Генетическая связь
Генетическая связь — это тенденция последовательностей ДНК , расположенных близко друг к другу на хромосоме , наследоваться вместе во время мейоза фазы полового размножения . Два генетических маркера , которые физически близки друг к другу, вряд ли будут разделены на разные хроматиды во время хромосомного кроссовера , и поэтому говорят, что они более связаны, чем маркеры, которые находятся далеко друг от друга. Другими словами, чем ближе два гена расположены в хромосоме, тем меньше вероятность рекомбинации между ними и тем больше вероятность того, что они будут унаследованы вместе. Маркеры на разных хромосомах совершенно не связаны между собой , хотя на пенетрантность потенциально вредных аллелей может влиять присутствие других аллелей, и эти другие аллели могут располагаться на других хромосомах, а не на той, на которой расположен конкретный потенциально вредный аллель. [1]
Генетическая связь является наиболее заметным исключением из Грегора Менделя Закона Независимого Ассортимента . Первый эксперимент, демонстрирующий связь, был проведен в 1905 году. В то время была неизвестна причина, по которой определенные признаки имеют тенденцию наследоваться вместе. Более поздние работы показали, что гены представляют собой физические структуры, связанные физическим расстоянием.
Типичной единицей генетического сцепления является сантиморган (сМ). Расстояние между двумя маркерами в 1 см означает, что маркеры распределяются по разным хромосомам в среднем один раз на 100 мейотических продуктов, то есть один раз на 50 мейозов.
Открытие [ править ]
Грегора Менделя гласит Закон независимого распределения , что каждый признак наследуется независимо от любого другого признака. Но вскоре после того, как работа Менделя была вновь открыта , были найдены исключения из этого правила. В 1905 году британские генетики Уильям Бейтсон , Эдит Ребекка Сондерс и Реджинальд Паннетт скрестили растения гороха в экспериментах, аналогичных экспериментам Менделя. [2] [3] Они интересовались наследованием признаков у душистого горошка и изучали два гена — ген цвета цветка ( P — фиолетовый и p — красный) и ген, влияющий на форму пыльцевых зерен ( L — длинный и l — круглый). . Они пересекли чистые линии PPLL и ppll , а затем самостоятельно пересекли полученные линии PpLl .
Согласно менделевской генетике , ожидаемые фенотипы будут встречаться в соотношении 9:3:3:1 PL:Pl:pL:pl. К своему удивлению, они наблюдали увеличение частоты PL и pl и снижение частоты PL и pL:
| Фенотип и генотип | Наблюдается | Ожидается соотношение 9:3:3:1 |
|---|---|---|
| Фиолетовый, длинный ( P_L_ ) | 284 | 216 |
| Фиолетовый, круглый ( P_ll ) | 21 | 72 |
| Красный, длинный ( ppL_ ) | 21 | 72 |
| Красный, круглый ( ppll ) | 55 | 24 |
Их эксперимент выявил связь между аллелями P и L и аллелями p и l . Частота встречаемости P вместе с L и p вместе с l выше, чем частота встречаемости рекомбинантных Pl и pL . Частоту рекомбинации труднее вычислить при скрещивании F2, чем при обратном скрещивании. [4] но несоответствие между наблюдаемым и ожидаемым количеством потомства в приведенной выше таблице указывает на то, что оно составляет менее 50%. Это указывало на то, что два фактора каким-то образом взаимодействовали, создавая это различие, маскируя появление двух других фенотипов. Это привело к выводу, что некоторые признаки связаны друг с другом из-за их близкой близости друг к другу на хромосоме.
Понимание связи было расширено работой Томаса Ханта Моргана . Наблюдение Моргана о том, что степень кроссинговера между связанными генами различна, привело к идее, что частота кроссинговера может указывать на расстояние, разделяющее гены на хромосоме . В его честь назван сантиморган , обозначающий частоту кроссинговера.
