италии [28] и три Helianthus. вида подсолнечника [67] потому что для большинства предполагаемых примеров гомоплоидного гибридного видообразования генетическая основа репродуктивной изоляции до сих пор неизвестна. [65]
Гибридные виды могут занимать экологическую нишу, отличную от родительских, и могут быть изолированы от родительских видов прежде всего через барьеры перед спариванием (гибридное видообразование с внешними барьерами). [68] ). Гибридные виды также могут быть репродуктивно изолированы от родительских видов путем сортировки несовместимостей, что приводит к новым комбинациям родительских аллелей, несовместимым с обоими родительскими видами, но совместимым внутри гибридного таксона (рекомбинационное гибридное видообразование). [29] Рекомбинационный гибридный таксон обычно также имеет значительную часть генома, полученного от донора интрогрессированного материала, хотя различия существуют как между таксонами, так и внутри линий гибридных таксонов. [69] [70]
Гомоплоидное и полиплоидное гибридное видообразование.
[ редактировать ]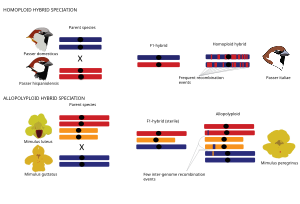
В общем, гибридные виды могут возникнуть в результате двух основных типов гибридного видообразования, которые определяются тем, связано ли событие видообразования с дупликацией генома ( полиплоидия ) или нет. Гомоплоидное гибридное видообразование определяется как эволюция нового гибридного вида с репродуктивной изоляцией от обоих родительских таксонов без изменения плоидности, то есть количества наборов хромосом. [1] Геномы гомоплоидных гибридных видов представляют собой мозаику родительских геномов, поскольку родословные от родительских видов разрушаются в результате рекомбинации . [66] [67] [71] [72] [73] [74] [70] В случае полиплоидного гибридного видообразования гибридизация связана с дупликацией генома, в результате чего образуется аллополиплоид с повышенной плоидностью по сравнению с их родительскими таксонами. В отличие от аллополиплоидов, аутополиплоиды характеризуются дупликацией генома внутри одного вида и поэтому далее в контексте данного обзора не обсуждаются. Аллополиплоидное видообразование чаще встречается у растений, чем у животных. [75] Полиплоидные гибриды можно мгновенно изолировать от родительских видов за счет различий в числе хромосом . [75]
Репродуктивная изоляция от родительских видов
[ редактировать ]Для успешного создания гибридного вида необходима достаточная репродуктивная изоляция от обоих родительских видов. [1] [65] [76] Репродуктивной изоляции от родительских видов труднее достичь для гомоплоидных гибридов, у которых различия кариотипа не способствуют внутренней изоляции. Репродуктивная изоляция между гибридным видом и его родительским видом может возникать из-за множества репродуктивных барьеров до или после оплодотворения (презиготических или постзиготических соответственно), которые сами по себе могут зависеть или независимы от условий окружающей среды (внешние или внутренние барьеры соответственно). [77] Например, внутренние постзиготические барьеры вызывают нежизнеспособность или стерильность гибридов независимо от среды, в которой они возникают, тогда как внешние постзиготические барьеры приводят к появлению гибридов с низкой приспособленностью из-за дезадаптации к конкретной среде. [30]
Также было показано, что презиготические внутренние и внешние различия важны для изоляции гибридов от их родительских видов. У растений изоляция, опосредованная опылителями, возникающая в результате изменений характеристик цветков, может быть важным внешним презиготическим экологическим барьером. [78] [79] [80] [81] Было показано, что сильная внешняя презигота изолирует гибридный вид Senecio eboracensis от его родительского вида, где гибриды практически отсутствуют в дикой природе, хотя в лабораторных экспериментах часть гибридного потомства оказывается плодовитой. [82] Лоу и Эбботт приходят к выводу, что самоопыление , время цветения и признаки, участвующие в привлечении опылителей, вероятно, способствуют этой внешней изоляции. [82] В нескольких таксонах также сообщалось о презиготической изоляции, обусловленной предпочтением партнера, возникающей в результате внутреннего ассортативного спаривания между гибридами. У африканских цихлид экспериментальные гибриды продемонстрировали комбинации родительских признаков и предпочтений, в результате чего гибриды преимущественно спаривались с другими гибридами. [83] Похожая картина была обнаружена у галапагосских вьюрков Geospiza , у которых особая гибридная песня возникла в результате трансгрессивной морфологии клюва. [8] а гибридные бабочки Heliconius предпочли гибридный рисунок крыльев таковому у обоих родительских видов. [12] Внутренние различия в использовании среды обитания [84] или в фенологии [85] может привести к некоторой степени репродуктивной изоляции от родительских видов, если спаривание зависит от времени и среды обитания. Например, раса яблочных хозяев у личинок Rhagoletis pomonella развилась после интрогрессии генов, связанных с диапаузой, от мексиканских мух altiplano, что позволило переключиться с предкового хозяина боярышника на яблоню с более поздним цветением. [86] [87] и изолировали две расы хозяев посредством аллохронной внутренней презиготической изоляции. У Xiphophorus ассортативное спаривание с сильными предками поддерживало отдельный гибридный генетический кластер в течение 25 поколений, но исчезло в манипулируемых условиях. меченосцев [88] Следовательно, презиготические репродуктивные барьеры на пути потока генов могут зависеть от окружающей среды.
Также было показано, что постзиготические изолирующие барьеры важны для различных гибридных линий. Работа над подсолнечником Helianthus показала, что внутренние постзиготические факторы могут вызывать репродуктивную изоляцию от родительских видов. Постзиготические барьеры заключаются в уже существующих структурных различиях, [73] [89] в сочетании с гибридизацией индуцировали структурные различия. [73] Сортировка несовместимостей между родительскими видами, когда одно из них изолирует гибридный таксон от одного родителя, а другое подмножество изолирует его от другого родителя, привело к внутренней постзиготической изоляции между итальянским воробьем Passer italiae и его родительским видом. [28] Моделирование показывает, что вероятность гибридного видообразования посредством этого механизма зависит от времени расхождения между родительскими видами. [90] размер популяции гибридного вида, [91] характер отбора, действующего на гибриды, и связь между несовместимостями друг с другом и с адаптивными вариантами. [92] Внешние экологические барьеры против родительских видов могут возникнуть как побочные продукты экологической дифференциации, если спаривание зависит от времени и/или среды обитания. Было показано, что гибридные виды адаптируются к новым экологическим нишам посредством трансгрессивных фенотипов . [84] или посредством новых комбинаций экологических признаков родительского вида, [93] а экологический отбор против гибридных фенотипов родительского гибрида приведет к внешней постзиготической изоляции.
Стабилизация
[ редактировать ]
Гибридизация может иметь много разных результатов. Гибридное видообразование приводит к репродуктивной изоляции как от родительских видов, так и от геномов, которые развиваются независимо от геномов родительских видов. Интрогрессивная гибридизация может перенести важные новые варианты в геномы видов, которые остаются отличными от других таксонов, несмотря на периодический поток генов. В этой статье оба типа геномов, полученных в результате гибридизации, называются персистентными гибридными геномами. После первоначальной гибридизации интрогрессионные пути, генетические блоки, унаследованные от каждого родительского вида, разрушаются в результате последовательных поколений и событий рекомбинации. Рекомбинация чаще встречается в гомоплоидных гибридных геномах, чем в аллополиплоидных гибридных геномах. У аллополиплоидов рекомбинация может дестабилизировать кариотип и привести к аберрантному мейотическому поведению и снижению фертильности, но также может генерировать новые комбинации генов и полезные фенотипические признаки. [94] как у гомоплоидных гибридов. Как только гибридизация между гибридным таксоном и его родительскими таксонами прекращается, различные блоки предков или пути интрогрессии могут фиксироваться - процесс, называемый «стабилизацией генома». [71] Некоторые пути интрогрессии удаляются путем отбора против несовместимости, а другие фиксируются. Теоретические модели гибридных зон предполагают, что разрушение блоков предков посредством рекомбинации подавляется вблизи генов, обеспечивающих репродуктивную изоляцию, из-за более низкой приспособленности рекомбинантных гибридов. [95] На силу подавления влияют форма отбора, доминирование и расположение локуса на аутосоме или половой хромосоме . [95] Время стабилизации генома варьируется. Было обнаружено, что фиксация предковых блоков происходит быстро в геномах экспериментальных гибридных видов подсолнечника Helianthus . [96] а стабилизация генома гибридных видов подсолнечника, по оценкам, займет сотни поколений. [71] Геномы грибов Zymoseptoria . стабилизировались примерно в течение ок 400 поколений, [97] тогда как в гибридных Xiphophorus геномах меченосца [98] стабилизация генома не была достигнута до тех пор, пока ок. 2000 и 2500 поколений. Лишь немногие неандертальские регионы закрепились в геномах человека на протяжении ок. 2000 поколений после гибридизации, [99] и сегрегирующая несовместимость присутствует у гибридного итальянского воробья примерно через 5000 поколений после первоначального события гибридизации. [100]
Со временем генетический дрейф в конечном итоге стохастически зафиксирует блоки, полученные от двух родительских видов, в конечных изолированных гибридных популяциях. [71] Отбор против локусов несовместимости может ускорить процесс фиксации родительских аллелей, поскольку гибриды, обладающие аллелями, которые с меньшей вероятностью вызывают несовместимость, будут иметь более высокую приспособленность, и благоприятные аллели будут распространяться в популяции. Однако фиксация рецессивных слабовредных аллелей в родительских таксонах может также привести к тому, что гибриды сохранят обе родительские аллели: поскольку гибриды с гаплотипами от обоих родителей не являются гомозиготными ни по каким слабовредным аллелям, они имеют более высокую приспособленность, чем гибриды только с одним родительским гаплотипом. . Это ассоциативное преобладание , [101] [102] может замедлить процесс фиксации родительских аллелей, способствуя сохранению обоих родительских гаплотипов. Эффект ассоциативного доминирования наиболее силен в областях низкой рекомбинации, включая инверсии. [103] Баланс между аллелями и аллельными комбинациями, обеспечивающими благоприятные фенотипические признаки, и сила отбора против несовместимости определяют, какие интрогрессивные пути будут унаследованы от каких родительских видов при гибридизации. [21] [104] [105] Область устойчивости к инсектицидам сохранилась после гибридизации у Anopheles coluzzi . [21] предполагая роль отбора в поддержании благоприятных интрогрессивных регионов. Скорость локальной рекомбинации важна для вероятности интрогрессии, поскольку в случае широко распространенной несовместимости интрогрессированные аллели с большей вероятностью рекомбинируют из несовместимостей в областях с высокой рекомбинацией. Эта закономерность была обнаружена у обезьяньих цветов Mimulus . [106] в Mus Domesticus , доме мышей [107] в Геликониуса бабочках [105] и у рыбы-меченосца Xiphophorus . [69]
