Армирование (вид)

| Часть серии о |
| Эволюционная биология |
|---|
 |
Подкрепление — это процесс видообразования , при котором естественный отбор увеличивает репродуктивную изоляцию (в дальнейшем разделяемую на презиготическую изоляцию и постзиготическую изоляцию ) между двумя популяциями видов. Это происходит в результате отбора, направленного против образования гибридных особей низкой приспособленности . Идея была первоначально разработана Альфредом Расселом Уоллесом и иногда называется эффектом Уоллеса . Современное понятие армирования берет свое начало от Феодосия Добжанского . Он представил себе вид, разделенный аллопатрически , где во время вторичного контакта две популяции спариваются, производя гибриды с более низкой приспособленностью. Естественный отбор является результатом неспособности гибрида производить жизнеспособное потомство; таким образом, представители одного вида, которые не спариваются с представителями другого вида, имеют больший репродуктивный успех. Это способствует развитию большей презиготической изоляции (различия в поведении или биологии, которые препятствуют образованию гибридных зигот ). Подкрепление — один из немногих случаев, когда отбор может способствовать усилению презиготической изоляции, напрямую влияя на процесс видообразования. [ 1 ] Этот аспект особенно привлекал биологов-эволюционистов. [ 2 ]
Поддержка подкрепления колебалась с момента его создания, а терминологическая путаница и различия в использовании на протяжении истории привели к множеству значений и осложнениям. выдвигали различные возражения Биологи-эволюционисты относительно вероятности его возникновения. С 1990-х годов данные теории, экспериментов и природы преодолели многие прежние возражения, сделав подкрепление широко принятым. [ 3 ] : 354 хотя его распространенность в природе остается неизвестной. [ 4 ] [ 5 ]
Чтобы понять его действие в природе, было разработано множество моделей, большинство из которых основаны на нескольких аспектах: генетике , популяционных структурах, влиянии отбора и брачном поведении. Эмпирическая поддержка подкрепления существует как в лаборатории, так и в природе. Документированные примеры встречаются у широкого круга организмов: как позвоночных , так и беспозвоночных , грибов и растений. Вторичный контакт первоначально разделенных зачаточных видов (начальная стадия видообразования) увеличивается из-за деятельности человека, такой как внедрение инвазивных видов или изменение естественной среды обитания . [ 6 ] Это имеет последствия для показателей биоразнообразия и может стать более актуальным в будущем. [ 6 ]
История
[ редактировать ]У подкрепления сложная история: его популярность среди ученых со временем менялась. [ 7 ] [ 8 ] Джерри Койн и Х. Аллен Орр утверждают, что теория подкрепления прошла три фазы исторического развития: [ 3 ] : 366
- правдоподобие на основе непригодных гибридов
- неправдоподобие, основанное на некоторой приспособленности гибридов
- правдоподобие, основанное на эмпирических исследованиях и биологически сложных и реалистичных моделях

Подкрепление, которое иногда называют эффектом Уоллеса, было первоначально предложено Альфредом Расселом Уоллесом в 1889 году. [ 9 ] : 353 Его гипотеза заметно отличалась от современной концепции тем, что сосредоточивалась на постзиготической изоляции , усиленной групповым отбором . [ 10 ] [ 11 ] [ 3 ] : 353 Феодосий Добжанский был первым, кто дал подробное описание процесса в 1937 году. [ 3 ] : 353 хотя сам термин не был придуман до 1955 года У. Фрэнком Блэром . [ 12 ] В 1930 году Рональд Фишер изложил первое генетическое описание процесса подкрепления в «Генетической теории естественного отбора» , а в 1965 и 1970 годах было проведено первое компьютерное моделирование для проверки его правдоподобия. [ 3 ] : 367 Более поздняя популяционная генетика [ 13 ] и количественные генетические [ 14 ] были проведены исследования, показавшие, что совершенно непригодные гибриды однозначно приводят к увеличению презиготической изоляции. [ 3 ] : 367
Идея Добжанского получила значительную поддержку; он предположил, что это иллюстрирует последний этап видообразования, например, после того, как аллопатрическая популяция вступает во вторичный контакт. [ 3 ] : 353 В 1980-х годах многие биологи-эволюционисты начали сомневаться в правдоподобности этой идеи. [ 3 ] : 353 основано не на эмпирических данных, а в основном на развитии теории, которая считала это маловероятным механизмом репродуктивной изоляции. [ 2 ] В то время возник ряд теоретических возражений, которые рассматриваются в разделе «Аргументы против подкрепления» ниже.