Карта связей [ править ]
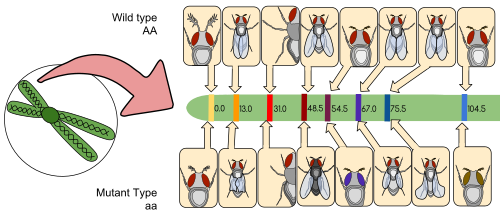
Карта сцепления (также известная как генетическая карта ) — это таблица для вида или экспериментальной популяции, которая показывает положение его известных генов или генетических маркеров относительно друг друга с точки зрения частоты рекомбинации, а не конкретного физического расстояния вдоль каждой хромосомы. . Карты связей были впервые разработаны Альфредом Стёртевантом , учеником Томаса Ханта Моргана .
Карта сцепления — это карта, основанная на частотах рекомбинации между маркерами во время скрещивания гомологичных хромосом . Предполагается, что чем выше частота рекомбинации (расщепления) между двумя генетическими маркерами, тем дальше друг от друга они находятся. И наоборот, чем ниже частота рекомбинации между маркерами, тем меньше физическое расстояние между ними. Исторически сложилось так, что первоначально использовавшимися маркерами были определяемые фенотипы (выработка ферментов, цвет глаз), полученные из кодирующих последовательностей ДНК; подтвержденные или предполагаемые некодирующие последовательности ДНК, такие как микросателлиты или те, которые генерируют полиморфизм длины рестрикционного фрагмента ( RFLP в конечном итоге стали использоваться ).
Карты сцепления помогают исследователям находить другие маркеры, например, другие гены, путем тестирования генетического сцепления уже известных маркеров. На ранних стадиях разработки карты сцепления данные используются для сборки групп сцепления — набора генов, о которых известно, что они связаны между собой. По мере развития знаний к группе можно добавлять больше маркеров, пока группа не охватит всю хромосому. [6] У хорошо изученных организмов группы сцепления взаимно однозначно соответствуют хромосомам.
Карта связей не является физической картой (например, гибридной картой с пониженным уровнем радиации ) или генной картой .
Анализ связей [ править ]
Анализ сцепления — это генетический метод, который ищет хромосомные сегменты, которые разделяют фенотип заболевания в семьях. [7] Его можно использовать для картирования генов как по бинарным, так и по количественным признакам. [7] Анализ сцепления может быть как параметрическим (если мы знаем связь между фенотипическим и генетическим сходством), так и непараметрическим. Параметрический анализ сцепления — это традиционный подход, при котором вероятность того, что ген, важный для заболевания, связан с генетическим маркером, изучается с помощью показателя LOD, который оценивает вероятность того, что данная родословная, в которой заболевание и маркер разделяются друг на друга, из-за существования связи (с заданным значением связи) или случайно. Непараметрический анализ сцепления, в свою очередь, изучает вероятность того, что аллель идентична самой себе по происхождению .

Параметрический анализ связей [ править ]
Показатель LOD (логарифм (по основанию 10) шансов), разработанный Ньютоном Мортоном , [8] представляет собой статистический тест, часто используемый для анализа сцепления в популяциях человека, животных и растений. Показатель LOD сравнивает вероятность получения тестовых данных, если два локуса действительно связаны, с вероятностью наблюдения тех же данных чисто случайно. Положительные оценки LOD свидетельствуют о наличии связи, тогда как отрицательные оценки LOD указывают на то, что связь менее вероятна. Компьютеризированный анализ показателей LOD — это простой способ анализа сложных семейных родословных с целью определения связи между менделевскими признаками (или между признаком и маркером, или двумя маркерами).
Более подробно метод описан Страчаном и Ридом. [1] Вкратце, это работает следующим образом:
- Установить родословную
- Сделайте ряд оценок частоты рекомбинации.
- Рассчитайте показатель LOD для каждой оценки
- Оценка с наивысшим показателем LOD будет считаться лучшей оценкой.