рыб выявлена полногеномная несовместимость У мечефорных . [108] химерные гены и мутации ортологичных генов вызывают несовместимость у экспериментальных гибридов золотых рыбок Cyprinidae и карпа ранних поколений [109] и обнаружено, что ключевую роль играет мито-ядерная несовместимость, например, у итальянских воробьев, [70] [110] грибок [111] и цитоядерная несовместимость у Mimulus . растений [112] Данные об измененных паттернах экспрессии у синтетических гибридов и отсутствующих комбинациях генов у гибридных видов также позволяют предположить, что репарация ДНК [70] [109] [113] и гены, участвующие в мутагенезе и путях, связанных с раком [109] может вызвать несовместимость у гибридов. Формирование генома гибридных видов определяется отбором несовместимых комбинаций. [69] [98] [104]
Измененные свойства генома
[ редактировать ]Гибридное происхождение может влиять на структуру и свойства генома. Было доказано, что он увеличивает частоту мутаций . [77] [114] [115] для активации мобильных элементов, [116] [117] [118] и вызывать хромосомные перестройки . [119] [120] Повышенная активация транспозонов , как предполагает теория «геномного шока» МакКлинтока, может привести к изменениям в экспрессии генов. Мобильные элементы могут, помимо изменения генных продуктов, если они вставлены в ген, также изменять активность промотора для генов, если они вставлены выше кодирующих областей, или могут вызывать молчание генов в результате разрушения генов. [121] [122] Для аллополиплоидных геномов хромосомные перестройки могут быть результатом «геномного шока», вызванного гибридизацией, при этом более отдаленно родственные виды более склонны к реорганизации генома, например, у Nicotiana . [123] Хромосомные перестройки, возникающие в результате геномного шока или событий рекомбинации между негомологичными субгеномами, могут привести к увеличению или уменьшению размеров генома. [124] Как увеличение, так и снижение были обнаружены у рода Nicotiana и не были связаны с возрастом после гибридизации. [125]
После дупликации генома у аллополиплоидов геном подвергается диплоидизации , то есть процессу, в котором геном перестраивается, чтобы действовать как мейотический диплоид. [126] [127] После такой диплоидизации большая часть генома теряется из-за фракционирования генома, потери функции того или иного из вновь дуплицированных генов. [127] [128] В ходе метаанализа Санкофф и его коллеги обнаружили доказательства, соответствующие устойчивым к редукции парам и концентрации функциональных генов на одной хромосоме, и предполагают, что процесс редукции частично ограничен. [128]
Родственное аллополиплоидное специфическое явление - доминирование субгенома. Например, у октоплоидной земляники Fragaria один из четырех субгеномов является доминантным и имеет значительно большее генное содержание, его гены чаще экспрессируются, а обмены между гомологичными хромосомами смещены в пользу этого субгенома по сравнению с остальными субгеномами. [129] Это исследование также показало, что определенные черты, например устойчивость к болезням, в высокой степени контролируются доминантным субгеномом. [129] Предлагаемый механизм возникновения доминирования субгенома предполагает, что относительное доминирование связано с плотностью мобильных элементов в каждом субгеноме. Субгеномы с более высокой плотностью мобильных элементов имеют тенденцию вести себя покорно по отношению к другим субгеномам, когда они объединяются в аллополиплоидный геном. [127] [130] Интересно, что субгеномное доминирование может возникнуть сразу же у аллополиплоидов, как показано у синтетических и недавно появившихся обезьяньих цветов. [130]
В дополнение к этим изменениям в структуре и свойствах генома исследования аллополиплоидного риса и сига предполагают, что у гибридных видов могут быть нарушены паттерны экспрессии генов. [131] [132] Исследования синтетических и природных аллополиплоидов Tragopogon miscellus показывают, что экспрессия генов менее строго регулируется непосредственно после гибридизации и что новые модели экспрессии возникают и стабилизируются в течение 40 поколений. [133] В то время как вариация экспрессии микроРНК изменяет экспрессию генов и влияет на рост в естественном аллополиплоидном Arabidopsis suecica и экспериментальных линиях, наследование siРНК стабильно и поддерживает стабильность хроматина и генома. [134] потенциально буферизующий против транскриптомного шока.
Факторы, влияющие на формирование и стойкость
[ редактировать ]Хотя гибридизация необходима для создания устойчивых гибридных геномов, ее недостаточно. Для сохранения гибридных геномов у гибридных видов они должны быть достаточно репродуктивно изолированы от родительских видов, чтобы избежать слияния видов. Отбор интрогрессивных вариантов позволяет сохранять гибридные геномы в интрогрессивных линиях. Частота гибридизации, жизнеспособность гибридов и легкость возникновения репродуктивной изоляции от родительских видов или сила отбора для поддержания интрогрессивных регионов являются, следовательно, факторами, влияющими на скорость формирования стабильных гибридных линий.
Можно сделать несколько общих выводов об относительной распространенности гибридизации, поскольку выборка распределена неравномерно, даже если есть свидетельства гибридизации во все большем числе таксонов. Одна из закономерностей заключается в том, что гибридизация чаще встречается у растений, где она встречается у 25% видов, тогда как у животных она встречается только у 10%. [135] У большинства растений, а также у многих групп животных гетероморфные половые хромосомы отсутствуют. [136] Отсутствие гетероморфных половых хромосом приводит к более медленному накоплению репродуктивной изоляции. [137] [138] и, следовательно, может обеспечить гибридизацию между филогенетически более отдаленными таксонами. Правило Холдейна [139] утверждает, что «когда у потомства F1 двух разных рас животных один пол отсутствует, редок или бесплоден, этот пол является гетерозиготным». Эмпирические данные подтверждают роль гетероморфных половых хромосом в стерильности и нежизнеспособности гибридов. Тесно связанное наблюдение - это сильный X-эффект, свидетельствующий о непропорциональном вкладе X/Z-хромосомы в снижение приспособленности гетерогаметных гибридов. [22] Эти закономерности, вероятно, возникают из-за того, что рецессивные аллели с вредными эффектами у гибридов оказывают более сильное влияние на гетерогаметный, чем гомогаметный пол, из-за гемизиготной экспрессии. [140] У таксонов с хорошо дифференцированными половыми хромосомами правило Холдейна оказалось близким к универсальному, а гетероморфные половые хромосомы демонстрируют пониженную интрогрессию по X в XY. [141] В соответствии с ролью гетероморфных половых хромосом в ограничении формирования гибридного генома повышенная дифференциация половых хромосом наблюдалась как в системах ZW, так и в XY. [142] Эта закономерность может отражать меньший эффективный размер популяции и более высокую склонность к дрейфу половых хромосом. [143] повышенная частота локусов, участвующих в репродуктивной изоляции [144] и/или повышенный конфликт половых хромосом. [145] Результаты отбора по однородительскому наследованию, например, мито-ядерных локусов, расположенных на Z-хромосоме у гибридных итальянских воробьев [70] согласуется с тем, что совместимые половые хромосомы важны для формирования жизнеспособных гибридных геномов.
Есть также несколько экологических факторов, влияющих на вероятность гибридизации. В целом гибридизация чаще наблюдается у видов с внешним оплодотворением, включая растения, а также рыб, чем у клад с внутренним оплодотворением. [4] У растений высокая скорость самоопыления у некоторых видов может препятствовать гибридизации, а система размножения также может влиять на частоту гетероспецифичного переноса пыльцы. [146] [147] У грибов гибриды могут образовываться путем амейотического слияния клеток или гиф. [148] помимо механизмов, доступных растениям и животным. Такое слияние вегетативных клеток и последующее парасексуальное спаривание с митотическим кроссинговером может привести к образованию рекомбинированных гибридных клеток. [148]
Для развития гибридных видов необходима репродуктивная изоляция от родительских видов. Таким образом, легкость, с которой возникает такая репродуктивная изоляция, также важна для скорости возникновения стабильных гибридных видов. Полиплоидизация и асексуальность являются механизмами, которые приводят к мгновенной изоляции и могут увеличить скорость образования гибридной линии. Способность к самоопылению может также способствовать стабилизации аллополиплоидных таксонов, обеспечивая совместимого партнера (самого себя) на ранних стадиях видообразования аллополиплоидов, когда редкие цитотипы находятся в невыгодном репродуктивном положении из-за межцитотипического спаривания. [149] Согласно моделированию, самоопыление также повысит вероятность появления гомоплоидных гибридов. [150] а более высокая вероятность самоопыления может способствовать более высокой частоте гибридных видов у растений. Гибридизация грибов может привести к появлению бесполых гибридных видов, таких как грибы Epichloe , где виды гибридов являются бесполыми, а негибриды включают как бесполые, так и половые виды. [151] Гибридизация между сильно расходящимися таксонами животных может также привести к появлению бесполых гибридных видов, как показано, например, у европейских щиповок, Cobitis , [152] и большинство, если не все, бесполых видов позвоночных имеют гибридное происхождение. [153] Интересно, что арктическая флора содержит необычно высокую долю аллополиплоидных растений. [154] предполагая, что эти гибридные таксоны могут иметь преимущество в экстремальных условиях, потенциально за счет уменьшения негативных последствий инбридинга. Следовательно, как геномная архитектура, так и экологические свойства могут влиять на вероятность образования гибридных видов.
Для интрогрессивных таксонов сила отбора интрогрессивных вариантов решает, будут ли интрогрессированные участки распространяться в популяции и будут ли формироваться стабильные интрогрессивные геномы. Было показано, что сильный отбор на устойчивость к инсектицидам увеличивает интрогрессию аллеля устойчивости Anopheles gambiae к A. coluzzi . малярийным комарам [155] У бабочек Heliconius сильный отбор на наличие локально обильных рисунков окраски крыльев неоднократно приводил к фиксации аллелей, которые интрогрессировали от локально адаптированных бабочек к новым колонизирующим видам или подвидам. [34] Шансы на фиксацию полезных интрогрессивных вариантов зависят от типа и силы отбора интрогрессивного варианта и связи с другими интрогрессивными вариантами, против которых происходит отбор.