К началу 1990-х годов популярность подкрепления среди биологов-эволюционистов возродилась; Прежде всего, это связано с внезапным увеличением количества данных — эмпирических данных, полученных в результате исследований в лабораториях, и, главным образом, на основе примеров, обнаруженных в природе. [ 3 ] : 354 Кроме того, компьютерное моделирование генетики и моделей миграции популяций обнаружило «что-то похожее на подкрепление». [ 3 ] : 372 Самая последняя теоретическая работа по видообразованию была основана на нескольких исследованиях (в частности, Лиу и Прайса, Келли и Нура , а также Киркпатрика и Серведио ) с использованием очень сложного компьютерного моделирования; все они пришли к одинаковым выводам: такое подкрепление возможно при нескольких условиях и во многих случаях оно проще, чем считалось ранее. [ 3 ] : 374
Терминология
[ редактировать ]Существует путаница вокруг значения термина «подкрепление». [ 15 ] Впервые он был использован для описания наблюдаемых различий в брачных криках у лягушек Gastroryne в пределах вторичной контактной гибридной зоны. [ 15 ] Термин «вторичный контакт» также использовался для описания подкрепления в контексте аллопатрически разделенной популяции, испытывающей контакт после потери географического барьера. [ 16 ] Эффект Уоллеса похож на подкрепление, но используется редко. [ 15 ] Роджер Батлин разграничил неполную постзиготическую изоляцию от полной изоляции, ссылаясь на неполную изоляцию как на усиление, а на полностью изолированные популяции — как на испытывающее смещение репродуктивного характера . [ 17 ] Дэниел Дж. Ховард считал, что смещение репродуктивных признаков представляет собой либо ассортативное спаривание , либо расхождение признаков для распознавания партнера (особенно между симпатрическими популяциями). [ 15 ] Подкрепление, по его определению, включало презиготическую дивергенцию и полную постзиготическую изоляцию. [ 18 ] Серведио и Нур включают любое обнаруженное увеличение презиготической изоляции в качестве подкрепления, если оно является реакцией на отбор против спаривания между двумя разными видами. [ 4 ] Койн и Орр утверждают, что «истинное подкрепление ограничивается случаями, когда усиливается изоляция между таксонами, которые все еще могут обмениваться генами». [ 3 ] : 352
Модели
[ редактировать ]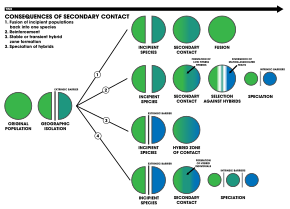
1. Внешний барьер разделяет популяцию вида на две части, но они вступают в контакт до того, как репродуктивная изоляция станет достаточной, чтобы привести к видообразованию. Обе популяции снова сливаются в один вид.
2. Видообразование путем подкрепления
3. Две разделенные популяции остаются генетически различными, в то время как гибридные стаи . в зоне контакта образуются
4. Рекомбинация генома приводит к видообразованию двух популяций с появлением дополнительных гибридных видов . Все три вида разделены внутренними репродуктивными барьерами. [ 19 ]
Одной из самых сильных форм репродуктивной изоляции в природе является сексуальная изоляция: черты организмов, связанные с спариванием. [ 20 ] Эта закономерность привела к идее, что, поскольку отбор так сильно влияет на признаки спаривания, он может быть вовлечен в процесс видообразования. [ 20 ] Этот процесс видообразования под влиянием естественного отбора является подкреплением и может происходить при любом способе видообразования. [ 3 ] : 355 ( например, географические способы видообразования или экологическое видообразование [ 21 ] ). Это требует наличия двух сил эволюции, которые влияют на выбор партнера : естественный отбор и поток генов . [ 22 ] Отбор действует как основной двигатель подкрепления, поскольку он отбирает гибридные генотипы с низкой приспособленностью , независимо от того, не оказывают ли индивидуальные предпочтения никакого влияния на выживание и воспроизводство. [ 22 ] Поток генов действует как основная сила, противостоящая подкреплению, поскольку обмен генами между особями, ведущий к гибридам, приводит к гомогенизации генотипов . [ 22 ]
Батлин выделил четыре основных критерия обнаружения подкрепления в естественных или лабораторных популяциях: [ 17 ]
- Поток генов между двумя таксонами существует или может быть установлено, что он существовал в какой-то момент.
- Между двумя таксонами наблюдается расхождение признаков, связанных со спариванием.
- Способы спаривания изменяются, что ограничивает производство гибридов с низкой приспособленностью.
- Других факторов отбора, ведущих к расхождению системы распознавания партнера, не произошло.