Оценка LOD рассчитывается следующим образом:

NR обозначает количество нерекомбинантного потомства, а R обозначает количество рекомбинантного потомства. Причина, по которой в знаменателе используется 0,5, заключается в том, что любые аллели, которые полностью несвязаны (например, аллели в отдельных хромосомах), имеют 50%-ную вероятность рекомбинации из-за независимого ассортимента. θ представляет собой рекомбинантную фракцию, т.е. долю рождений, в которых произошла рекомбинация между изучаемым генетическим маркером и предполагаемым геном, связанным с заболеванием. Таким образом, оно равно R /( NR + R ) .
По соглашению, показатель LOD более 3,0 считается свидетельством связи, поскольку он указывает на вероятность 1000 к 1, что наблюдаемая связь не возникла случайно. С другой стороны, показатель LOD менее -2,0 считается свидетельством исключения связи. Хотя очень маловероятно, что оценка LOD, равная 3, будет получена на основе одной родословной, математические свойства теста позволяют объединять данные из нескольких родословных путем суммирования их оценок LOD. Показатель LOD, равный 3, соответствует p значению примерно 0,05. [9] и не множественная коррекция тестирования (например, коррекция Бонферрони ). требуется [10]
Ограничения [ править ]
Анализ сцепления имеет ряд методологических и теоретических ограничений, которые могут значительно увеличить частоту ошибок типа 1 и снизить возможности картирования локусов количественных признаков человека (QTL). [11] Хотя анализ сцепления успешно использовался для выявления генетических вариантов, которые способствуют развитию редких заболеваний, таких как болезнь Хантингтона , он не показал столь хороших результатов при применении к более распространенным заболеваниям, таким как болезни сердца или различные формы рака . [12] Объяснение этому состоит в том, что генетические механизмы, влияющие на распространенные заболевания, отличаются от механизмов, вызывающих некоторые редкие заболевания. [13]
Частота рекомбинации [ править ]
Частота рекомбинации является мерой генетического сцепления и используется при создании карты генетического сцепления. Частота рекомбинации ( θ ) — это частота, с которой одиночный хромосомный кроссинговер происходит между двумя генами во время мейоза . Сантиморган (см) — единица измерения, описывающая частоту рекомбинации 1%. Таким образом, мы можем измерить генетическое расстояние между двумя локусами, основываясь на частоте их рекомбинации. Это хорошая оценка реального расстояния. Двойные кроссоверы обернутся никакой рекомбинацией. В этом случае мы не можем сказать, имели ли место кроссоверы. Если локусы, которые мы анализируем, очень близки (менее 7 см), двойной кроссинговер очень маловероятен. Когда расстояния становятся больше, вероятность двойного пересечения возрастает. По мере увеличения вероятности двойного скрещивания можно систематически недооценивать генетическое расстояние между двумя локусами, если не использовать соответствующую математическую модель.
Во время мейоза хромосомы случайным образом распределяются по гаметам , так что разделение аллелей одного гена не зависит от аллелей другого гена. Об этом говорится во втором законе Менделя , который известен как закон независимого ассортимента . Закон независимого ассортимента всегда справедлив для генов, расположенных в разных хромосомах, но для генов, находящихся в одной хромосоме, он справедлив не всегда.
В качестве примера независимого ассортимента рассмотрим скрещивание чистопородной гомозиготной родительской линии с генотипом ААВВ с другой чистопородной линией с генотипом аабб . A, a, B и b представляют собой аллели генов A и B. Скрещивание этих гомозиготных родительских линий приведет к получению потомства поколения F1, которое является двойной гетерозиготой с генотипом AaBb. Потомство F1 AaBb производит гаметы AB , Ab , aB и ab с одинаковой частотой (25%), поскольку аллели гена A сортируются независимо от аллелей гена B во время мейоза. Обратите внимание, что 2 из 4 гамет (50%) — Ab и aB — не присутствовали в родительском поколении. Эти гаметы представляют собой рекомбинантные гаметы . Рекомбинантные гаметы — это гаметы, которые отличаются от обеих гаплоидных гамет, составлявших исходную диплоидную клетку. В этом примере частота рекомбинации составляет 50%, поскольку 2 из 4 гамет были рекомбинантными.