Факторы, влияющие на пораженные гены и геномные регионы
[ редактировать ]Генетический обмен может происходить между популяциями или зарождающимися видами, дивергирующими в географической близости, или между дивергентными таксонами, вступающими в вторичный контакт . Ожидается, что гибридизация между более дивергентными линиями будет иметь больший потенциал для внесения полезных аллелей или создания новизны, чем гибридизация между менее дивергентными популяциями, поскольку более дивергентные аллели объединяются и, таким образом, с большей вероятностью будут иметь большой эффект приспособленности для создания трансгрессивных фенотипов. [156] Гибридизация между более разнородными линиями также с большей вероятностью приведет к образованию несовместимых комбинаций аллелей, что снижает первоначальную гибридную приспособленность. [157] но потенциально также способствует гибридному видообразованию, если их сортировать взаимно, как описано выше. [156] Таким образом, промежуточная генетическая дистанция может быть наиболее благоприятной для гибридного видообразования. [156] Экспериментальные лабораторные скрещивания подтверждают эту гипотезу. [90]
Доля генома, унаследованного от реципиента интрогрессированного материала, сильно различается среди видов и внутри них. После первоначального события гибридизации представительство во многих полиплоидных таксонах составляет 50%, хотя копии родительских генов последовательно теряются и могут искажать вклад в один мажоритарный родительский геном. [158] Относительно равный родительский вклад также обнаружен у некоторых гомоплоидных гибридных видов. [74] но в других случаях они крайне неравны, как, например, у некоторых видов Heliconius . [159] Большинство предков может даже происходить от донора интрогрессированного материала, как было показано для Anopheles gambiae . комаров [160] Интересно, что внутри гибридного вида также могут быть различия во вкладе родителей. И у меченосцев, и у итальянских воробьев существуют популяции, которые сильно различаются по пропорциям родительских геномов, которые они унаследовали. [69] [70]
Паттерны интрогрессии могут сильно различаться в зависимости от генома, даже на коротких хромосомных расстояниях. Примеры адаптивной интрогрессии четко определенных областей включают инверсную область, содержащую гены, участвующие в устойчивости к инсектицидам. [21] а интрогрессия дивергентного инвертированного хромосомного сегмента привела к появлению « супергена », который кодирует полиморфизм мимикрии у бабочки Heliconius numata . [161] Эти данные согласуются с моделями, предполагающими, что геномные перестройки важны для соединения локально адаптивных локусов. [162] Гены и геномные области, являющиеся адаптивными, могут легко интрогрессировать между видами, например, в гибридных зонах, если они не связаны с локусами несовместимости. Часто это относится к полупроницаемым границам видов, [19] [163] [164] и примеры включают, например, гены, участвующие в обонянии, которые интрогрессируют через гибридную зону Mus musculus и M. Domesticus . [165] В гибридных зонах с преимущественно проницаемыми границами видов структура интрогрессивных регионов позволяет определить, какие геномные регионы участвуют в несовместимости и репродуктивной изоляции. [166]
Ссылки
[ редактировать ]![]() Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2019 ) ( отчеты рецензента ): Анна Рунемарк; Марио Вальехо-Марин; Джоанна I Мейер (27 ноября 2019 г.). «Гибридные геномы эукариот» . ПЛОС Генетика . 15 (11):e1 doi : 10.1371/JOURNAL.PGEN.1008404 . ISSN 1553-7390 . ПМК 6880984 . ПМИД 31774811 . Викиданные Q8
Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2019 ) ( отчеты рецензента ): Анна Рунемарк; Марио Вальехо-Марин; Джоанна I Мейер (27 ноября 2019 г.). «Гибридные геномы эукариот» . ПЛОС Генетика . 15 (11):e1 doi : 10.1371/JOURNAL.PGEN.1008404 . ISSN 1553-7390 . ПМК 6880984 . ПМИД 31774811 . Викиданные Q8
- ^ Jump up to: а б с д и ж г Эбботт Р., Альбах Д., Анселл С., Арнтцен Дж.В., Бэрд С.Дж., Бирн Н. и др. (февраль 2013 г.). «Гибридизация и видообразование» . Журнал эволюционной биологии . 26 (2): 229–46. дои : 10.1111/j.1420-9101.2012.02599.x . ПМИД 23323997 . S2CID 830823 .
- ^ Фишер Р.А. (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон Пресс. дои : 10.5962/bhl.title.27468 .
- ^ Майр Э (1963). Виды животных и эволюция . Кембридж, Массачусетс и Лондон, Англия: Издательство Гарвардского университета. doi : 10.4159/harvard.9780674865327 . ISBN 9780674865327 .
- ^ Jump up to: а б Стеббинс Г.Л. (1959). «Роль гибридизации в эволюции». Труды Американского философского общества . 103 (2): 231–251. ISSN 0003-049X . JSTOR 985151 .
- ^ Андерсон Э., Стеббинс Г.Л. (1954). «Гибридизация как эволюционный стимул» . Эволюция . 8 (4): 378–388. дои : 10.1111/j.1558-5646.1954.tb01504.x .
- ^ Арнольд М.Л. (1997). Естественная гибридизация и эволюция . Кэри: Издательство Оксфордского университета. ISBN 9780195356687 . OCLC 960164734 .
- ^ Маллет Дж., Безански Н., Хан М.В. (февраль 2016 г.). «Насколько сетчатыми являются виды?» . Биоэссе . 38 (2): 140–9. doi : 10.1002/bies.201500149 . ПМК 4813508 . ПМИД 26709836 .
- ^ Jump up to: а б Ламичхани С., Хан Ф., Вебстер М.Т., Андерссон Л., Грант Б.Р., Грант PR (январь 2018 г.). «Быстрое гибридное видообразование у вьюрков Дарвина» . Наука . 359 (6372): 224–228. Бибкод : 2018Sci...359..224L . дои : 10.1126/science.aao4593 . ПМИД 29170277 .
- ^ Мейер Дж.И., Маркес Д.А., Мвайко С., Вагнер К.Э., Экскофье Л., Зеехаузен О. (февраль 2017 г.). «Древняя гибридизация способствует быстрой адаптивной радиации цихлидных рыб» . Природные коммуникации . 8 (1): 14363. Бибкод : 2017NatCo...814363M . дои : 10.1038/ncomms14363 . ПМК 5309898 . ПМИД 28186104 .
- ^ Jump up to: а б Маварес Дж., Саласар К.А., Бермингем Э., Сальседо К., Джиггинс К.Д., Линарес М. (июнь 2006 г.). «Видообразование путем гибридизации бабочек Heliconius». Природа . 441 (7095): 868–71. Бибкод : 2006Natur.441..868M . дои : 10.1038/nature04738 . ПМИД 16778888 . S2CID 2457445 .
- ^ Jump up to: а б Салазар С., Бакстер С.В., Пардо-Диас С., Ву Г., Серридж А., Линарес М. и др. (апрель 2010 г.). Уолш Б. (ред.). «Генетические доказательства видообразования гибридных признаков у бабочек Heliconius» . ПЛОС Генетика . 6 (4): e1000930. дои : 10.1371/journal.pgen.1000930 . ПМЦ 2861694 . ПМИД 20442862 .
- ^ Jump up to: а б с Мело MC, Салазар С, Джиггинс CD, Линарес М (июнь 2009 г.). «Ассортативные предпочтения спаривания среди гибридов открывают путь к гибридному видообразованию». Эволюция; Международный журнал органической эволюции . 63 (6): 1660–5. дои : 10.1111/j.1558-5646.2009.00633.x . ПМИД 19492995 . S2CID 17250691 .
- ^ Карлквист С.Дж., Болдуин Б.Г., Карр Г.Д. (2003). Тарвиды и серебряные мечи: эволюция сложноцветных . Сент-Луис Сент-Луис: Издательство Ботанического сада Миссури. ISBN 1930723202 . OCLC 52892451 .
- ^ Jump up to: а б Вольф Д.Е., Такебаяши Н., Ризеберг Л.Х. (2001). «Прогнозирование риска вымирания посредством гибридизации». Биология сохранения . 15 (4): 1039–1053. Бибкод : 2001ConBi..15.1039W . дои : 10.1046/j.1523-1739.2001.0150041039.x . ISSN 0888-8892 . S2CID 86704425 .
- ^ Прентис П.Дж., Уайт Э.М., Рэдфорд И.Дж., Лоу А.Дж., Кларк А.Р. (2007). «Может ли гибридизация вызвать локальное вымирание: случай демографического затопления австралийского аборигена Senecio pinnatifolius инвазивным Senecio madagascariensis?» (PDF) . Новый фитолог . 176 (4): 902–12. дои : 10.1111/j.1469-8137.2007.02217.x . ПМИД 17850249 .
- ^ Серведио М.Р., Нур М.А. (2003). «Роль подкрепления в видообразовании: теория и данные». Ежегодный обзор экологии, эволюции и систематики . 34 (1): 339–364. doi : 10.1146/annurev.ecolsys.34.011802.132412 . ISSN 1543-592X .
- ^ Раймер Дж. М., Симберлофф Д. (1996). «Вымирание путем гибридизации и интрогрессии». Ежегодный обзор экологии и систематики . 27 (1): 83–109. doi : 10.1146/annurev.ecolsys.27.1.83 . ISSN 0066-4162 .
- ^ Зеехаузен О (май 2006 г.). «Сохранение: потеря биоразнообразия из-за обратного видообразования» . Современная биология . 16 (9): Р334-7. Бибкод : 2006CBio...16.R334S . дои : 10.1016/j.cub.2006.03.080 . ПМИД 16682344 . S2CID 17611278 .
- ^ Jump up to: а б Томпсон Дж.Д. (1994). «Харрисон, Р.Г. (ред.). Гибридные зоны и эволюционный процесс. Oxford University Press, Оксфорд. 364 стр. Цена 45,00 фунтов стерлингов». Журнал эволюционной биологии . 7 (5): 631–634. дои : 10.1046/j.1420-9101.1994.7050631.x . ISBN 0-19-506917-Х . ISSN 1010-061X .
- ^ Jump up to: а б Дасмахапатра К.К., Уолтерс-младший, Бриско А.Д., Дэйви Дж.В., Уибли А., Надо, штат Нью-Джерси, и др. (Консорциум Heliconius Genome) (июль 2012 г.). «Геном бабочки демонстрирует беспорядочный обмен мимикриальными адаптациями между видами» . Природа . 487 (7405): 94–8. Бибкод : 2012Natur.487...94T . дои : 10.1038/nature11041 . ПМК 3398145 . ПМИД 22722851 .
- ^ Jump up to: а б с д и Ханемаайер М.Дж., Коллиер Т.С., Чанг А., Шотт К.С., Хьюстон П.Д., Шмидт Х. и др. (декабрь 2018 г.). «Судьба генов, которые пересекают границы видов после крупного события гибридизации в естественной популяции комаров». Молекулярная экология . 27 (24): 4978–4990. Бибкод : 2018MolEc..27.4978H . дои : 10.1111/mec.14947 . ПМИД 30447117 . S2CID 53568503 .