После того, как происходит видообразование путем подкрепления, изменения после полной репродуктивной изоляции (и дальнейшей изоляции после нее) представляют собой форму смещения репродуктивных признаков . [ 23 ] Общим признаком появления подкрепления в природе является смещение репродуктивного характера ; характеристики населения расходятся в симпатии, но не в аллопатрии. [ 6 ] [ 5 ] Одна из трудностей обнаружения заключается в том, что смещение экологического характера может привести к тем же закономерностям. [ 24 ] Кроме того, поток генов может уменьшить изоляцию, наблюдаемую в симпатрических популяциях. [ 24 ] На исход процесса влияют два важных фактора: 1) специфические механизмы, вызывающие презиготную изоляцию, и 2) количество аллелей, измененных мутациями, влияющими на выбор партнера. [ 25 ]
В случаях перипатрического видообразования подкрепление вряд ли приведет к завершению видообразования в случае, если периферически изолированная популяция вступает во вторичный контакт с основной популяцией. [ 26 ] При симпатрическом видообразовании необходим отбор против гибридов; поэтому подкрепление может сыграть свою роль, учитывая эволюцию той или иной формы компромисса в отношении приспособленности. [ 1 ] При симпатии часто наблюдаются модели сильной дискриминации при спаривании, которые объясняются подкреплением. [ 7 ] Считается, что подкрепление является агентом гаметической изоляции. [ 27 ]
Генетика
[ редактировать ]Основную генетику подкрепления можно понять на примере идеальной модели двух гаплоидных популяций, испытывающих усиление неравновесия по сцеплению . Здесь отбор отвергает низкую приспособленность или комбинации аллелей , отдавая предпочтение комбинациям аллели (в первой субпопуляции) и аллели (во второй субпопуляции). Третий локус или (ассортативные аллели спаривания) влияют на характер спаривания, но не находятся под прямым отбором. Если выбор в и вызывают изменения частоты аллеля , способствует ассортативному спариванию, что приводит к подкреплению. Необходимы как отбор, так и ассортативное спаривание, то есть спаривание и встречаются чаще, чем спаривания и . [ 28 ] Ограничение миграции между популяциями может еще больше увеличить вероятность подкрепления, поскольку снижает вероятность обмена разными генотипами. [ 15 ]












Существует альтернативная модель для устранения антагонизма рекомбинации , поскольку она может уменьшить связь между аллелями, которые связаны с приспособленностью, и аллелями ассортативного спаривания, которые этого не делают. [ 15 ] Генетические модели часто различаются по количеству признаков, связанных с локусами ; [ 29 ] некоторые полагаются на один локус на каждый признак [ 26 ] [ 30 ] [ 31 ] и другие по полигенным признакам. [ 23 ] [ 22 ] [ 32 ]
Структура населения
[ редактировать ]Структура и модели миграции популяции могут влиять на процесс видообразования путем подкрепления. Было показано, что это происходит в рамках островной модели, где существуют условия с нечастыми миграциями, происходящими в одном направлении. [ 22 ] и в моделях симметричной миграции, где виды равномерно мигрируют между популяциями. [ 26 ] [ 30 ]
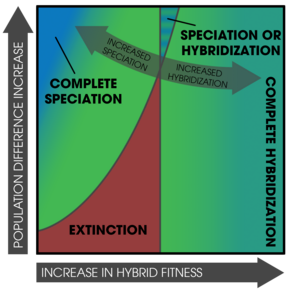
Подкрепление также может происходить в отдельных популяциях. [ 29 ] [ 23 ] мозаичные гибридные зоны (пятнистое распределение родительских форм и субпопуляций), [ 31 ] и в парапатрических популяциях с узкими контактными зонами. [ 33 ]
Плотность населения является важным фактором усиления, часто в сочетании с вымиранием . [ 23 ] Не исключено, что при вторичном контакте двух видов одна популяция может вымереть — прежде всего из-за низкой приспособленности гибридов , сопровождающейся высокими темпами роста популяции. [ 23 ] Вымирание менее вероятно, если гибриды нежизнеспособны , а не бесплодны , поскольку плодовитые особи все еще могут выжить достаточно долго, чтобы размножаться. [ 23 ]
Выбор
[ редактировать ]Видообразование путем подкрепления напрямую зависит от отбора, способствующего усилению презиготной изоляции. [ 1 ] и природа роли отбора в подкреплении широко обсуждалась с моделями, применяющими различные подходы. [ 29 ] Отбор, действующий на гибриды, может происходить несколькими различными способами. Все полученные гибриды могут быть равнонизкопригодными, [ 23 ] давая широкий недостаток. В других случаях отбор может благоприятствовать множественным и различным фенотипам. [ 26 ] например, в случае мозаичной гибридной зоны. [ 31 ] Естественный отбор может действовать на определенные аллели как прямо , так и косвенно. [ 29 ] [ 22 ] [ 34 ] При прямом отборе частота выбранного аллеля максимально повышается. В случаях, когда аллель выбирается косвенно, ее частота увеличивается из-за того, что другой связанный аллель подвергается селекции ( неравновесие по сцеплению ). [ 15 ]
Состояние отбираемых гибридов может играть роль в постзиготической изоляции, поскольку нежизнеспособность гибрида (гибрид, неспособный вырасти в пригодного взрослого человека) и бесплодие (неспособность полностью производить потомство) препятствуют потоку генов между популяциями. [ 7 ] Отбор против гибридов может быть вызван даже любой неудачей в получении партнера, поскольку это фактически неотличимо от бесплодия - любое обстоятельство не приводит к отсутствию потомства. [ 7 ]
Спаривание и предпочтения партнера
[ редактировать ]Для того чтобы произошло подкрепление, должно присутствовать некоторое первоначальное расхождение в предпочтениях партнера. [ 7 ] [ 23 ] [ 35 ] Любые признаки, способствующие изоляции, могут быть подвергнуты усилению, например, сигналы спаривания ( например, демонстрация ухаживания ), сигнальные реакции, расположение мест размножения , время спаривания ( например, сезонное размножение, например, при аллохронном видообразовании ) или даже восприимчивость к яйцам. [ 15 ] Особи могут также дискриминировать партнеров, которые различаются по различным признакам, таким как брачный призыв или морфология . [ 36 ] Многие из этих примеров описаны ниже.