Частота рекомбинации составит 50%, если два гена расположены на разных хромосомах или когда они сильно разнесены на одной хромосоме. Это следствие самостоятельного ассортимента.
Когда два гена расположены близко друг к другу в одной хромосоме, они не сортируются независимо и называются сцепленными. В то время как гены, расположенные на разных хромосомах, сортируются независимо и имеют частоту рекомбинации 50%, сцепленные гены имеют частоту рекомбинации менее 50%.
В качестве примера связи рассмотрим классический эксперимент Уильяма Бейтсона и Реджинальда Паннета . [14] Они интересовались наследованием признаков у душистого горошка и изучали два гена — ген цвета цветка ( P — фиолетовый и p — красный) и ген, влияющий на форму пыльцевых зерен ( L — длинный и l — круглый). . Они пересекли чистые линии PPLL и ppll , а затем самостоятельно пересекли полученные линии PpLl . Согласно менделевской генетике , ожидаемые фенотипы будут встречаться в соотношении 9:3:3:1 PL:Pl:pL:pl. К своему удивлению, они наблюдали увеличение частоты PL и pl и снижение частоты PL и pL (см. таблицу ниже).
| Фенотип и генотип | Наблюдается | Ожидается соотношение 9:3:3:1 |
|---|---|---|
| Фиолетовый, длинный ( P_L_ ) | 284 | 216 |
| Фиолетовый, круглый ( P_ll ) | 21 | 72 |
| Красный, длинный ( ppL_ ) | 21 | 72 |
| Красный, круглый ( ppll ) | 55 | 24 |

Их эксперимент выявил связь между аллелями P и L и аллелями p и l . Частота встречаемости P вместе с L и p вместе с l выше, чем частота встречаемости рекомбинантных Pl и pL . Частоту рекомбинации труднее вычислить при скрещивании F2, чем при обратном скрещивании. [4] но несоответствие между наблюдаемым и ожидаемым количеством потомства в приведенной выше таблице указывает на то, что оно составляет менее 50%.
Потомство в этом случае получило два доминантных аллеля, сцепленных на одной хромосоме (так называемое сцепление или цис-расположение ). Однако после скрещивания некоторое потомство могло получить одну родительскую хромосому с доминантным аллелем одного признака (например, пурпурного), сцепленным с рецессивным аллелем второго признака (например, круглой), тогда как для другой родительской хромосомы (например, красной) было бы наоборот. и Лонг). Это называется отталкиванием или транс-расположением . Фенотип здесь по-прежнему будет фиолетовым и длинным , но тестовое скрещивание этой особи с рецессивным родителем даст потомство с гораздо большей долей двух фенотипов скрещивания. Хотя такая проблема может показаться маловероятной из этого примера, неблагоприятные отталкивающие связи действительно появляются при селекции на устойчивость к болезням у некоторых культур.
Два возможных расположения аллелей, цис и транс, в двойной гетерозиготе называются гаметическими фазами , а фазирование — это процесс определения того, какая из двух присутствует у данного индивидуума.
Когда два гена расположены в одной хромосоме, вероятность кроссовера , вызывающего рекомбинацию между генами, связана с расстоянием между двумя генами. Таким образом, использование частот рекомбинации использовалось для разработки карт сцепления или генетических карт .
Однако важно отметить, что частота рекомбинации имеет тенденцию недооценивать расстояние между двумя сцепленными генами. Это связано с тем, что, поскольку два гена расположены дальше друг от друга, вероятность двойного или даже количества кроссинговеров между ними также увеличивается. Двойное или четное количество кроссинговеров между двумя генами приводит к тому, что они сегрегируются в одной и той же гамете, давая родительское потомство вместо ожидаемого рекомбинантного потомства. Как упоминалось выше, преобразования Косамби и Холдейна пытаются исправить множественные кроссоверы. [15]
Связывание генетических участков внутри гена [ править ]
В начале 1950-х годов преобладала точка зрения, что гены в хромосоме представляют собой дискретные сущности, неделимые в результате генетической рекомбинации и расположенные как бусинки на нитке. В период с 1955 по 1959 год Бензер проводил по генетической рекомбинации эксперименты с использованием rII- мутантов бактериофага Т4 . Он обнаружил, что на основе тестов рекомбинации сайты мутаций можно картировать в линейном порядке. [16] [17] Этот результат подтвердил ключевую идею о том, что ген имеет линейную структуру, эквивалентную длине ДНК, со многими сайтами, которые могут независимо мутировать.