- ^ Jump up to: а б Койн Дж.А., Орр Х.А. (2004). Видообразование . Сандерленд: Sinauer Associates. ISBN 0878930914 . OCLC 55078441 .
- ^ Прайс ТД, Бувье ММ (2002). «Эволюция постзиготической несовместимости F1 у птиц». Эволюция . 56 (10): 2083–9. doi : 10.1554/0014-3820(2002)056[2083:teofpi]2.0.co;2 . ISSN 0014-3820 . ПМИД 12449494 . S2CID 24378606 .
- ^ Стелкенс Р.Б., Янг К.А., Зеехаузен О. (март 2010 г.). «Накопление репродуктивной несовместимости у африканских цихлид» . Эволюция; Международный журнал органической эволюции . 64 (3): 617–33. дои : 10.1111/j.1558-5646.2009.00849.x . ПМИД 19796149 . S2CID 10319450 .
- ^ Реберниг К.А., Лафон-Пласетт С., Хаторанган М.Р., Слотте Т., Кёлер С. (июнь 2015 г.). Бомблис К (ред.). «Невзаимные барьеры межвидовой гибридизации у рода Capsella установлены в эндосперме» . ПЛОС Генетика . 11 (6): e1005295. дои : 10.1371/journal.pgen.1005295 . ПМЦ 4472357 . ПМИД 26086217 .
- ^ Притчард В.Л., Кнутсон В.Л., Ли М., Зиеба Дж., Эдмандс С. (февраль 2013 г.). «Приспособленность и морфологические результаты многих поколений гибридизации копепод Tigriopus Californicus» . Журнал эволюционной биологии . 26 (2): 416–33. дои : 10.1111/jeb.12060 . ПМИД 23278939 . S2CID 10092426 .
- ^ Ризеберг Л.Х., Арчер М.А., Уэйн Р.К. (октябрь 1999 г.). «Трансгрессивная сегрегация, адаптация и видообразование» . Наследственность . 83 (Часть 4) (4): 363–72. дои : 10.1038/sj.hdy.6886170 . ПМИД 10583537 .
- ^ Jump up to: а б с Берк Дж. М., Арнольд М. Л. (2001). «Генетика и приспособленность гибридов». Ежегодный обзор генетики . 35 (1): 31–52. дои : 10.1146/annurev.genet.35.102401.085719 . ПМИД 11700276 . S2CID 26683922 .
- ^ Jump up to: а б с д Маллет Дж. (март 2007 г.). «Гибридное видообразование». Природа . 446 (7133): 279–83. Бибкод : 2007Natur.446..279M . дои : 10.1038/nature05706 . ПМИД 17361174 . S2CID 1016526 .
- ^ Jump up to: а б Вальехо-Марин М., Хискок С.Дж. (сентябрь 2016 г.). «Гибридизация и гибридное видообразование в условиях глобальных изменений» . Новый фитолог . 211 (4): 1170–87. дои : 10.1111/nph.14004 . hdl : 1893/23581 . ПМИД 27214560 .
- ^ Бартон Н., Бенгтссон Б.О. (декабрь 1986 г.). «Барьер для генетического обмена между гибридизующимися популяциями» . Наследственность . 57 (Часть 3) (3): 357–76. дои : 10.1038/hdy.1986.135 . ПМИД 3804765 .
- ^ Демон I, Хакку П., ван ден Бош Ф (сентябрь 2007 г.). «Интрогрессия генов устойчивости между популяциями: модельное исследование устойчивости к инсектицидам у Bemisia tabaci». Теоретическая популяционная биология . 72 (2): 292–304. дои : 10.1016/j.tpb.2007.06.005 . ПМИД 17658572 .
- ^ Юкер Х., Сеттер Д., Хермисон Дж. (июнь 2015 г.). «Адаптивная интрогрессия генов после вторичного контакта» . Журнал математической биологии . 70 (7): 1523–80. дои : 10.1007/s00285-014-0802-y . ПМЦ 4426140 . ПМИД 24992884 .
- ^ Jump up to: а б Пардо-Диас С., Салазар С., Бакстер С.В., Мерот С., Фигейредо-Риди В., Джорон М. и др. (2012). Р. Кронфорст М. (ред.). «Адаптивная интрогрессия через границы видов у бабочек Heliconius» . ПЛОС Генетика . 8 (6): e1002752. дои : 10.1371/journal.pgen.1002752 . ПМК 3380824 . ПМИД 22737081 .
- ^ Арнольд Б.Дж., Ланер Б., ДаКоста Дж.М., Вейсман К.М., Холлистер Дж.Д., Солт Д.Э. и др. (июль 2016 г.). «Заимствованные аллели и конвергенция в змеиной адаптации» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (29): 8320–5. Бибкод : 2016PNAS..113.8320A . дои : 10.1073/pnas.1600405113 . ПМЦ 4961121 . ПМИД 27357660 .
- ^ Расимо Ф., Санкарараман С., Нильсен Р., Уэрта-Санчес Э. (июнь 2015 г.). «Доказательства архаичной адаптивной интрогрессии у людей» . Обзоры природы. Генетика . 16 (6): 359–71. дои : 10.1038/nrg3936 . ПМЦ 4478293 . ПМИД 25963373 .
- ^ Кронфорст М.Р., Папа Р. (май 2015 г.). «Функциональная основа рисунка крыльев бабочек Heliconius: молекулы, лежащие в основе мимикрии» . Генетика . 200 (1): 1–19. дои : 10.1534/genetics.114.172387 . ПМЦ 4423356 . ПМИД 25953905 .
- ^ Меро С., Салазар С., Меррилл Р.М., Джиггинс К.Д., Джорон М. (июнь 2017 г.). «Бабочки Геликониусы» . Слушания. Биологические науки . 284 (1856): 20170335. doi : 10.1098/rspb.2017.0335 . ПМЦ 5474069 . ПМИД 28592669 .
- ^ Причард Дж. К., Стивенс М., Доннелли П. (июнь 2000 г.). «Вывод о структуре популяции с использованием данных мультилокусного генотипа» . Генетика . 155 (2): 945–59. дои : 10.1093/генетика/155.2.945 . ПМК 1461096 . ПМИД 10835412 .
- ^ Александр Д.Х., Новембре Дж., Ланге К. (сентябрь 2009 г.). «Быстрая оценка происхождения неродственных лиц на основе модели» . Геномные исследования . 19 (9): 1655–64. дои : 10.1101/гр.094052.109 . ПМЦ 2752134 . ПМИД 19648217 .
- ^ Лоусон DJ, Хелленталь Дж., Майерс С., Фалуш Д. (январь 2012 г.). Копенхейвер GP (ред.). «Вывод о структуре населения с использованием данных плотных гаплотипов» . ПЛОС Генетика . 8 (1): e1002453. дои : 10.1371/journal.pgen.1002453 . ПМК 3266881 . ПМИД 22291602 .
- ^ Лоусон DJ, ван Дорп Л., Фалуш Д. (август 2018 г.). «Учебное пособие о том, как не переоценивать гистограммы СТРУКТУРЫ и ПРИМЕШЕНИЯ» . Природные коммуникации . 9 (1): 3258. Бибкод : 2018NatCo...9.3258L . дои : 10.1038/s41467-018-05257-7 . ПМК 6092366 . ПМИД 30108219 .
- ^ Кулатинал Р.Дж., Стевисон Л.С., Нур М.А. (июль 2009 г.). Нахман М.В. (ред.). «Геномика видообразования дрозофилы: разнообразие, дивергенция и интрогрессия, оцененные с использованием секвенирования генома с низким охватом» . ПЛОС Генетика . 5 (7): е1000550. дои : 10.1371/journal.pgen.1000550 . ПМК 2696600 . ПМИД 19578407 .
- ^ Грин Р.Э., Краузе Дж., Бриггс А.В., Маричич Т., Стензель У., Кирхер М. и др. (май 2010 г.). «Проект последовательности генома неандертальца» . Наука . 328 (5979): 710–722. Бибкод : 2010Sci...328..710G . дои : 10.1126/science.1188021 . ПМК 5100745 . ПМИД 20448178 .
- ^ Дюран Э.Ю., Паттерсон Н., Райх Д., Слаткин М. (август 2011 г.). «Тестирование древней примеси между близкородственными популяциями» . Молекулярная биология и эволюция . 28 (8): 2239–52. дои : 10.1093/molbev/msr048 . ПМК 3144383 . ПМИД 21325092 .
- ^ Питер Б.М. (апрель 2016 г.). «Примесь, структура популяции и F-статистика» . Генетика . 202 (4): 1485–501. дои : 10.1534/genetics.115.183913 . ПМЦ 4905545 . ПМИД 26857625 .
- ^ Jump up to: а б Райх Д., Тангарадж К., Паттерсон Н., Прайс А.Л., Сингх Л. (сентябрь 2009 г.). «Реконструкция истории населения Индии» . Природа . 461 (7263): 489–94. Бибкод : 2009Natur.461..489R . дои : 10.1038/nature08365 . ПМЦ 2842210 . ПМИД 19779445 .
- ^ Мартин С.Х., Дэйви Дж.В., компакт-диск Джиггинса (январь 2015 г.). «Оценка использования статистики ABBA-BABA для обнаружения интрогрессивных локусов» . Молекулярная биология и эволюция . 32 (1): 244–57. дои : 10.1093/molbev/msu269 . ПМЦ 4271521 . ПМИД 25246699 .
- ^ Пиз Дж.Б., Хан М.В. (июль 2015 г.). «Обнаружение и поляризация интрогрессии в пятитаксонной филогении» . Систематическая биология . 64 (4): 651–62. дои : 10.1093/sysbio/syv023 . ПМИД 25888025 .
- ^ Итон Д.А., Ри Р.Х. (сентябрь 2013 г.). «Вывод о филогении и интрогрессии с использованием данных RADseq: пример цветковых растений (Pedularis: Orobanchaceae)» . Систематическая биология . 62 (5): 689–706. дои : 10.1093/sysbio/syt032 . ПМЦ 3739883 . ПМИД 23652346 .
- ^ Пикрелл Дж. К., Причард Дж. К. (2012). Тан Х (ред.). «Вывод о разделении и смесях популяций на основе данных о частоте аллелей по всему геному» . ПЛОС Генетика . 8 (11): e1002967. arXiv : 1206.2332 . Бибкод : 2012arXiv1206.2332P . дои : 10.1371/journal.pgen.1002967 . ПМК 3499260 . ПМИД 23166502 .