Доказательство
[ редактировать ]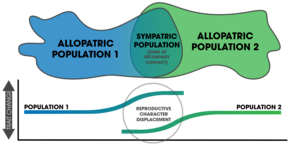
Доказательства подкрепления основаны на наблюдениях в природе, сравнительных исследованиях и лабораторных экспериментах. [ 3 ] : 354
Природа
[ редактировать ]Можно показать, что подкрепление происходит (или происходило в прошлом), измеряя силу презиготической изоляции в симпатрической популяции по сравнению с аллопатрической популяцией того же вида. [ 3 ] : 357 Сравнительные исследования этого позволяют выявить крупномасштабные закономерности в природе по различным таксонам. [ 3 ] : 362 Модели спаривания в гибридных зонах также можно использовать для обнаружения подкрепления. [ 18 ] Репродуктивное смещение характера рассматривается как результат подкрепления, [ 7 ] очень многие случаи в природе выражают эту закономерность в сочувствии. Вездесущность подкрепления неизвестна, [ 4 ] но закономерности смещения репродуктивных признаков обнаруживаются у многих таксонов и считаются обычным явлением в природе. [ 18 ] Исследования подкрепления в природе часто оказываются трудными, поскольку можно предложить альтернативные объяснения обнаруженным закономерностям. [ 3 ] : 358 Тем не менее, существуют эмпирические доказательства того, что подкрепление происходит в различных таксонах. [ 7 ] и его роль в ускорении видообразования неоспорима. [ 15 ]
Сравнительные исследования
[ редактировать ]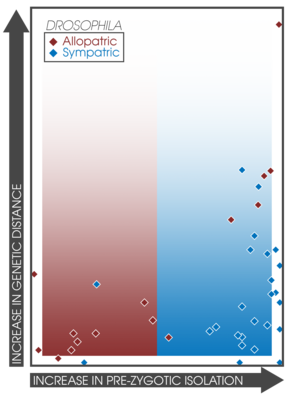
Ожидается, что ассортативное спаривание увеличится среди симпатрических популяций, испытывающих подкрепление. [ 15 ] Этот факт позволяет провести прямое сравнение силы презиготической изоляции при симпатрии и аллопатрии между различными экспериментами и исследованиями. [ 3 ] : 362 Койн и Орр исследовали 171 пару видов, собрав данные об их географическом положении, генетическом расстоянии и силе как презиготической, так и постзиготической изоляции; обнаружили, что презиготическая изоляция была значительно сильнее в симпатрических парах, что коррелирует с возрастом вида. [ 3 ] : 362 Кроме того, сила постзиготической изоляции не различалась между симпатрическими и аллопатрическими парами. [ 15 ] Этот вывод подтверждает предсказания видообразования путем подкрепления и хорошо коррелирует с более поздними исследованиями. [ 18 ] которые обнаружили 33 исследования, демонстрирующие паттерны сильной презиготической изоляции при симпатрии . [ 3 ] : 363 Исследование скорости видообразования у рыб и связанных с ними гибридных зон обнаружило сходные закономерности в симпатии, что подтверждает наличие подкрепления. [ 38 ]
Лабораторные эксперименты
[ редактировать ]Лабораторные исследования, которые явно проверяют наличие подкрепления, ограничены. [ 3 ] : 357 при этом многие эксперименты проводились на дрозофилы плодовых мушках . В целом проводилось два типа экспериментов: использование искусственного отбора для имитации естественного отбора, устраняющего гибриды (часто называемого «уничтожение гибридов»), и использование разрушительного отбора для отбора признака (независимо от его функции в половом отношении). воспроизводство). [ 3 ] : 355–357 Многие эксперименты с использованием метода уничтожения гибридов обычно считаются поддерживающими подкрепление; однако некоторые исследователи, такие как Койн и Орр, Уильям Р. Райс и Эллен Э. Хостерт, утверждают, что они на самом деле не моделируют подкрепление, поскольку поток генов между двумя популяциями полностью ограничен. [ 39 ] [ 3 ] : 356
Альтернативные гипотезы
[ редактировать ]Были предложены различные альтернативные объяснения закономерностей, наблюдаемых в природе. [ 3 ] : 375 Не существует единого всеобъемлющего признака подкрепления; однако есть две предлагаемые возможности: [ 3 ] : 379 половая асимметрия (когда женщины в симпатрических популяциях вынуждены становиться разборчивыми перед лицом двух разных мужчин) [ 40 ] и аллельное доминирование : любая из аллелей, подвергающихся отбору для выделения, должна доминировать. [ 7 ] Хотя эта подпись не полностью учитывает вероятности фиксации или смещения экологического характера . [ 3 ] : 380 Койн и Орр расширяют понятие половой асимметрии и утверждают, что, независимо от изменений, наблюдаемых у женщин и мужчин в симпатии, изоляция в большей степени вызвана женщинами. [ 3 ] : 380
Экологические или этологические влияния
[ редактировать ]Экология также может играть роль в наблюдаемых закономерностях, называемых смещением экологического характера. Естественный отбор может способствовать уменьшению перекрытия ниш между видами вместо того, чтобы уменьшать гибридизацию. [ 3 ] : 377 Хотя один эксперимент с колюшкой , который явно проверял эту гипотезу, не нашел никаких доказательств. [ 41 ]
Взаимодействие видов также может привести к смещению репродуктивных признаков (как в предпочтениях партнера, так и в сигнале спаривания). [ 20 ] Примеры включают давление хищников и конкуренции, паразитов , обманное опыление и мимикрию . [ 20 ] Поскольку эти и другие факторы могут привести к смещению репродуктивного характера, Конрад Дж. Хоскин и Меган Хигги выделяют пять критериев подкрепления, позволяющих различать экологические и этологические влияния:
(1) у очагового вида выявлены признаки спаривания; (2) на признаки спаривания влияет взаимодействие видов, поэтому вероятен отбор по признакам спаривания; (3) взаимодействия видов различаются в разных популяциях (присутствуют или отсутствуют или различные взаимодействия видов влияют на признаки спаривания в каждой популяции); (4) признаки спаривания (сигналы и/или предпочтения) различаются среди популяций из-за различий во взаимодействии видов; (5) видообразование требует демонстрации того, что расхождение признаков спаривания приводит к полной или почти полной половой изоляции среди популяций. Результаты будут наиболее информативными в хорошо изученных биогеографических условиях, где известны взаимоотношения и история популяций. [ 20 ]
Слияние
[ редактировать ]Вполне возможно, что модель усиленной изоляции может быть просто временным результатом вторичного контакта, когда два аллопатрических вида уже имеют различный диапазон презиготической изоляции: у некоторых она проявляется больше, чем у других. [ 42 ] Те, у кого более слабая презиготическая изоляция, в конечном итоге сливаются, теряя свою отличительность. [ 7 ] Эта гипотеза не объясняет тот факт, что отдельные виды в аллопатрии, испытывающие постоянный поток генов, не будут различаться по уровням потока генов при вторичном контакте. [ 7 ] [ 43 ] Более того, закономерности, обнаруженные у дрозофилы, обнаруживают высокие уровни презиготической изоляции в симпатрии, но не в аллопатрии. [ 44 ] Гипотеза слияния предсказывает, что сильная изоляция должна быть обнаружена как в аллопатрии, так и в симпатрии. [ 44 ] Считается, что этот процесс слияния происходит в природе, но он не полностью объясняет закономерности, обнаруженные при подкреплении. [ 3 ] : 376
Сочувствие
[ редактировать ]
Вполне возможно, что сам процесс симпатрического видообразования может привести к наблюдаемым закономерностям подкрепления. [ 3 ] : 378 Один из методов различения между ними - построить филогенетическую историю вида, поскольку сила презиготической изоляции между группой родственных видов должна различаться в зависимости от того, как они видоизменились в прошлом. [ 45 ] Два других способа определить, происходит ли подкрепление (в отличие от симпатрического видообразования):
- если два недавно образовавшихся таксона не обнаруживают признаков постзиготической изоляции как симпатрических, так и аллопатрических популяций (при симпатрическом видообразовании постзиготическая изоляция не является обязательным условием); [ 46 ]
- если клина существует между двумя видами по ряду признаков (симпатрическое видообразование вообще не требует существования клины). [ 47 ]
Половой отбор
[ редактировать ]В процессе безудержного спаривания (мало чем отличающегося от фишеровского отбора ) отбор против гибридов с низкой приспособленностью благоприятствует ассортативному спариванию, быстро увеличивая распознавание партнера. [ 7 ] [ 44 ] Кроме того, когда предпочтения самок обходятся низкой ценой, могут возникнуть изменения в мужских фенотипах, выражающие закономерность, идентичную закономерности смещения репродуктивных признаков. [ 48 ] Постзиготическая изоляция не требуется, поскольку она инициируется просто тем фактом, что непригодные гибриды не могут получить партнеров. [ 7 ]
Аргументы против усиления
[ редактировать ]Был выдвинут ряд возражений, главным образом в 1980-е годы, в которых утверждалось, что усиление неправдоподобно. [ 7 ] [ 20 ] [ 3 ] : 369 Большинство из них полагаются на теоретические работы, которые предполагают, что антагонизм между силами естественного отбора и потоком генов был самым большим препятствием на пути его реализации. [ 3 ] : 369–372 С тех пор эти возражения были в значительной степени опровергнуты данными природы. [ 18 ] [ 3 ] : 372
Поток генов
[ редактировать ]Опасения по поводу того, что гибридная приспособленность играет роль в подкреплении, привели к возражениям, основанным на взаимосвязи между отбором и рекомбинацией. [ 5 ] [ 3 ] : 369 То есть, если поток генов не равен нулю (если гибриды не полностью непригодны), отбор не может привести к фиксации аллелей для презиготической изоляции. [ 28 ] Например: Если население имеет презиготный изолирующий аллель и постзиготические аллели высокой приспособленности и ; и население имеет презиготический аллель а и постзиготические аллели высокой приспособленности и , оба и генотипы будут подвергаться рекомбинации перед лицом потока генов. Каким-то образом популяцию нужно поддерживать. [ 3 ] : 369






Кроме того, специфические аллели, обладающие селективным преимуществом в перекрывающихся популяциях, полезны только внутри этой популяции. [ 49 ] Однако, если они избирательно выгодны, поток генов должен позволить аллелям распространиться в обеих популяциях. [ 49 ] Чтобы предотвратить это, аллели должны быть вредными или нейтральными. [ 3 ] : 371 Это не без проблем, поскольку поток генов из предположительно крупных аллопатрических регионов может захлестнуть территорию, когда две популяции перекрываются. [ 3 ] : 371 Чтобы подкрепление работало, поток генов должен присутствовать, но очень ограниченно. [ 26 ] [ 31 ]