Эдгар и др. [18] провели эксперименты по картированию с r-мутантами бактериофага Т4, показавшие, что частоты рекомбинации между rII-мутантами не являются строго аддитивными. Частота рекомбинации от скрещивания двух мутантов rII (axd) обычно меньше суммы частот рекомбинации соседних внутренних подинтервалов (axb) + (bxc) + (cxd). Хотя это и не является строго аддитивным, наблюдалась систематическая связь. [19] это, вероятно, отражает основной молекулярный механизм генетической рекомбинации .
Изменение частоты рекомбинации [ править ]
Хотя рекомбинация хромосом является важным процессом во время мейоза, существует широкий диапазон частоты кроссоверов между организмами и внутри видов. Скорость рекомбинации полового диморфизма называется гетерохиазмией и наблюдается чаще, чем обычная скорость между мужчинами и женщинами. У млекопитающих самки часто имеют более высокий уровень рекомбинации по сравнению с самцами. Предполагается, что существуют уникальные факторы отбора, или движущие силы мейоза, которые влияют на разницу в скоростях. Разница в скоростях может также отражать совершенно разные среды и условия мейоза в оогенезе и сперматогенезе. [20]
рекомбинации влияющие на частоту , Гены
Мутации в генах , кодирующих белки, участвующие в процессинге ДНК, часто влияют на частоту рекомбинации . В бактериофаге Т4 мутации, которые снижают экспрессию репликативной ДНК-полимеразы [продукт гена 43 (gp43)], увеличивают рекомбинацию (уменьшают связь) в несколько раз. [21] [22] Увеличение рекомбинации может быть связано с ошибками репликации , вызванными дефектной ДНК-полимеразой, которые сами по себе являются событиями рекомбинации, такими как переключение матрицы, то есть события рекомбинации с выбором копии. [23] Рекомбинация также усиливается мутациями, которые снижают экспрессию ДНК-лигазы (gp30). [24] [22] и гидроксиметилаза dCMP (gp42), [21] [22] два фермента, участвующих в синтезе ДНК .
Рекомбинация снижается (увеличивается сцепление) за счет мутаций в генах, кодирующих белки с нуклеазы (gp46 и gp47). функциями [24] [22] и ДНК-связывающий белок (gp32) [22] Мутация гена бактериофага uvsX также существенно снижает рекомбинацию. [25] Ген uvsX аналогичен хорошо изученному RecA гену Escherichia coli , который играет центральную роль в рекомбинации. [26]
Индикаторы мейоза [ править ]
Имея очень большие родословные или очень плотные данные о генетических маркерах, например, полученные в результате полногеномного секвенирования, можно точно определить местонахождение рекомбинаций. При этом типе генетического анализа индикатор мейоза присваивается каждой позиции генома для каждого мейоза в родословной. Индикатор указывает, какая копия родительской хромосомы способствует передаче гаметы в этом положении. Например, если передается аллель из «первой» копии родительской хромосомы, этому мейозу может быть присвоен «0». Если передается аллель из «второй» копии родительской хромосомы, этому мейозу будет присвоена цифра «1». Два аллеля у родителя произошли, по одному, от двух бабушек и дедушек. Эти индикаторы затем используются для определения состояний идентичности по происхождению (IBD) или состояний наследования, которые, в свою очередь, используются для идентификации генов, ответственных за заболевания.
См. также [ править ]
Ссылки [ править ]
- ^ Купер, Д.Н.; Кравчак, М; Полихронакос, К; Тайлер-Смит, центральный; Керер-Савацки, Х. (октябрь 2013 г.). «Там, где генотип не предсказывает фенотип: к пониманию молекулярной основы пониженной пенетрантности при наследственных заболеваниях человека» . Генетика человека . 132 (10): 1077–130. дои : 10.1007/s00439-013-1331-2 . ПМЦ 3778950 . ПМИД 23820649 .
- ^ Лобо, Ингрид; Шоу, Кенна. «Открытие и типы генетического сцепления» . Возбудимый . Природное образование . Проверено 21 января 2017 г.
- ^ Бейтсон, В ; Сондерс, ER ; Паннетт, Р.К. (18 мая 1904 г.). Отчеты перед комитетом по эволюции Королевского общества . Лондон: Харрисон и сыновья, принтеры . Проверено 21 января 2017 г.
- ^ Jump up to: Перейти обратно: а б Фишер, РА ; Балмуканд, Б. (июль 1928 г.). «Оценка сцепления потомства самоопыляющихся гетерозигот». Журнал генетики . 20 (1): 79–92. дои : 10.1007/BF02983317 . S2CID 27688031 .
- ^ Мадер, Сильвия (2007). Биология девятое издание . Нью-Йорк: МакГроу-Хилл. п. 209. ИСБН 978-0-07-325839-3 .
- ^ Гриффитс, AJF (2000). Введение в генетический анализ (7-е изд.). У. Х. Фриман.
- ^ Jump up to: Перейти обратно: а б Кантор, Рита М. (2013), «Анализ генетического сцепления», Римоин, Дэвид; Пьериц, Рид; Корф, Брюс (ред.), Принципы и практика медицинской генетики Эмери и Римуана (6-е изд.), Academic Press, стр. 1–9, doi : 10.1016/b978-0-12-383834-6.00010-0 , ISBN 9780123838346
- ^ Мортон Н.Е. (1955). «Последовательные тесты на выявление сцепления» . Американский журнал генетики человека . 7 (3): 277–318. ПМК 1716611 . ПМИД 13258560 .
- ^ Нихолт, Дейл Р. (август 2000 г.). «Все уровни детализации не созданы равными» . Американский журнал генетики человека . 67 (2): 282–288. дои : 10.1086/303029 . ПМЦ 1287176 . ПМИД 10884360 .
- ^ Риш, Нил (июнь 1991 г.). «Примечание о процедурах множественного тестирования в анализе связей» . Американский журнал генетики человека . 48 (6): 1058–1064. ПМК 1683115 . ПМИД 2035526 .
- ^ Феррейра, Мануэль А.Р. (1 октября 2004 г.). «Анализ сцепления: принципы и методы анализа количественных характеристик человека». Исследования близнецов и генетика человека . 7 (5): 513–530. дои : 10.1375/твин.7.5.513 . ISSN 2053-6003 . ПМИД 15527667 . S2CID 199001341 .
- ^ Гуселла, Джеймс Ф.; Фронтали, Марина; Васмут, Джон Дж.; Коллинз, Фрэнсис С.; Лерах, Ганс; Майерс, Ричард; Альтерр, Майкл; Аллитто, Бернис; Тейлор, Шерри (1 мая 1992 г.). «Регион-кандидат болезни Хантингтона демонстрирует множество различных гаплотипов». Природная генетика . 1 (2): 99–103. дои : 10.1038/ng0592-99 . ISSN 1546-1718 . ПМИД 1302016 . S2CID 25472459 .
- ^ Марк Дж. Дейли; Хиршхорн, Джоэл Н. (1 февраля 2005 г.). «Полногеномные исследования ассоциаций распространенных заболеваний и сложных признаков». Обзоры природы Генетика . 6 (2): 95–108. дои : 10.1038/nrg1521 . ISSN 1471-0064 . ПМИД 15716906 . S2CID 2813666 .
- ^ Паннетт, Колорадо; Бейтсон, В. (15 мая 1908 г.). «Наследственность пола» . Наука . 27 (698): 785–787. Бибкод : 1908Sci....27..785P . дои : 10.1126/science.27.698.785 . ISSN 0036-8075 . ПМИД 17791047 .
- ^ Гриффитс, AJF; Миллер, Дж. Х.; Сузуки, DT (2000). «Точный расчет больших расстояний по карте, рисунок 6-4» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman . ISBN 978-0-7167-3520-5 . График функции отображения по сравнению с идеализированным эквивалентом 1-1 процента частоты рекомбинации (RF%) для отображения единиц измерения.
- ^ Бензер С. Тонкая структура генетической области бактериофага. Proc Natl Acad Sci US A. 1955;41(6):344-354. doi:10.1073/pnas.41.6.344
- ^ Бензер С. О топологии тонкой генетической структуры. Proc Natl Acad Sci, США, 1959; 45 (11): 1607-1620. doi:10.1073/pnas.45.11.1607
- ^ Эдгар Р.С., Фейнман Р.П., Кляйн С., Лиелаусис I, Стейнберг CM. Эксперименты по картированию r-мутантов бактериофага T4D. Генетика. 1962;47:179–186. ПМК 1210321 . ПМИД 13889186
- ^ Фишер К.М., Бернштейн Х. Аддитивность интервалов в цистроне RIIA фага T4D. Генетика. 1965;52 (6):1127–1136. ПМК 1210971 . ПМИД 5882191
- ^ Макки, Брюс Д. (15 марта 2004 г.). «Гомологическое спаривание и динамика хромосом в мейозе и митозе» . Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1677 (1–3): 165–180. дои : 10.1016/j.bbaexp.2003.11.017 . ISSN 0006-3002 . ПМИД 15020057 .
- ^ Jump up to: Перейти обратно: а б Бернштейн Х. Влияние на рекомбинацию мутационных дефектов ДНК-полимеразы и дезоксицитидилатгидроксиметилазы фага T4D. Генетика. 1967;56(4):755-769
- ^ Jump up to: Перейти обратно: а б с д и Бергер Х., Уоррен А.Дж., Фрай К.Э. Вариации генетической рекомбинации из-за янтарных мутаций в бактериофаге T4D. Дж Вирол. 1969;3(2):171-175. doi:10.1128/JVI.3.2.171-175.1969
- ^ Бернштейн Х. О механизме внутригенной рекомбинации. I. Область rII бактериофага Т4. (1962) Журнал теоретической биологии. 1962 год; 3, 335–353. https://doi.org/10.1016/S0022-5193(62)80030-7
- ^ Jump up to: Перейти обратно: а б Бернштейн Х. Репарация и рекомбинация в фаге Т4. I. Гены, влияющие на рекомбинацию. Холодный источник Harb Symp Quant Biol. 1968;33:325-331. doi:10.1101/sqb.1968.033.01.037
- ^ Гамлетт Н.В., Бергер Х. Мутации, изменяющие генетическую рекомбинацию и восстановление ДНК в бактериофаге Т4. Вирусология. 1975;63(2):539-567. doi:10.1016/0042-6822(75)90326-8
- ^ Фудзисава Х., Йонесаки Т., Минагава Т. Последовательность гена рекомбинации Т4, uvsX, и ее сравнение с последовательностью гена RecA Escherichia coli. Нуклеиновые кислоты Рез. 1985;13(20):7473-7481. дои: 10.1093/нар/13.20.7473
- Гриффитс AJF; Миллер Дж. Х.; Сузуки ДТ; Левонтин РЦ; и др. (1993). «Глава 5» . Введение в генетический анализ (5-е изд.). Нью-Йорк: WH Freeman and Company. ISBN 978-0-7167-2285-4 .
- Полман Дж. М.; Слепер Д.А. (1995). «Глава 3». Селекция полевых культур (4-е изд.). Айова: Пресса штата Айова. ISBN 978-0-8138-2427-7 .
| Базы данных органов управления : Национальные |
|---|