- ^ Jump up to: а б Паттерсон Н., Мурджани П., Луо Й., Маллик С., Роланд Н., Жан Й. и др. (ноябрь 2012 г.). «Древняя примесь в истории человечества» . Генетика . 192 (3): 1065–93. дои : 10.1534/genetics.112.145037 . ПМК 3522152 . ПМИД 22960212 .
- ^ Липсон М., Лох П.Р., Левин А., Райх Д., Паттерсон Н., Бергер Б. (август 2013 г.). «Эффективный вывод параметров примесей и источников потока генов на основе моментов» . Молекулярная биология и эволюция . 30 (8): 1788–802. дои : 10.1093/molbev/mst099 . ПМК 3708505 . ПМИД 23709261 .
- ^ Ю Ю, Барнетт Р.М., Накле Л. (сентябрь 2013 г.). «Экономный вывод о гибридизации при неполной сортировке линий» . Систематическая биология . 62 (5): 738–51. дои : 10.1093/sysbio/syt037 . ПМЦ 3739885 . ПМИД 23736104 .
- ^ Вэнь Д., Ю Ю, Накле Л. (май 2016 г.). Эдвардс С. (ред.). «Байесовский вывод сетчатой филогении в условиях слияния многовидовой сети» . ПЛОС Генетика . 12 (5): e1006006. дои : 10.1371/journal.pgen.1006006 . ПМЦ 4856265 . ПМИД 27144273 .
- ^ Мурджани П., Паттерсон Н., Хиршхорн Дж.Н., Кейнан А., Хао Л., Ацмон Г. и др. (апрель 2011 г.). Маквин Дж. (ред.). «История потока африканских генов в южных европейцев, левантийцев и евреев» . ПЛОС Генетика . 7 (4): e1001373. дои : 10.1371/journal.pgen.1001373 . ПМК 3080861 . ПМИД 21533020 .
- ^ Мурджани П., Санкарараман С., Фу К., Пшеворски М., Паттерсон Н., Райх Д. (май 2016 г.). «Генетический метод датирования древних геномов обеспечивает прямую оценку интервала поколений людей за последние 45 000 лет» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (20): 5652–7. Бибкод : 2016PNAS..113.5652M . дои : 10.1073/pnas.1514696113 . ПМЦ 4878468 . ПМИД 27140627 .
- ^ Лох П.Р., Липсон М., Паттерсон Н., Мурджани П., Пикрелл Дж.К., Райх Д., Бергер Б. (апрель 2013 г.). «Вывод об истории смешения человеческих популяций с использованием неравновесия по сцеплению» . Генетика . 193 (4): 1233–54. дои : 10.1534/genetics.112.147330 . ПМК 3606100 . ПМИД 23410830 .
- ^ Санкарараман С., Паттерсон Н., Ли Х., Паабо С., Райх Д. (2012). Эйки Дж.М. (ред.). «Дата скрещивания неандертальцев и современных людей» . ПЛОС Генетика . 8 (10): e1002947. arXiv : 1208.2238 . Бибкод : 2012arXiv1208.2238S . дои : 10.1371/journal.pgen.1002947 . ПМЦ 3464203 . ПМИД 23055938 .
- ^ Пиньо С., Эй Джей (2010). «Дивергенция с потоком генов: модели и данные» . Ежегодный обзор экологии, эволюции и систематики . 41 (1): 215–230. doi : 10.1146/annurev-ecolsys-102209-144644 . ISSN 1543-592X . S2CID 45813707 .
- ^ Экскофье Л., Дюпанлуп И., Уэрта-Санчес Э., Соуза В.К., Фолль М. (октябрь 2013 г.). Эйки Дж.М. (ред.). «Надежные демографические выводы на основе геномных данных и данных SNP» . ПЛОС Генетика . 9 (10): е1003905. дои : 10.1371/journal.pgen.1003905 . ПМК 3812088 . ПМИД 24204310 .
- ^ Гутенкунст Р.Н., Эрнандес Р.Д., Уильямсон С.Х., Бустаманте CD (октябрь 2009 г.). Маквин Дж. (ред.). «Вывод совместной демографической истории нескольких популяций на основе многомерных данных о частоте SNP» . ПЛОС Генетика . 5 (10): е1000695. arXiv : 0909.0925 . Бибкод : 2009arXiv0909.0925G . дои : 10.1371/journal.pgen.1000695 . ПМК 2760211 . ПМИД 19851460 .
- ^ Бомонт, Массачусетс (2010). «Приблизительные байесовские вычисления в эволюции и экологии». Ежегодный обзор экологии, эволюции и систематики . 41 (1): 379–406. doi : 10.1146/annurev-ecolsys-102209-144621 .
- ^ Тойнерт С., Слаткин М. (февраль 2017 г.). «Отличение недавней примеси от структуры предкового населения» . Геномная биология и эволюция . 9 (3): 427–437. дои : 10.1093/gbe/evx018 . ПМЦ 5381645 . ПМИД 28186554 .
- ^ Jump up to: а б с Шумер М., Розенталь Г.Г., Андольфатто П. (июнь 2014 г.). «Насколько распространено гомоплоидное гибридное видообразование?» . Эволюция; Международный журнал органической эволюции . 68 (6): 1553–60. дои : 10.1111/evo.12399 . ПМИД 24620775 . S2CID 22702297 .
- ^ Jump up to: а б Ньето Фелинер Дж., Альварес И., Фуэртес-Агилар Дж., Хойертц М., Маркес И., Мохаррек Ф. и др. (июнь 2017 г.). «Неужели гомоплоидное гибридное видообразование настолько редкое? Взгляд эмпирика» . Наследственность . 118 (6): 513–516. дои : 10.1038/hdy.2017.7 . ПМЦ 5436029 . ПМИД 28295029 .
- ^ Jump up to: а б Ризеберг Л.Х., Раймонд О., Розенталь Д.М., Лай З., Ливингстон К., Наказато Т. и др. (август 2003 г.). «Основные экологические изменения в диких подсолнечниках, вызванные гибридизацией» . Наука . 301 (5637): 1211–6. Бибкод : 2003Sci...301.1211R . дои : 10.1126/science.1086949 . ПМИД 12907807 . S2CID 9232157 .
- ^ Грант V (1981). Видообразование растений (2-е изд.). Нью-Йорк: Издательство Колумбийского университета. ISBN 0231051123 . OCLC 7552165 .
- ^ Jump up to: а б с д Шумер М., Сюй С., Пауэлл Д.Л., Дурвасула А., Сков Л., Холланд С. и др. (май 2018 г.). «Естественный отбор взаимодействует с рекомбинацией, формируя эволюцию гибридных геномов» . Наука . 360 (6389): 656–660. Бибкод : 2018Sci...360..656S . дои : 10.1126/science.aar3684 . ПМК 6069607 . ПМИД 29674434 .
- ^ Jump up to: а б с д и ж Рунемарк А., Трир К.Н., Ерухманов Ф., Хермансен Дж.С., Матчинер М., Равинет М. и др. (март 2018 г.). «Вариации и ограничения в формировании гибридного генома». Экология и эволюция природы . 2 (3): 549–556. Бибкод : 2018NatEE...2..549R . дои : 10.1038/s41559-017-0437-7 . ПМИД 29335572 . S2CID 3375880 .
- ^ Jump up to: а б с д Бюеркле, Калифорния, Ризеберг, Л.Х. (февраль 2008 г.). «Скорость стабилизации генома гомоплоидных гибридных видов» . Эволюция; Международный журнал органической эволюции . 62 (2): 266–75. дои : 10.1111/j.1558-5646.2007.00267.x . ПМК 2442919 . ПМИД 18039323 .
- ^ Унгерер MC, Бэрд С.Дж., Пан Дж., Ризеберг Л.Х. (сентябрь 1998 г.). «Быстрое гибридное видообразование диких подсолнечников» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (20): 11757–62. Бибкод : 1998PNAS...9511757U . дои : 10.1073/pnas.95.20.11757 . ПМК 21713 . ПМИД 9751738 .
- ^ Jump up to: а б с Лай З., Наказато Т., Салмасо М., Берк Дж.М., Тан С., Кнапп С.Дж., Ризеберг Л.Х. (сентябрь 2005 г.). «Обширная хромосомная перестройка и эволюция барьеров стерильности у гибридных видов подсолнечника» . Генетика . 171 (1): 291–303. дои : 10.1534/genetics.105.042242 . ПМЦ 1456521 . ПМИД 16183908 .
- ^ Jump up to: а б Элгвин Т.О., Трир К.Н., Торресен О.К., Хаген И.Дж., Лиен С., Недербрагт А.Дж. и др. (июнь 2017 г.). «Геномный мозаицизм гибридного видообразования» . Достижения науки . 3 (6): e1602996. Бибкод : 2017SciA....3E2996E . дои : 10.1126/sciadv.1602996 . ПМК 5470830 . ПМИД 28630911 .
- ^ Jump up to: а б Отто С.П., Уиттон Дж (2000). «Заболеваемость и эволюция полиплоидов». Ежегодный обзор генетики . 34 (1): 401–437. дои : 10.1146/annurev.genet.34.1.401 . ПМИД 11092833 .
- ^ Эбботт Р.Дж., Ризеберг Л.Х. (2012). Гибридный вид . John Wiley & Sons, Ltd. doi : 10.1002/9780470015902.a0001753.pub2 . ISBN 9780470016176 . S2CID 242640180 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ Jump up to: а б Койн Дж. А. (октябрь 1989 г.). «Степень мутаций в гибридах между родственными видами дрозофилы» . Наследственность . 63 (Часть 2) (2): 155–62. дои : 10.1038/hdy.1989.87 . ПМИД 2553645 .
- ^ Чейз М.В., Паун О., Фэй М.Ф. (2010). «Гибридизация и видообразование покрытосеменных: роль в смене опылителей?» . Журнал биологии . 9 (3): 21. дои : 10.1186/jbiol231 . ISSN 1475-4924 .
- ^ Грант V (март 1949 г.). «Системы опыления как изолирующие механизмы у покрытосеменных» . Эволюция; Международный журнал органической эволюции . 3 (1): 82–97. дои : 10.1111/j.1558-5646.1949.tb00007.x . ПМИД 18115119 .
- ^ Сегрейвс К.А., Томпсон Дж.Н. (август 1999 г.). «Полиплоидия и опыление растений: цветочные особенности и посещения насекомыми диплоида и тетраплоидейхеры Grossulariifolia» . Эволюция; Международный журнал органической эволюции . 53 (4): 1114–1127. дои : 10.1111/j.1558-5646.1999.tb04526.x . ПМИД 28565509 . S2CID 28429201 .