Недавние исследования показывают, что подкрепление может происходить в более широком диапазоне условий, чем считалось ранее. [ 29 ] [ 46 ] [ 3 ] : 372–373 и что эффект потока генов можно преодолеть путем отбора. [ 50 ] [ 51 ] Например, два вида Drosophila santomea и D. yakuba на африканском острове Сан-Томе иногда гибридизируются друг с другом, в результате чего образуются плодовитые потомки женского пола и бесплодные потомки мужского пола. [ 50 ] Эта естественная ситуация была воспроизведена в лаборатории, напрямую моделируя усиление: удаление некоторых гибридов и допущение различных уровней потока генов. [ 51 ] Результаты эксперимента убедительно свидетельствуют о том, что подкрепление работает в различных условиях: эволюция половой изоляции возникает в 5–10 поколениях плодовых мух. [ 51 ]
Быстрые требования
[ редактировать ]В сочетании с гипотезой слияния подкрепление можно рассматривать как гонку против слияния и вымирания. [ 42 ] Производство непригодных гибридов фактически равносильно недостатку гетерозигот ; при этом отклонение от генетического равновесия приводит к потере непригодного аллеля. [ 52 ] Этот эффект приведет к исчезновению одной из популяций. [ 53 ] Это возражение преодолевается, если обе популяции не находятся в одинаковых экологических условиях. [ 3 ] : 370 Тем не менее, вымирание одной популяции все еще возможно, и это было показано в моделировании популяций. [ 54 ] Чтобы произошло подкрепление, презиготическая изоляция должна произойти быстро. [ 3 ] : 370
Ссылки
[ редактировать ]- ^ Jump up to: а б с Ханнес Шулер, Глен Р. Худ, Скотт П. Иган и Джеффри Л. Федер (2016), Мейерс, Роберт А. (редактор), «Способы и механизмы видообразования» , Обзоры по клеточной биологии и молекулярной медицине , 2 (3 ): 60–93, номер домена : 10.1002/3527600906 , ISBN. 9783527600908
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Джереми Л. Маршалл, Майкл Л. Арнольд и Дэниел Дж. Ховард (2002), «Укрепление: путь не выбран», Trends in Ecology & Evolution , 17 (12): 558–563, doi : 10.1016/S0169-5347 (02)02636-8
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а Джерри А. Койн ; Х. Аллен Орр (2004), Видообразование , Sinauer Associates, стр. 1–545, ISBN 978-0-87893-091-3
- ^ Jump up to: а б с Мария Р. Серведио; Мохамед А. Ф. Нур (2003), «Роль подкрепления в видообразовании: теория и данные», Ежегодный обзор экологии, эволюции и систематики , 34 : 339–364, doi : 10.1146/annurev.ecolsys.34.011802.132412
- ^ Jump up to: а б с Дэниел Ортис-Барриентос, Алисия Грили и Патрик Носил (2009), «Генетика и экология подкрепления: последствия для эволюции презиготической изоляции в симпатрии и за ее пределами», Анналы Нью-Йоркской академии наук , 1168 : 156–182 , дои : 10.1111/j.1749-6632.2009.04919.x , PMID 19566707 , S2CID 4598270
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с Мария Р. Серведио (2004), «Что и почему исследования подкрепления», PLOS Biology , 2 (12): 2032–2035, doi : 10.1371/journal.pbio.0020420 , PMC 535571 , PMID 15597115
- ^ Jump up to: а б с д и ж г час я дж к л м Мохамед А. Ф. Нур (1999), «Подкрепление и другие последствия симпатии», Наследственность , 83 (5): 503–508, doi : 10.1038/sj.hdy.6886320 , PMID 10620021 , S2CID 26625194
- ^ Роджер К. Батлин и Кэрол М. Смаджа (2018), «Соединение, подкрепление и видообразование» (PDF) , The American Naturalist , 191 (2): 155–172, doi : 10.1086/695136 , PMID 29351021 , S2CID 3397377
- ^ Уоллес, Альфред Рассел (1889). Дарвинизм . Macmillan & Co., стр. 174–179.
- ^ MJ Литтлджон (1981). Репродуктивная изоляция: критический обзор. В книге WR Atchley и DS Woodruff (ред.) «Эволюция и видообразование» , Cambridge University Press, стр. 298–334.
- ^ Марио А. Фарес (2015), Естественный отбор: методы и приложения , CRC Press, стр. 3, ISBN 9781482263725
- ^ Блэр, В. Франк (1955), «Брачный призыв и стадия видообразования в комплексе Microhyla olivacea-M. carolinensis », Evolution , 9 (4): 469–480, doi : 10.1111/j.1558-5646.1955.tb01556. х , S2CID 88238743
- ^ Стэнли Сойер и Дэниел Хартл (1981), «Об эволюции поведенческой репродуктивной изоляции: эффект Уоллеса», Theoretical Population Biology , 19 (1): 261–273, doi : 10.1016/0040-5809(81)90021-6
- ^ Дж. А. Свед (1981), «Двуполая полигенная модель эволюции изоляции предматирующих. I. Детерминистическая теория для природных популяций», Genetics , 97 (1): 197–215, doi : 10.1093/genetics/97.1.197 , ПМК 1214384 , ПМИД 17249073
- ^ Jump up to: а б с д и ж г час я дж к Гленн-Питер Сэтре (2012). «Укрепление». Энциклопедия наук о жизни . дои : 10.1002/9780470015902.a0001754.pub3 . ISBN 978-0470016176 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Добжанский, Феодосий (1937). Генетика и происхождение видов . Издательство Колумбийского университета.
- ^ Jump up to: а б Батлин, Роджер К. (1989). «Усиление прематической изоляции» . Ин Отте, Д.; Эндлер, Джон А. (ред.). Видообразование и его последствия . Синауэр Ассошиэйтс. стр. 158–179 . ISBN 978-0-87893-657-1 .
- ^ Jump up to: а б с д и Ховард, Дэниел Дж. (1993). «Подкрепление: происхождение, динамика и судьба эволюционной гипотезы». В Харрисоне, Р.Г. (ред.). Гибридные зоны и эволюционный процесс . Издательство Оксфордского университета. стр. 46–69. ISBN 978-0-19-506917-4 .
- ^ Джон А. Хвала и Трой Э. Вуд (2012). «Видение: Введение». Энциклопедия наук о жизни . дои : 10.1002/9780470015902.a0001709.pub3 . ISBN 978-0470016176 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б с д и ж Конрад Дж. Хоскин и Меган Хигги (2010), «Видообразование посредством взаимодействия видов: расхождение признаков спаривания внутри видов», Ecology Letters , 13 (4): 409–420, Bibcode : 2010EcolL..13..409H , doi : 10.1111/j.1461-0248.2010.01448.x , PMID 20455922 , S2CID 16175451
- ^ Марк Киркпатрик (2001), «Подкрепление во время экологического видообразования», Proceedings of the Royal Society B , 268 (1473): 1259–1263, doi : 10.1098/rspb.2000.1427 , PMC 1088735 , PMID 11410152
- ^ Jump up to: а б с д и ж Марк Киркпатрик и Мария Р. Серведио (1999), «Усиление брачных предпочтений на острове», Genetics , 151 (2): 865–884, doi : 10.1093/genetics/151.2.865 , PMC 1460501 , PMID 9927476
- ^ Jump up to: а б с д и ж г час я Лили В. Лиу и Тревор Д. Прайс (1994), «Видение путем усиления изоляции перед сматыванием», Evolution , 48 (5): 1451–1459, doi : 10.1111/j.1558-5646.1994.tb02187.x , PMID 28568419 , S2CID 22630822
- ^ Jump up to: а б Сеть Марии Кюри СПЕЦИАЦИЯ (2012 г.), «Что нам нужно знать о видообразовании?» , Тенденции в экологии и эволюции , 27 (1): 27–39, doi : 10.1016/j.tree.2011.09.002 , PMID 21978464
- ^ Клаудия Бэнк, Иоахим Хермиссия и Марк Киркпатрик (2012), «Может ли подкрепление завершить видообразование?», Evolution , 66 (1): 229–239, doi : 10.1111/j.1558-5646.2011.01423.x , PMID 22220877 , S2CID 15602575
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с д и Мария Р. Серведио и Марк Киркпатрик (1997), «Влияние потока генов на подкрепление», Evolution , 51 (6): 1764–1772, doi : 10.1111/j.1558-5646.1997.tb05100.x , PMID 28565111 , S2CID 12269299
- ^ Дэниел Р. Матуте (2010), «Усиление гаметической изоляции у дрозофилы », PLOS Biology , 8 (6): e1000341, doi : 10.1371/journal.pbio.1000341 , PMC 2843595 , PMID 20351771
- ^ Jump up to: а б Джозеф Фельзенштейн (1981), «Скептицизм в отношении Санта-Розалии, или Почему так мало видов животных?», Evolution , 35 (1): 124–138, doi : 10.2307/2407946 , JSTOR 2407946 , PMID 28563447
- ^ Jump up to: а б с д и Майкл Турелли; Николас Х. Бартон; Джерри А. Койн (2001), «Теория и видообразование», Тенденции в экологии и эволюции , 16 (7): 330–343, doi : 10.1016/S0169-5347(01)02177-2 , PMID 11403865
- ^ Jump up to: а б Мария Р. Серведио (2000), «Подкрепление и генетика неслучайного спаривания», Evolution , 54 (1): 21–29, doi : 10.1111/j.0014-3820.2000.tb00003.x , PMID 10937179 , S2CID 12563023
- ^ Jump up to: а б с д Майкл Л. Кейн, Вигго Андреасен и Дэниел Дж. Ховард (1999), «Усиливающий отбор эффективен в относительно широком наборе условий в мозаичной гибридной зоне», Evolution , 53 (5): 1343–1353, doi : 10.1111. /j.1558-5646.1999.tb05399.x , PMID 28565558 , S2CID 31107731
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Марк Киркпатрик (2000), «Подкрепление и расхождение при ассортативном спаривании», Proceedings of the Royal Society B , 267 (1453): 1649–1655, doi : 10.1098/rspb.2000.1191 , PMC 1690725 , PMID 11467428
- ^ Нил Сандерсон (1989), «Может ли поток генов предотвратить подкрепление?», Evolution , 43 (6): 1223–1235, doi : 10.2307/2409358 , JSTOR 2409358 , PMID 28564502
- ^ Мария Р. Серведио (2001), «Помимо подкрепления: эволюция изоляции перед спариванием путем прямого отбора по предпочтениям и после спаривания, презиготическая несовместимость», Evolution , 55 (10): 1909–1920, doi : 10.1111/j.0014-3820.2001. tb01309.x , PMID 11761053 , S2CID 25296147
- ^ Дж. Келли и Мохамед А. Ф. Нур (1996), «Видование путем подкрепления: модель, полученная на основе исследований дрозофилы», Genetics , 143 (3): 1485–1497, doi : 10.1093/genetics/143.3.1485 , PMC 1207414 , PMID 8807317
- ^ Конрад Дж. Хоскин, Меган Хигги, Кейт Р. Макдональд и Крейг Мориц (2005), «Подкрепление приводит к быстрому аллопатрическому видообразованию», Nature , 437 (7063): 1353–1356, Бибкод : 2005Natur.437.1353H , doi : 10.1038/ природа04004 , PMID 16251964 , S2CID 4417281
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Джерри А. Койн и Х. Аллен Орр (1997), « Возвращение к закономерностям видообразования у дрозофилы », Evolution , 51 (1): 295–303, doi : 10.1111/j.1558-5646.1997.tb02412.x , PMID 28568795 , S2CID 40390753
- ^ А. Р. МакКьюн и Н. Р. Лавджой. (1998). Относительная скорость симпатрического и аллопатрического видообразования у рыб. В книге DJ Howard и SH Berlocher (ред.) «Бесконечные формы: виды и видообразование» , Oxford University Press, стр. 172–185.