- ^ Мо А.М., Вайблен Г.Д. (декабрь 2012 г.). «Репродуктивная изоляция, опосредованная опылителями, среди раздельнополых видов инжира (Ficus, Moraceae)» . Эволюция; Международный журнал органической эволюции . 66 (12): 3710–21. дои : 10.1111/j.1558-5646.2012.01727.x . ПМИД 23206130 . S2CID 26585628 .
- ^ Jump up to: а б Лоу А.Дж., Эбботт Р.Дж. (май 2004 г.). «Репродуктивная изоляция нового гибридного вида Senecio eboracensis Abbott & Lowe (Asteraceae)» . Наследственность . 92 (5): 386–95. дои : 10.1038/sj.hdy.6800432 . ПМИД 15014422 .
- ^ Зельц О.М., Томмен Р., Маан М.Е., Зеехаузен О. (февраль 2014 г.). «Поведенческая изоляция может способствовать образованию гомоплоидных гибридов у цихлид» . Журнал эволюционной биологии . 27 (2): 275–89. дои : 10.1111/jeb.12287 . ПМИД 24372872 .
- ^ Jump up to: а б Шварцбах А.Е., Донован Л.А., Ризеберг Л.Х. (2001). «Трансгрессивное выражение характера у гибридных видов подсолнечника». Американский журнал ботаники . 88 (2): 270–277. дои : 10.2307/2657018 . ISSN 0002-9122 . JSTOR 2657018 . ПМИД 11222249 .
- ^ Мамели Дж., Лопес-Альварадо Дж., Фаррис Э., Сюзанна А., Филигедду Р., Гарсия-Хакас Н. (2014). «Роль родительских и гибридных видов в множественных событиях интрогрессии: свидетельства гомоплоидного гибридного видообразования у Centaurea (Cardueae, Asteraceae): интрогрессия у Centaurea» . Журнал Ботанического общества Линнеевского общества . 175 (3): 453–467. дои : 10.1111/box.12177 .
- ^ Се X, Мишель А.П., Шварц Д., Рулл Дж., Велес С., Форбс А.А. и др. (май 2008 г.). «Радиация и дивергенция в комплексе видов Rhagoletis pomonella: выводы на основе данных о последовательности ДНК» . Журнал эволюционной биологии . 21 (3): 900–13. дои : 10.1111/j.1420-9101.2008.01507.x . ПМИД 18312319 .
- ^ Федер Дж.Л., Се Х., Рулл Дж., Велес С., Форбс А., Люнг Б. и др. (май 2005 г.). «Майр, Добжанский и Буш и сложности симпатрического видообразования у Rhagoletis» . Труды Национальной академии наук Соединенных Штатов Америки . 102 Приложение 1 (Приложение 1): 6573–80. Бибкод : 2005PNAS..102.6573F . дои : 10.1073/pnas.0502099102 . ПМЦ 1131876 . ПМИД 15851672 .
- ^ Шумер М., Пауэлл Д.Л., Дельклос П.Дж., Сквайр М., Кюи Р., Андольфатто П., Розенталь Г.Г. (октябрь 2017 г.). «Ассортативное спаривание и стойкая репродуктивная изоляция у гибридов» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (41): 10936–10941. Бибкод : 2017PNAS..11410936S . дои : 10.1073/pnas.1711238114 . ПМК 5642718 . ПМИД 28973863 .
- ^ Ризеберг Л.Х., Линдер Ч.Р., Зайлер Г.Дж. (ноябрь 1995 г.). «Хромосомные и генные барьеры на пути интрогрессии у Helianthus» . Генетика . 141 (3): 1163–71. дои : 10.1093/генетика/141.3.1163 . ПМК 1206838 . ПМИД 8582621 .
- ^ Jump up to: а б Comeault AA, Matute DR (сентябрь 2018 г.). «Генетическая дивергенция и количество гибридизующихся видов влияют на путь к гомоплоидному гибридному видообразованию» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (39): 9761–9766. Бибкод : 2018PNAS..115.9761C . дои : 10.1073/pnas.1809685115 . ПМК 6166845 . ПМИД 30209213 .
- ^ Бланкарт А, Банк C (сентябрь 2018 г.). Чжан Дж (ред.). «В поисках зоны Златовласки гибридного видообразования» . ПЛОС Генетика . 14 (9): e1007613. дои : 10.1371/journal.pgen.1007613 . ПМК 6145587 . ПМИД 30192761 .
- ^ Шумер М., Кюи Р., Розенталь Г.Г., Андольфатто П. (март 2015 г.). Payseur BA (ред.). «Репродуктивная изоляция гибридных популяций, вызванная генетической несовместимостью» . ПЛОС Генетика . 11 (3): e1005041. дои : 10.1371/journal.pgen.1005041 . ПМК 4359097 . ПМИД 25768654 .
- ^ Верекен, штат Нью-Джерси, Коццолино С., Шистль Ф.П. (апрель 2010 г.). «Новинка гибридного цветочного аромата приводит к смене опылителей у сексуально обманчивых орхидей» . Эволюционная биология BMC . 10 (1): 103. Бибкод : 2010BMCEE..10..103V . дои : 10.1186/1471-2148-10-103 . ПМЦ 2875231 . ПМИД 20409296 .
- ^ Гаэта RT, Крис Пирес Дж (апрель 2010 г.). «Гомеологическая рекомбинация у аллополиплоидов: полиплоидный храповик» . Новый фитолог . 186 (1): 18–28. дои : 10.1111/j.1469-8137.2009.03089.x . ПМИД 20002315 .
- ^ Jump up to: а б Хвала Дж.А., Фрайер М.Э., Payseur BA (май 2018 г.). «Признаки гибридизации и видообразования в геномных моделях происхождения» . Эволюция; Международный журнал органической эволюции . 72 (8): 1540–1552. дои : 10.1111/evo.13509 . ПМК 6261709 . ПМИД 29806154 .
- ^ Ризеберг Л.Х., Синерво Б., Линдер Ч.Р., Унгерер М.С., Ариас Д.М. (май 1996 г.). «Роль взаимодействия генов в гибридном видообразовании: данные древних и экспериментальных гибридов». Наука . 272 (5262): 741–5. Бибкод : 1996Sci...272..741R . дои : 10.1126/science.272.5262.741 . ПМИД 8662570 . S2CID 39005242 .
- ^ Стукенброк Э.Х., Кристиансен Ф.Б., Хансен Т.Т., Дютейл Ю.Ю., Шируп М.Х. (июль 2012 г.). «Слияние двух расходящихся грибковых особей привело к недавнему появлению уникального широко распространенного вида патогена» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (27): 10954–9. Бибкод : 2012PNAS..10910954S . дои : 10.1073/pnas.1201403109 . ПМК 3390827 . ПМИД 22711811 .
- ^ Jump up to: а б Шумер М., Брандвейн Ю. (июнь 2016 г.). «Определение эпистатического отбора в смешанных популяциях». Молекулярная экология . 25 (11): 2577–91. Бибкод : 2016MolEc..25.2577S . дои : 10.1111/mec.13641 . ПМИД 27061282 . S2CID 10566380 .
- ^ Санкарараман С., Маллик С., Даннеманн М., Прюфер К., Келсо Дж., Паабо С. и др. (март 2014 г.). «Геномный ландшафт предков неандертальцев у современных людей» . Природа . 507 (7492): 354–7. Бибкод : 2014Natur.507..354S . дои : 10.1038/nature12961 . ПМК 4072735 . ПМИД 24476815 .
- ^ Ерухманов Ф., Бейли Р.И., Элгвин Т.О., Хермансен Дж.С., Рунемарк А.Р., Трир К.Н., Сэтре Г.П. (2017). «Разрешение конфликта между родительскими геномами у гибридного вида» . биоRxiv . дои : 10.1101/102970 . hdl : 20.500.11820/e14c46c6-ef76-41a4-a76d-88604ba5b30b .
- ^ Охта Т (1971). «Ассоциативное доминирование, вызванное связанными вредными мутациями» . Генетические исследования . 18 (3): 277–286. дои : 10.1017/s0016672300012684 . ISSN 0016-6723 . ПМИД 5158298 .
- ^ Чжао Л., Чарльзуорт Б. (июль 2016 г.). «Разрешение конфликта между ассоциативным доминированием и выбором фона» . Генетика . 203 (3): 1315–34. дои : 10.1534/genetics.116.188912 . ПМЦ 4937488 . ПМИД 27182952 .
- ^ Фариа Р., Йоханнессон К. , Батлин Р.К., Вестрам А.М. (март 2019 г.). «Развивающиеся инверсии» . Тенденции в экологии и эволюции . 34 (3): 239–248. дои : 10.1016/j.tree.2018.12.005 . ПМИД 30691998 .
- ^ Jump up to: а б Бартон, Нью-Хэмпшир (декабрь 2018 г.). «Последствия интрогрессионного события» . Молекулярная экология . 27 (24): 4973–4975. Бибкод : 2018MolEc..27.4973B . дои : 10.1111/mec.14950 . ПМИД 30599087 .
- ^ Jump up to: а б Мартин С.Х., Дэйви Дж.В., Салазар С., Джиггинс CD (февраль 2019 г.). Мойл Л. (ред.). «Вариации скорости рекомбинации формируют барьеры для интрогрессии в геномах бабочек» . ПЛОС Биология . 17 (2): e2006288. дои : 10.1371/journal.pbio.2006288 . ПМК 6366726 . ПМИД 30730876 .
- ^ Брандвейн Ю., Кенни А.М., Флагель Л., Куп Г., Свейгарт А.Л. (июнь 2014 г.). Компакт-диск Джиггинса (ред.). «Видообразование и интрогрессия между Mimulus nasutus и Mimulus Guttatus» . ПЛОС Генетика . 10 (6): e1004410. дои : 10.1371/journal.pgen.1004410 . ПМК 4072524 . ПМИД 24967630 .
- ^ Янушек В., Манклингер П., Ван Л., Титер К.К., Такер П.К. (май 2015 г.). «Функциональная организация генома может определять границы вида домовой мыши» . Молекулярная биология и эволюция . 32 (5): 1208–20. дои : 10.1093/molbev/msv011 . ПМК 4408407 . ПМИД 25631927 .
- ^ Шумер М., Кюи Р., Пауэлл Д.Л., Дреснер Р., Розенталь Г.Г., Андольфатто П. (июнь 2014 г.). «Картирование с высоким разрешением выявляет сотни генетических несовместимостей при гибридизации видов рыб» . электронная жизнь . 3 . doi : 10.7554/eLife.02535 . ПМК 4080447 . ПМИД 24898754 .
- ^ Jump up to: а б с Лю С., Луо Дж., Чай Дж., Рен Л., Чжоу Ю., Хуан Ф. и др. (февраль 2016 г.). «Геномная несовместимость у диплоидного и тетраплоидного потомства скрещивания золотой рыбки и карпа» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (5): 1327–32. Бибкод : 2016PNAS..113.1327L . дои : 10.1073/pnas.1512955113 . ПМЦ 4747765 . ПМИД 26768847 .
- ^ Трир К.Н., Хермансен Дж.С., Сатре Г.П., Бейли Р.И. (январь 2014 г.). Компакт-диск Джиггинса (ред.). «Доказательства существования мито-ядерных и сцепленных с полом репродуктивных барьеров между гибридным итальянским воробьем и его родительскими видами» . ПЛОС Генетика . 10 (1): e1004075. дои : 10.1371/journal.pgen.1004075 . ПМЦ 3886922 . ПМИД 24415954 .
- ^ Джордано Л., Силло Ф., Гарбелотто М., Гонтье П. (январь 2018 г.). «Митоядерные взаимодействия могут способствовать приспособленности грибных гибридов» . Научные отчеты . 8 (1): 1706. Бибкод : 2018НатСР...8.1706Г . doi : 10.1038/s41598-018-19922-w . ПМК 5786003 . ПМИД 29374209 .
- ^ Дело А.Л., Финсет Ф.Р., Барр С.М., Фишман Л. (сентябрь 2016 г.). «Эгоистичная эволюция цитоядерной гибридной несовместимости у Mimulus» . Слушания. Биологические науки . 283 (1838): 20161493. doi : 10.1098/rspb.2016.1493 . ПМК 5031664 . ПМИД 27629037 .
- ^ Дэвид В.М., Митчелл Д.Л., Уолтер Р.Б. (июль 2004 г.). «Репарация ДНК гибридных рыб рода Xiphophorus». Сравнительная биохимия и физиология. Токсикология и фармакология . 138 (3): 301–9. дои : 10.1016/j.cca.2004.07.006 . ПМИД 15533788 .
- ^ Авила В., Чаварриас Д., Санчес Э., Манрике А., Лопес-Фанжул С., Гарсиа-Дорадо А. (май 2006 г.). «Увеличение частоты спонтанных мутаций в длительном эксперименте с Drosophila melanogaster» . Генетика . 173 (1): 267–77. дои : 10.1534/генетика.106.056200 . ПМЦ 1461422 . ПМИД 16547099 .
- ^ Башир Т., Сайлер С., Гербер Ф., Логанатан Н., Бхупалан Х., Эйхенбергер С. и др. (май 2014 г.). «Гибридизация изменяет частоту спонтанных мутаций у арабидопсиса в зависимости от родителя» . Физиология растений . 165 (1): 424–37. дои : 10.1104/стр.114.238451 . ПМК 4012600 . ПМИД 24664208 .
- ^ Денненмозер С., Седлазек Ф.Дж., Ивашкевич Е., Ли XY, Альтмюллер Дж., Нольте А.В. (сентябрь 2017 г.). «Увеличение числа копий мобильных элементов и генов, кодирующих белки, у инвазивных рыб гибридного происхождения» . Молекулярная экология . 26 (18): 4712–4724. Бибкод : 2017MolEc..26.4712D . дои : 10.1111/mec.14134 . ПМЦ 5638112 . ПМИД 28390096 .
- ^ Дион-Коте А.М., Рено С., Нормандо Э., Бернатчес Л. (май 2014 г.). «РНК-секвенирование выявляет транскриптомный шок, включающий реактивацию мобильных элементов у гибридов молодых видов озерного сига» . Молекулярная биология и эволюция . 31 (5): 1188–99. дои : 10.1093/molbev/msu069 . ПМИД 24505119 .
- ^ Сенерчия Н., Фельбер Ф., Парисод К. (апрель 2015 г.). «Реорганизация генома гибридов F1 раскрывает роль ретротранспозонов в репродуктивной изоляции» . Слушания. Биологические науки . 282 (1804): 20142874. doi : 10.1098/rspb.2014.2874 . ПМЦ 4375867 . ПМИД 25716787 .
- ^ Остберг К.О., Хаузер Л., Притчард В.Л., Гарза Дж.К., Наиш К.А. (август 2013 г.). «Хромосомные перестройки, подавление рекомбинации и ограниченное искажение сегрегации у гибридов йеллоустонской головорезной форели (Oncorhynchus clarkii bouvieri) и радужной форели (O. mykiss)» . БМК Геномика . 14 (1): 570. дои : 10.1186/1471-2164-14-570 . ПМЦ 3765842 . ПМИД 23968234 .
- ^ Хираи Х., Хираи Ю., Моримото М., Канеко А., Каманака Ю., Кога А. (апрель 2017 г.). «Гибриды ночных обезьян демонстрируют геномные и кариотипические изменения De Novo: первый такой случай у приматов» . Геномная биология и эволюция . 9 (4): 945–955. дои : 10.1093/gbe/evx058 . ПМЦ 5388293 . ПМИД 28369492 .
- ^ Баркан А., Мартиенссен Р.А. (апрель 1991 г.). «Инактивация кукурузного транспозона Mu подавляет мутантный фенотип путем активации промотора внешнего считывания вблизи конца Mu1» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (8): 3502–6. Бибкод : 1991PNAS...88.3502B . дои : 10.1073/pnas.88.8.3502 . ПМК 51476 . ПМИД 1849660 .
- ^ Райзада М.Н., Бенито М.И., Уолбот В. (2008). «Концевой инвертированный повтор транспозона MuDR содержит сложный растительный промотор, управляющий различными соматическими и зародышевыми программами: образец экспрессии промотора транспозона» . Заводской журнал . 25 (1): 79–91. дои : 10.1111/j.1365-313X.2001.00939.x . S2CID 26084899 .
- ^ Лим К.Ю., Матьясек Р., Коварик А., Лейтч А.Р. (2004). «Эволюция генома аллотетраплоида Никотианы» . Биологический журнал Линнеевского общества . 82 (4): 599–606. дои : 10.1111/j.1095-8312.2004.00344.x .
- ^ Баак Э.Дж., Уитни К.Д., Ризеберг Л.Х. (август 2005 г.). «Гибридизация и эволюция размера генома: время и величина увеличения содержания ядерной ДНК у гомоплоидных гибридных видов Helianthus» . Новый фитолог . 167 (2): 623–30. дои : 10.1111/j.1469-8137.2005.01433.x . ПМЦ 2442926 . ПМИД 15998412 .
- ^ Лейтч И.Дж., Хэнсон Л., Лим К.Ю., Коварик А., Чейз М.В., Кларксон Дж.Дж., Лейтч А.Р. (апрель 2008 г.). «Взлеты и падения размера генома полиплоидных видов Nicotiana (Solanaceae)» . Анналы ботаники . 101 (6): 805–14. дои : 10.1093/aob/mcm326 . ПМК 2710205 . ПМИД 18222910 .
- ^ Вулф К.Х. (май 2001 г.). «Вчерашние полиплоиды и тайна диплоидизации» . Обзоры природы. Генетика . 2 (5): 333–41. дои : 10.1038/35072009 . ПМИД 11331899 .
- ^ Jump up to: а б с Фрилинг М., Скэнлон М.Дж., Фаулер Дж.Э. (декабрь 2015 г.). «Фракционирование и субфункционализация после дупликации генома: механизмы, которые управляют содержанием генов и их последствия». Текущее мнение в области генетики и развития . 35 : 110–8. дои : 10.1016/j.где.2015.11.002 . ПМИД 26657818 .
- ^ Jump up to: а б Санкофф Д., Чжэн С., Чжу Ц (май 2010 г.). «Коллапс набора генов после полной дупликации генома» . БМК Геномика . 11 (1): 313. дои : 10.1186/1471-2164-11-313 . ПМЦ 2896955 . ПМИД 20482863 .
- ^ Jump up to: а б Эджер П.П., Поортен Т.Дж., ВанБюрен Р., Хардиган М.А., Колле М., Маккейн М.Р. и др. (март 2019 г.). «Происхождение и эволюция октоплоидного генома клубники» . Природная генетика . 51 (3): 541–547. дои : 10.1038/s41588-019-0356-4 . ПМЦ 6882729 . ПМИД 30804557 .
- ^ Jump up to: а б Эджер П.П., Смит Р., Маккейн М.Р., Кули А.М., Вальехо-Марин М., Юань Ю. и др. (сентябрь 2017 г.). «Субгеномное доминирование в межвидовом гибриде, синтетическом аллополиплоиде и 140-летнем естественно созданном неоаллополиплоидном обезьяньем цветке» . Растительная клетка . 29 (9): 2150–2167. дои : 10.1105/tpc.17.00010 . ПМК 5635986 . ПМИД 28814644 .
- ^ Сюй С, Бай Ю, Линь Х, Чжао Н, Ху Л, Гонг З и др. (май 2014 г.). «Общегеномное нарушение экспрессии генов у аллополиплоидов, но не у гибридов подвидов риса» . Молекулярная биология и эволюция . 31 (5): 1066–76. дои : 10.1093/molbev/msu085 . ПМЦ 3995341 . ПМИД 24577842 .
- ^ Ренаут С., Нолте А.В., Бернатчес Л. (апрель 2009 г.). «Расхождение в экспрессии генов и гибридная неправильная экспрессия между парами видов озерного сига (Coregonus spp. Salmonidae)» . Молекулярная биология и эволюция . 26 (4): 925–36. дои : 10.1093/molbev/msp017 . ПМИД 19174479 .
- ^ Баггс Р.Дж., Чжан Л., Майлз Н., Тейт Дж.А., Гао Л., Вэй В. и др. (апрель 2011 г.). «Транскриптомный шок порождает эволюционную новизну во вновь сформированном природном аллополиплоидном растении» . Современная биология . 21 (7): 551–6. Бибкод : 2011CBio...21..551B . дои : 10.1016/j.cub.2011.02.016 . ПМИД 21419627 . S2CID 18417406 .
- ^ Ха М., Лу Дж., Тиан Л., Рамачандран В., Касшау К.Д., Чепмен Э.Дж. и др. (октябрь 2009 г.). «Маленькие РНК служат генетическим буфером против геномного шока у межвидовых гибридов и аллополиплоидов Arabidopsis» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (42): 17835–40. Бибкод : 2009PNAS..10617835H . дои : 10.1073/pnas.0907003106 . ПМЦ 2757398 . ПМИД 19805056 .
- ^ Маллет Дж. (май 2005 г.). «Гибридизация как вторжение в геном». Тенденции в экологии и эволюции . 20 (5): 229–37. дои : 10.1016/j.tree.2005.02.010 . ПМИД 16701374 .
- ^ Чарльзуорт Д. (апрель 2016 г.). «Половые хромосомы растений». Ежегодный обзор биологии растений . 67 (1): 397–420. doi : 10.1146/annurev-arplant-043015-111911 . ПМИД 26653795 .
- ^ Ризеберг Л.Х. (2001). «Хромосомные перестройки и видообразование». Тенденции в экологии и эволюции . 16 (7): 351–358. дои : 10.1016/s0169-5347(01)02187-5 . ISSN 0169-5347 . ПМИД 11403867 .
- ^ Левин Д.А. (ноябрь 2012 г.). «Долгое ожидание гибридной стерильности цветковых растений» . Новый фитолог . 196 (3): 666–70. дои : 10.1111/j.1469-8137.2012.04309.x . ПМИД 22966819 .
- ^ Холдейн Дж. Б. (1922). «Соотношение полов и однополое бесплодие у гибридных животных» . Журнал генетики . 12 (2): 101–109. дои : 10.1007/BF02983075 . ISSN 0022-1333 . S2CID 32459333 .
- ^ Турелли М., Орр Х.А. (май 1995 г.). «Теория доминирования правления Холдейна» . Генетика . 140 (1): 389–402. дои : 10.1093/генетика/140.1.389 . ПМК 1206564 . ПМИД 7635302 .
- ^ Рунемарк А., Ерухманов Ф., Нава-Боланьос А., Хермансен Дж.С., Мейер Дж.И. (октябрь 2018 г.). «Гибридизация, половая геномная архитектура и локальная адаптация» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 373 (1757): 20170419. doi : 10.1098/rstb.2017.0419 . ПМК 6125728 . ПМИД 30150218 .
- ^ Payseur BA, Rieseberg LH (июнь 2016 г.). «Геномный взгляд на гибридизацию и видообразование» . Молекулярная экология . 25 (11): 2337–60. Бибкод : 2016MolEc..25.2337P . дои : 10.1111/mec.13557 . ПМЦ 4915564 . ПМИД 26836441 .
- ^ Линч М. (1998). Генетика и анализ количественных признаков . Уолш, Брюс, 1957–. Сандерленд, Массачусетс: Синауэр. ISBN 0878934812 . ОСЛК 37030646 .
- ^ Масли Дж. П., Прегрейвс, округ Колумбия (сентябрь 2007 г.). Бартон, Нью-Хэмпшир (ред.). «Полногеномное исследование двух правил видообразования у дрозофилы» . ПЛОС Биология . 5 (9): е243. doi : 10.1371/journal.pbio.0050243 . ЧВК 1971125 . ПМИД 17850182 .
- ^ Манк Дж. Э., Хоскен DJ, Веделл Н. (октябрь 2014 г.). «Конфликт половых хромосом: причина, следствие и сложность» . Перспективы Колд-Спринг-Харбор в биологии . 6 (12): а017715. doi : 10.1101/cshperspect.a017715 . ПМЦ 4292157 . ПМИД 25280765 .
- ^ Брис Р., Ванден Брук А., Мергей Дж., Жакемин Х. (май 2014 г.). «Вклад вариаций системы спаривания в репродуктивную изоляцию у двух близкородственных видов Centaurium (Gentianaceae) с генерализованной морфологией цветков» . Эволюция; Международный журнал органической эволюции . 68 (5): 1281–93. дои : 10.1111/evo.12345 . ПМИД 24372301 .
- ^ Видмер А., Лексер С., Коццолино С. (январь 2009 г.). «Эволюция репродуктивной изоляции у растений» . Наследственность . 102 (1): 31–8. дои : 10.1038/hdy.2008.69 . ПМИД 18648386 .
- ^ Jump up to: а б Шардл CL, Крэйвен К.Д. (ноябрь 2003 г.). «Межвидовая гибридизация растительных грибов и оомицетов: обзор» . Молекулярная экология . 12 (11): 2861–73. Бибкод : 2003MolEc..12.2861S . дои : 10.1046/j.1365-294x.2003.01965.x . ПМИД 14629368 . S2CID 25879264 .
- ^ Левин Д.А. (1975). «Исключение цитотипов меньшинств в местных популяциях растений». Таксон . 24 (1): 35–43. дои : 10.2307/1218997 . JSTOR 1218997 .
- ^ Маккарти Э.М., Асмуссен М.А., Андерсон В.В. (1995). «Теоретическая оценка рекомбинационного видообразования» . Наследственность . 74 (5): 502–509. дои : 10.1038/hdy.1995.71 . ISSN 0018-067X .
- ^ Чарльтон Н.Д., Крэйвен К.Д., Афхами М.Э., Холл Б.А., Гимире С.Р., Янг К.А. (октябрь 2014 г.). «Межвидовая гибридизация и вариации биологически активных алкалоидов увеличивают разнообразие эндофитных видов Epichloë Bromus laevipes» . ФЭМС Микробиология Экология . 90 (1): 276–89. Бибкод : 2014FEMME..90..276C . дои : 10.1111/1574-6941.12393 . ПМИД 25065688 .
- ^ Янко К., Пачес Дж., Уилкинсон-Херботс Х., Коста Р.Дж., Рослейн Дж., Дрозд П. и др. (январь 2018 г.). «Гибридная асексуальность как первичный постзиготический барьер между возникающими видами: о взаимосвязи асексуальности, гибридизации и видообразования» . Молекулярная экология . 27 (1): 248–263. Бибкод : 2018MolEc..27..248J . дои : 10.1111/mec.14377 . ПМК 6849617 . ПМИД 28987005 .
- ^ Нивз В.Б., Бауманн П. (март 2011 г.). «Однополое размножение среди позвоночных». Тенденции в генетике . 27 (3): 81–8. дои : 10.1016/j.tig.2010.12.002 . ПМИД 21334090 .
- ^ Брохманн С., Бристинг А.К., Алсос И.Г., Борген Л., Грундт Х.Х., Шин А.К., Эльф Р. (2004). «Полиплоидия арктических растений» . Биологический журнал Линнеевского общества . 82 (4): 521–536. дои : 10.1111/j.1095-8312.2004.00337.x .
- ^ Норрис Л.К., Мейн Б.Дж., Ли Ю., Кольер Т.К., Фофана А., Корнел А.Дж., Ланзаро Г.К. (январь 2015 г.). «Адаптивная интрогрессия у африканского малярийного комара совпала с более широким использованием обработанных инсектицидами надкроватных сеток» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (3): 815–20. Бибкод : 2015PNAS..112..815N . дои : 10.1073/pnas.1418892112 . ПМЦ 4311837 . ПМИД 25561525 .
- ^ Jump up to: а б с Маркес Д.А., Мейер Дж.И., Зеехаузен О. (июнь 2019 г.). «Комбинаторный взгляд на видообразование и адаптивное излучение» . Тенденции в экологии и эволюции . 34 (6): 531–544. дои : 10.1016/j.tree.2019.02.008 . ПМИД 30885412 . S2CID 83462549 .
- ^ Махешвари С., Барбаш Д.А. (2011). «Генетика гибридной несовместимости». Ежегодный обзор генетики . 45 (1): 331–55. doi : 10.1146/annurev-genet-110410-132514 . ПМИД 21910629 .
- ^ Баггс Р.Дж., Дуст А.Н., Тейт Дж.А., Ко Дж., Солтис К., Фелтус Ф.А. и др. (июль 2009 г.). «Потеря генов и молчание у Tragopogon miscellus (Asteraceae): сравнение природных и синтетических аллотетраплоидов» . Наследственность . 103 (1): 73–81. дои : 10.1038/hdy.2009.24 . ПМИД 19277058 .
- ^ Джиггинс К.Д., Салазар С., Линарес М., Маварес Дж. (сентябрь 2008 г.). «Обзор. Видообразование по гибридному признаку и бабочки Heliconius» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 363 (1506): 3047–54. дои : 10.1098/rstb.2008.0065 . ПМК 2607310 . ПМИД 18579480 .
- ^ Фонтейн М.К., Пиз Дж.Б., Стил А., Уотерхаус Р.М., Нифси Д.Е., Шарахов И.В. и др. (январь 2015 г.). «Геномика комаров. Обширная интрогрессия в комплексе видов переносчиков малярии, выявленная с помощью филогеномики» . Наука . 347 (6217): 1258524. doi : 10.1126/science.1258524 . ПМЦ 4380269 . ПМИД 25431491 .
- ^ Джей П., Уибли А., Фрезал Л., Родригес де Кара Ма, Ноуэлл Р.В., Маллет Дж. и др. (июнь 2018 г.). «Эволюция супергенов, вызванная интрогрессией хромосомной инверсии» . Современная биология . 28 (11): 1839–1845.e3. Бибкод : 2018CBio...28E1839J . дои : 10.1016/j.cub.2018.04.072 . ПМИД 29804810 .
- ^ Йеман С. (май 2013 г.). «Геномные перестройки и эволюция кластеров локально адаптивных локусов» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (19): Е1743-51. Бибкод : 2013PNAS..110E1743Y . дои : 10.1073/pnas.1219381110 . ПМК 3651494 . ПМИД 23610436 .
- ^ У ЦИ (2001). «Генический взгляд на процесс видообразования: Генетический взгляд на процесс видообразования». Журнал эволюционной биологии . 14 (6): 851–865. дои : 10.1046/j.1420-9101.2001.00335.x . S2CID 54863357 .
- ^ Харрисон Р.Г., Ларсон Э.Л. (2014). «Гибридизация, интрогрессия и природа видовых границ» . Журнал наследственности . 105 Приложение 1 (S1): 795–809. дои : 10.1093/jhered/esu033 . ПМИД 25149255 .
- ^ Титер К.С., Пейсер Б.А., Харрис Л.В., Бэйкуэлл М.А., Тибодо Л.М., О'Брайен Дж.Э. и др. (январь 2008 г.). «Общегеномные закономерности потока генов в гибридной зоне домашней мыши» . Геномные исследования . 18 (1): 67–76. дои : 10.1101/гр.6757907 . ПМК 2134771 . ПМИД 18025268 .
- ^ Хупер Д.М., Гриффит СК, Прайс ТД (март 2019 г.). «Инверсия половых хромосом усиливает репродуктивную изоляцию в птичьей гибридной зоне» . Молекулярная экология . 28 (6): 1246–1262. Бибкод : 2019MolEc..28.1246H . дои : 10.1111/mec.14874 . ПМИД 30230092 .