- ^ Уильям Р. Райс и Эллен Э. Хостерт (1993), «Лабораторные эксперименты по видообразованию: чему мы научились за 40 лет?», Evolution , 47 (6): 1637–1653, doi : 10.1111/j.1558-5646.1993. tb01257.x , PMID 28568007 , S2CID 42100751
- ^ Л. Партридж и Г. А. Паркер. (1999). Сексуальный конфликт и видообразование. В А.Е. Магурране и Р.М. Мэе (ред.) «Эволюция биологического разнообразия» . Издательство Оксфордского университета, стр. 130–159.
- ^ Говард Д. Рандл (1998), «Усиление предпочтений партнера-колюшки: симпатия порождает презрение» , Дольф Шлютер , 52 (1): 200–208, doi : 10.1111/j.1558-5646.1998.tb05153.x , hdl : 2429/ 6366 , ПМИД 28568163 , С2КИД 40648544
- ^ Jump up to: а б Алан Р. Темплтон (1981), «Механизмы видообразования – популяционно-генетический подход», Ежегодный обзор экологии, эволюции и систематики , 12 : 23–48, doi : 10.1146/annurev.es.12.110181.000323
- ^ , обусловленное естественным отбором Мохамед А.Ф. Нур (1995), «Видование у дрозофилы », Nature , 375 (6533): 674–675, Бибкод : 1995Natur.375..674N , doi : 10.1038/375674a0 , PMID 7791899 , S2CID 4252448
- ^ Jump up to: а б с Джерри А. Койн ; Х. Аллен Орр (1989), «Способы видообразования у дрозофилы », Evolution , 43 (2): 362–381, doi : 10.1111/j.1558-5646.1989.tb04233.x , PMID 28568554 , S2CID 1678429
- ^ Jump up to: а б Мохамед А. Ф. Нур (1997), «Как часто симпатия влияет на сексуальную изоляцию у дрозофилы ?», The American Naturalist , 149 (6): 1156–1163, doi : 10.1086/286044 , PMID 18811269 , S2CID 5406442
- ^ Jump up to: а б Марк Киркпатрик и Виржини Равинье (2002), «Видообразование путем естественного и полового отбора: модели и эксперименты», The American Naturalist , 159 : S22–35, doi : 10.1086/338370 , PMID 18707367 , S2CID 16516804
- ^ Н. Х. Бартон и Г. М. Хьюитт (1989), «Адаптация, видообразование и гибридные зоны», Nature , 341 (6242): 497–503, Бибкод : 1989Natur.341..497B , doi : 10.1038/341497a0 , PMID 2677747 , S2CID 4360057
- ^ Трой Дэй (2000), «Половой отбор и эволюция дорогостоящих женских предпочтений: пространственные эффекты», Evolution , 54 (3): 715–730, doi : 10.1554/0014-3820(2000)054[0715:SSATEO]2.3. CO;2 , PMID 10937247 , S2CID 27621958
- ^ Jump up to: а б Дж. А. Мур. (1957). Взгляд эмбриолога на понятие вида. В книге Эрнста Майра (ред.) «Проблема видов» , Американская ассоциация содействия развитию науки, стр. 325–338.
- ^ Jump up to: а б Дэниел Р. Матуте (2010), «Подкрепление может преодолеть поток генов во время видообразования у дрозофилы », Current Biology , 20 (24): 2229–2233, doi : 10.1016/j.cub.2010.11.036 , PMC 3019097 , PMID 21129972
- ^ Jump up to: а б с Джерри А. Койн (2010), «Подкрепление» и происхождение видов , Wordpress
- ^ А. А. Харпер и Д. М. Ламберт (1983), «Популяционная генетика усиливающего отбора», Genetica , 62 (1): 15–23, doi : 10.1007/BF00123305 , S2CID 7947934.
- ^ Х. Х. Патерсон (1978), «Больше доказательств против видообразования путем подкрепления», South African Journal of Science , 74 : 369–371.
- ^ Хэмиш Г. Спенсер, Брайан Х. Макардл и Дэвид М. Ламберт (1986), «Теоретическое исследование видообразования путем подкрепления», The American Naturalist , 128 (2): 241–262, doi : 10.1086/284557 , S2CID 83906561
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка )