Двухдоменная система
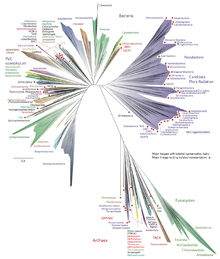
Двухдоменная система — это биологическая классификация , согласно которой все организмы на древе жизни делятся на два домена : бактерии и археи . [1] [2] [3] Оно возникло в результате развития знаний о разнообразии архей и бросило вызов широко принятой трехдоменной системе , которая классифицирует жизнь на бактерии, археи и эукарии . [4] Этому предшествовала гипотеза эоцитов Джеймса А. Лейка в 1980-х годах. [5] которая, согласно имеющимся на тот момент данным, в значительной степени была заменена трехдоменной системой. [6] Лучшее понимание архей, особенно их роли в происхождении эукариот посредством симбиогенеза с бактериями, привело к возрождению гипотезы эоцитов в 2000-х годах. [7] [8] Двухдоменная система получила более широкое распространение после открытия в 2017 году большой группы ( супертипа ) архей под названием Асгард . [9] данные свидетельствуют о том, что они являются эволюционным корнем эукариот, что делает эукариот членами домена Archaea. [10]
Хотя особенности архей Асгарда не исключают полностью трехдоменную систему, [11] [12] Представление о том, что эукариоты произошли от архей, было подтверждено генетическими и протеомными исследованиями. [13] В трехдоменной системе эукариоты в основном отличаются наличием «сигнатурных белков эукариот», которых нет у архей и бактерий. Однако археи Асгарда содержат гены, кодирующие несколько таких белков. [3]
Фон
[ редактировать ]Классификация жизни на два основных отдела не является новой концепцией: первое такое предложение было сделано французским биологом Эдуардом Шаттоном в 1938 году. Чаттон разделил организмы на:
- Прокариоты (включая бактерии)
- Эукариоты (включая простейших) [14]
Позже они были названы империями, а Чаттон классифицировал их как систему двух империй . [15] Чаттон использовал название «эукариоты» только для простейших, исключил других эукариот и опубликовал ограниченным тиражом, так что его работа не получила признания. Его классификация была заново открыта канадским бактериологом Роджером Йейтсом Станьером из Калифорнийского университета в Беркли в 1961 году в Институте Пастера в Париже. [14] В следующем году Станье и его коллега Корнелис Бернардус ван Ниль опубликовали в Archiv für Mikrobiologie (ныне Архивы микробиологии ) классификацию Чаттона с эукариотами, в которую были включены высшие водоросли, простейшие, грибы, растения и животные. [16] Она стала популярной системой классификации, как писал Джон О. Корлисс в 1986 году: «[Концепция Чаттона-Стэнье о царстве (точнее, суперцарстве) прокариот для бактерий (в самом широком смысле) и втором суперцарстве эукариот для всех остальных организмов был широко принят с энтузиазмом». [17]
В 1977 году Карл Вёзе и Джордж Э. Фокс классифицировали прокариоты на две группы (царства): архебактерии (от метаногенов , первые известные археи) и эубактерии, на основании их 16S рибосомальной РНК (16S рРНК). генов [18] В 1984 году Джеймс А. Лейк , Майкл Кларк, Эрик Хендерсон и Мелани Оукс из Калифорнийского университета в Лос-Анджелесе описали то, что было известно как «группа серозависимых бактерий», как новую группу организмов, называемую эоцитами (эоцитами). для «утренних клеток») и создал новое царство Эоцита. С его помощью они предположили существование четырех царств, основанных на строении и составе субъединиц рибосом, а именно архебактерий, эубактерий, эукариот и эоцитов. [19] Далее Лейк проанализировал последовательности рРНК четырех групп и предположил, что эукариоты произошли от эоцитов, а не от архебактерий, как обычно предполагалось. [20] Это было основой эоцитарной гипотезы . [6] В 1988 году он предложил разделить все формы жизни на две таксономические группы: [5]
- Кариоты (включая эукариоты и протоэукариотические организмы, такие как эоциты)
- Паркариоты (состоящие из эубактерий и архей, таких как галобактерии и метаногены). [21]
В 1990 году Вёзе, Отто Кандлер и Марк Уилис показали, что археи представляют собой отдельную группу организмов и что эоциты (переименованные в Crenarchaeota как тип архей [22] но исправлено как Thermoproteota в 2021 году. [23] ) являются археями. Они ввели основное разделение жизни на трехдоменную систему, включающую домен Эукария, домен Бактерии и домен Археи. [24] Благодаря ряду пересмотров деталей и открытию нескольких новых линий архей классификация Вёзе постепенно получила признание как «возможно, наиболее разработанная и наиболее широко принятая научная гипотеза [с классификацией пяти царств] относительно эволюционной истории жизни». [25]
Однако концепция трех доменов не решила проблем взаимоотношений между архей и эукариотами. [12] [26] Как выразился в 2020 году Форд Дулиттл , работавший тогда в Университете Далхаузи: «[Трёхдоменное дерево] неправильно представляет эволюционные взаимоотношения, представляя ошибочное представление о том, как эукариоты произошли от прокариотов. Трехдоменное дерево действительно распознаёт конкретную архейную… родство с эукариотами, но последнее возникнет независимо, а не изнутри, первое». [4]
Концепция
[ редактировать ]Двухдоменная система опирается главным образом на две ключевые концепции, которые определяют эукариотов как членов домена Archaea, а не как отдельный домен: эукариоты произошли от Archaea, а Asgards представляют собой происхождение эукариотов. [27] [28]
Эукариоты произошли от архей
[ редактировать ]Трехдоменная система предполагает, что эукариоты более тесно связаны с архей, чем с бактериями, и являются сестринской группой архей, поэтому она рассматривает их как отдельный домен. [29] По мере того, как в начале 2000-х годов было обнаружено больше новых архей, это различие стало сомнительным, поскольку эукариоты стали глубоко гнездиться внутри архей. Происхождение эукариот от архей, то есть они принадлежат к одной и той же большой группе, было подтверждено исследованиями, основанными на секвенировании рибосомных белков и филогенетическом анализе в 2004 году. [30] [31] Филогеномный анализ около 6000 наборов генов из 185 геномов бактерий, архей и эукариот в 2007 году также предположил происхождение эукариот от Euryarchaeota (в частности, Thermoplasmatales ). [32]
В 2008 году исследователи из Музея естественной истории в Лондоне и Университета Ньюкасла сообщили о комплексном анализе 53 генов архей, бактерий и эукариот, которые включали важные компоненты механизмов репликации, транскрипции и трансляции нуклеиновых кислот. Был сделан вывод, что эукариоты произошли от архей, в частности от Crenarchaeota (эоцитов), и результаты «в пользу топологии, которая поддерживает гипотезу эоцитов, а не архебактериальную монофилию и трехдоменное древо жизни». [26] Исследование, проведенное примерно в то же время, также обнаружило несколько генов, общих для эукариот и кренархеот. [33] Эти накапливающиеся доказательства подтверждают двухдоменную систему. [22]
В 2019 году исследование, проведенное доцентом ELTE Гергели Й. Сёллёси, также пришло к выводу, что правильной системой являются два домена. В проведенных исследованиях использовалось моделирование более 3000 семейств генов. Исследование пришло к выводу, что эукариоты , вероятно, произошли от бактерии, проникшей в хозяина Асгарда (вероятно, из типа Heimdallarchaeota ). [34] [35] [36]
Одним из отличий домена Eukarya в трехдоменной системе является то, что эукариоты обладают уникальными белками, такими как актин ( цитоскелета, микрофиламент участвующий в подвижности клеток), тубулин (компонент большого цитоскелета, микротрубочек ) и убиквитиновая система (деградация белков и переработка), которые не встречаются у прокариот. Однако эти так называемые «эукариотические сигнатурные белки» [3] кодируются в геномах архей TACK (включая типы Thaumarchaeota , Aigarchaeota , Crenarchaeota и Korarchaeota ), но не кодируются в геномах других архей. [22] Первыми эукариотическими белками, идентифицированными у Crenarchaeota, были актин и родственные актину белки (Arp) 2 и 3, что, возможно, объясняет происхождение эукариот симбиогенным фагоцитозом , при котором древний архейный хозяин имел основанный на актине механизм, с помощью которого он охватывал другие клетки, такие как протомитохондриальные бактерии. [37]
Тубулиноподобные белки, называемые артубулинами, обнаружены в геномах некоторых окисляющих аммоний Thaumarchaeota. [38] Эндосомальные сортировочные комплексы, необходимые для транспорта ( ESCRT III), участвующие в делении эукариотических клеток, обнаружены во всех группах TACK. [39] ESCRT-III-подобные белки составляют первичную систему деления клеток у этих архей. [40] [41] Гены, кодирующие систему убиквитина, известны из множества геномов Aigarchaeota. [42] Белок, родственный убиквитину, под названием Urm1, также присутствует у Crenarchaeota. [43] Система репликации ДНК (белки GINS) у Crenarchaeota и Halobacteria аналогична комплексу CMG (CDC45, MCM, GINS) эукариот. [44] Наличие этих эукариотических белков у архей указывает на их прямое родство и на то, что эукариоты произошли от архей. [22] [45]
Асгарды — последний общий предок эукариот.
[ редактировать ]Открытие Асгарда, описанного как «эукариотоподобные археи». [46] в 2012 году [47] [48] и следующий филогенетический анализ укрепил двухдоменный взгляд на жизнь. [49] Асгардские археи, называемые Lokiarchaeota, содержат даже больше эукариотических белковых генов, чем группа TACK. Первоначальный генетический анализ и последующий повторный анализ показали, что из более чем 31 выбранного эукариотического гена архей 75% из них напрямую поддерживают группировку эукариот-архей, то есть единый домен архей, включая эукариоты; [50] [51] хотя результаты не полностью исключили трехдоменную систему. [52]
Поскольку впоследствии было обнаружено больше групп Асгарда, включая Thorarchaeota , Odinarchaeota и Heimdallarchaeota, их отношения с эукариотами стали более устойчивыми. Филогенетический анализ с использованием генов рибосомальной РНК показал, что эукариоты произошли от Асгардов и что Heimdallarchaeota являются ближайшими родственниками эукариот. [9] [53] Происхождение эукариот от Heimdallarchaeota также подтверждено филогеномными исследованиями, проведенными в 2020 году. [13] Новая группа Асгардов, обнаруженная в 2021 году (предварительно названная Wukongarchaeota), также указывает на глубокие корни эукариотического происхождения. [54] В отчете 2022 года о другом Асгарде, названном Ньордархеота, указывается, что ветвь Хеймдаллархеота-Вуконгархеота, возможно, является исходной группой эукариот. [55]
Асгарды содержат по меньшей мере 80 генов сигнатурных белков эукариот. [56] Помимо актина, тубулина, убиквитина и белков ESCRT, обнаруженных у архей TACK, Асгарды содержат функциональные гены нескольких других эукариотических белков, таких как профилины . [57] убиквитиновая система (белки E1-подобный, E2-подобный и Srfp), [58] системы мембранного транспорта (такие как домены Sec23/24 и TRAPP), множество небольших GTPases [49] (включая ортологи ГТФазы семейства Gtr/Rag [59] ) и гельзолины . [60] Хотя эта информация не разрешает полностью противоречия между тремя и двумя доменами, [46] Обычно считается, что они отдают предпочтение двухдоменной системе. [3] [13] [61]
Классификация
[ редактировать ]Двухдоменная система определяет классификацию всех известных клеточных форм жизни на два домена: бактерии и археи. Он отменяет домен Eukaryota, признанный в трехдоменной классификации одним из основных доменов. В отличие от гипотезы эоцитов, которая предлагала две основные группы жизни (похожие на домены) и утверждала, что архей можно разделить как на бактериальные, так и на эукариотические группы, она объединила архей и эукариотов в один домен, а бактерии полностью в отдельный домен. [4]
Доменные бактерии
[ редактировать ]Он состоит из всех бактерий, которые являются прокариотами (без ядра), поэтому доменные бактерии состоят исключительно из прокариотических организмов. [62] [63] Некоторые примеры:
- Цианобактерии – фотосинтезирующие бактерии, родственные пластидам эукариот. [64]
- Spirochaetota – грамотрицательные бактерии, вызывающие такие заболевания человека, как сифилис и болезнь Лайма. [65]
- Actinomycetota – грамположительные бактерии, включая виды Streptomyces , из которых получены несколько антибиотиков, включая стрептомицин , неомицин , боттромицин и хлорамфеникол . [66] [67]
Домен Археи
[ редактировать ]В его состав входят как прокариотические, так и эукариотические организмы. [68]
Архея
Археи - это прокариотические организмы, некоторые примеры:
- Все метаногены – которые производят газ метан .
- Большинство галофилов – которые живут в очень соленой воде.
- Большинство термоацидофилов – обитают в кислой воде высокой температуры.
Эукария
Эукариоты имеют ядро в своих клетках и включают:
- Протисты – многие одноклеточные эукариоты, в том числе малярийные паразиты, амебы и диатомовые водоросли. [69]
- Царство грибов – эукариоты, такие как грибы, дрожжи и плесень.
- Королевство Plantae – все растения.
- Королевство Animalia – все животные.
Ссылки
[ редактировать ]- ^ Большой, Александр; Волкович, Зеев (Владимир); Киржнер, Валерий; Барзили, Зеев (2010), «Биологическая классификация» , Кластеризация генома , Исследования в области вычислительного интеллекта, том. 286, Берлин, Гейдельберг: Springer Berlin Heidelberg, стр. 17–22, doi : 10.1007/978-3-642-12952-0_2 , ISBN 978-3-642-12951-3 , получено 14 мая 2022 г.
- ^ Рэйманн, Кэйси; Брошье-Армане, Селин; Грибальдо, Симонетта (2015). «Двудоменное древо жизни связано с новым корнем архей» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (21): 6670–6675. Бибкод : 2015PNAS..112.6670R . дои : 10.1073/pnas.1420858112 . ПМК 4450401 . ПМИД 25964353 .
- ^ Jump up to: а б с д Нобс, Стефани-Джейн; Маклауд, Фрейзер И.; Вонг, Хон Лун; Бернс, Брендан П. (2022). «Эукария-химера: эукариоты, вторичное нововведение двух областей жизни?» . Тенденции в микробиологии . 30 (5): 421–431. дои : 10.1016/j.tim.2021.11.003 . ПМИД 34863611 . S2CID 244823103 .
- ^ Jump up to: а б с Дулитл, У. Форд (2020). «Эволюция: две области жизни или три?» . Современная биология . 30 (4): Р177–Р179. дои : 10.1016/j.cub.2020.01.010 . ПМИД 32097647 .
- ^ Jump up to: а б Лейк, Джеймс А. (1988). «Происхождение эукариотического ядра определяется с помощью скоростно-инвариантного анализа последовательностей рРНК». Природа . 331 (6152): 184–186. Бибкод : 1988Natur.331..184L . дои : 10.1038/331184a0 . ПМИД 3340165 . S2CID 4368082 .
- ^ Jump up to: а б Арчибальд, Джон М. (2008). «Эоцитарная гипотеза и происхождение эукариотических клеток» . Труды Национальной академии наук . 105 (51): 20049–20050. Бибкод : 2008PNAS..10520049A . дои : 10.1073/pnas.0811118106 . ПМЦ 2629348 . ПМИД 19091952 .
- ^ Пул, Энтони М.; Пенни, Дэвид (2007). «Оценка гипотез происхождения эукариот» . Биоэссе . 29 (1): 74–84. doi : 10.1002/bies.20516 . ПМИД 17187354 .
- ^ Фостер, Питер Г.; Кокс, Саймон Дж.; Эмбли, Т. Мартин (2009). «Основные подразделения жизни: филогеномный подход с использованием методов гетерогенного состава» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 364 (1527): 2197–2207. дои : 10.1098/rstb.2009.0034 . ПМК 2873002 . ПМИД 19571240 .
- ^ Jump up to: а б Заремба-Недзведска, Катажина; Касерес, Ева Ф.; Пила, Джимми Х.; Бэкстрем, Диса; Юзокайте, Лина; Ванкастер, Эммельен; Зейтц, Кили В.; Анантараман, Картик; Старнавский, Петр (2017). «Археи Асгарда освещают происхождение сложности эукариотических клеток» (PDF) . Природа . 541 (7637): 353–358. Бибкод : 2017Natur.541..353Z . дои : 10.1038/nature21031 . ОСТИ 1580084 . ПМИД 28077874 . S2CID 4458094 .
- ^ Эме, Лаура; Спанг, Аня; Ломбард, Джонатан; Лестница, Кортни В.; Эттема, Тийс Дж.Г. (10 ноября 2017 г.). «Археи и происхождение эукариот» . Обзоры природы Микробиология . 15 (12): 711–723. дои : 10.1038/nrmicro.2017.133 . ISSN 1740-1534 . ПМИД 29123225 . S2CID 8666687 .
- ^ Да Кунья, Виолетт; Гайя, Морган; Насир, Аршан; Фортерре, Патрик (2018). «Асгардские археи не закрывают дискуссию о топологии универсального древа жизни» . ПЛОС Генетика . 14 (3): e1007215. дои : 10.1371/journal.pgen.1007215 . ПМЦ 5875737 . ПМИД 29596428 .
- ^ Jump up to: а б Чжоу, Чжичао; Лю, Ян; Ли, Мэн; Гу, Цзи-Донг (2018). «Два или три домена: новый взгляд на древо жизни в эпоху геномики» . Прикладная микробиология и биотехнология . 102 (7): 3049–3058. дои : 10.1007/s00253-018-8831-x . ПМИД 29484479 . S2CID 3541409 .
- ^ Jump up to: а б с Уильямс, Том А.; Кокс, Саймон Дж.; Фостер, Питер Г.; Сёллёси, Гергеи Дж.; Эмбли, Т. Мартин (2020). «Филогеномика обеспечивает надежную поддержку двухдоменного древа жизни» . Экология и эволюция природы . 4 (1): 138–147. дои : 10.1038/s41559-019-1040-x . ПМК 6942926 . ПМИД 31819234 .
- ^ Jump up to: а б Качер, Фридрих (2004). «История терминов прокариоты и эукариоты» . Протист . 155 (2): 257–263. дои : 10.1078/143446104774199637 . ПМИД 15305800 .
- ^ Майр, Эрнст (1998). «Две империи или три?» . Труды Национальной академии наук . 95 (17): 9720–9723. Бибкод : 1998PNAS...95.9720M . дои : 10.1073/pnas.95.17.9720 . ПМК 33883 . ПМИД 9707542 .
- ^ Станье, Р.Ю.; Ван Нил, CB (1962). «Концепция бактерии» (PDF) . Архив микробиологии . 42 :17-35. дои : 10.1007/BF00425185 . ПМИД 13916221 . S2CID 29859498 .
- ^ Корлисс, Джон О. (1986). «Царства организмов: с точки зрения микроскописта» . Труды Американского микроскопического общества . 105 (1): 1–10. дои : 10.2307/3226544 . JSTOR 3226544 .
- ^ Woese CR, Fox GE (ноябрь 1977 г.). «Филогенетическая структура прокариотического домена: первичные царства» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (11): 5088–90. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ПМК 432104 . ПМИД 270744 .
- ^ Лейк, Джеймс А.; Хендерсон, Эрик; Оукс, Мелани; Кларк, Майкл В. (июнь 1984 г.). «Эоциты: новая структура рибосом указывает на царство, имеющее близкое родство с эукариотами» . ПНАС . 81 (12): 3786–3790. Бибкод : 1984PNAS...81.3786L . дои : 10.1073/pnas.81.12.3786 . ПМК 345305 . ПМИД 6587394 .
- ^ Лейк, Дж.А. (1987). «Прокариоты и архебактерии не монофилетичны: инвариантный анализ генов рРНК показывает, что эукариоты и эоциты образуют монофилетический таксон» . Симпозиумы Колд-Спринг-Харбор по количественной биологии . 52 : 839–846. дои : 10.1101/SQB.1987.052.01.091 . ISSN 0091-7451 . ПМИД 3454292 .
- ^ Лейк, Джеймс А. (1991). «Отслеживание происхождения с помощью молекулярных последовательностей: многоклеточные и эукариотические начала» . Тенденции биохимических наук . 16 (2): 46–50. дои : 10.1016/0968-0004(91)90020-В . ПМИД 1858129 .
- ^ Jump up to: а б с д Кунин, Евгений В. (2015). «Происхождение эукариот из архей, архейных эукариом и всплески увеличения генов: эукариогенез стал проще?» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1678): 20140333. doi : 10.1098/rstb.2014.0333 . ПМЦ 4571572 . ПМИД 26323764 .
- ^ Орен, Аарон; Гаррити, Джордж М. (2021). «Действительная публикация названий сорока двух типов прокариот» . Международный журнал систематической и эволюционной микробиологии . 71 (10): Онлайн. дои : 10.1099/ijsem.0.005056 . ISSN 1466-5034 . ПМИД 34694987 . S2CID 239887308 .
- ^ Вёзе Ч.Р., Кандлер О., Уилис М.Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–9. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Кейс, Эмили (2008). «Преподавание таксономии: сколько королевств?» . Американский учитель биологии . 70 (8): 472–477. doi : 10.1662/0002-7685(2008)70[472:TTHMK]2.0.CO;2 . S2CID 198969439 .
- ^ Jump up to: а б Кокс, Саймон Дж.; Фостер, Питер Г.; Хирт, Роберт П.; Харрис, Саймон Р.; Эмбли, Т. Мартин (2008). «Архебактериальное происхождение эукариот» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (51): 20356–20361. Бибкод : 2008PNAS..10520356C . дои : 10.1073/pnas.0810647105 . ПМЦ 2629343 . ПМИД 19073919 .
- ^ Маклауд, Фрейзер; Киндлер, Гарет С.; Вонг, Хон Лун; Чен, Рэй; Бернс, Брендан П. (2019). «Асгардские археи: разнообразие, функции и эволюционные последствия в ряде микробиомов» . АИМС Микробиология . 5 (1): 48–61. дои : 10.3934/микробиол.2019.1.48 . ПМК 6646929 . ПМИД 31384702 .
- ^ Юттнер, Майкл; Феррейра-Серка, Себастьен (2022). «Взгляд сквозь призму эволюционной истории биогенеза рибосом: возможные последствия для филогении архей и эукариогенеза» . Молекулярная биология и эволюция . 39 (4): msac054. дои : 10.1093/molbev/msac054 . ПМЦ 8997704 . ПМИД 35275997 .
- ^ Эме, Лаура; Спанг, Аня; Ломбард, Джонатан; Лестница, Кортни В.; Эттема, Тийс Дж.Г. (2017). «Археи и происхождение эукариот» . Обзоры природы Микробиология . 15 (12): 711–723. дои : 10.1038/nrmicro.2017.133 . ПМИД 29123225 . S2CID 8666687 .
- ^ Вишванат, Прашант; Фаваретто, Паола; Хартман, Хайман; Мор, Скотт С.; Смит, Темпл Ф. (2004). «Структура блоков рибосомальных белковых последовательностей предполагает сложную эволюцию прокариот, что имеет значение для происхождения эукариот» . Молекулярная филогенетика и эволюция . 33 (3): 615–625. дои : 10.1016/j.ympev.2004.07.003 . ПМИД 15522791 .
- ^ Ривера, Мария К.; Лейк, Джеймс А. (2004). «Кольцо жизни является доказательством происхождения эукариот в результате слияния геномов» . Природа . 431 (7005): 152–155. Бибкод : 2004Natur.431..152R . дои : 10.1038/nature02848 . ПМИД 15356622 . S2CID 4349149 .
- ^ Пизани, Давиде; Коттон, Джеймс А.; Макинерни, Джеймс О. (2007). «Супердеревья раскрывают химерическое происхождение геномов эукариот» . Молекулярная биология и эволюция . 24 (8): 1752–1760. дои : 10.1093/molbev/msm095 . ПМИД 17504772 .
- ^ Ютин, Наталья; Макарова Кира С.; Мехедов Сергей Л.; Вольф, Юрий И.; Кунин, Евгений В. (2008). «Глубокие архейные корни эукариотов» . Молекулярная биология и эволюция . 25 (8): 1619–1630. дои : 10.1093/molbev/msn108 . ПМЦ 2464739 . ПМИД 18463089 .
- ^ Уильямс, Том А.; Кокс, Саймон Дж.; Фостер, Питер Г.; Сёллёси, Гергеи Дж.; Эмбли, Т. Мартин (январь 2020 г.). «Филогеномика обеспечивает надежную поддержку двухдоменного древа жизни» . Экология и эволюция природы . 4 (1): 138–147. дои : 10.1038/s41559-019-1040-x . ISSN 2397-334X . ПМК 6942926 .
- ^ «Империи сокращают расходы: совместный успех ELTE и Бристольского университета в исследовании наших самых ранних предков» . MTA.hu (на венгерском языке). 10 декабря 2019 г. Проверено 25 июля 2024 г.
- ^ Чаба, Мольнар (13 декабря 2019 г.). «Венгерский исследователь переписывает историю эволюции» . index.hu (на венгерском языке) . Проверено 25 июля 2024 г.
- ^ Ютин, Наталья; Вольф, Максим Юрьевич; Вольф, Юрий И.; Кунин, Евгений В. (2009). «Происхождение фагоцитоза и эукариогенеза» . Биология Директ . 4 :9. дои : 10.1186/1745-6150-4-9 . ПМК 2651865 . ПМИД 19245710 .
- ^ Ютин, Наталья; Кунин, Евгений В. (2012). «Археальное происхождение тубулина» . Биология Директ . 7:10 . дои : 10.1186/1745-6150-7-10 . ПМЦ 3349469 . ПМИД 22458654 .
- ^ Самсон, Рэйчел Ю.; Добро, Меган Дж.; Дженсен, Грант Дж.; Белл, Стивен Д. (2017). «Структура, функции и роли архейного аппарата ESCRT» . Прокариотические цитоскелеты . Субклеточная биохимия. Том. 84. С. 357–377. дои : 10.1007/978-3-319-53047-5_12 . ISBN 978-3-319-53045-1 . ПМИД 28500532 .
- ^ Пельве, Эрик А.; Линдос, Анн-Кристин; Мартенс-Хаббена, Вильм; из Тауэра, Джозеф Р.; Шталь, Дэвид А.; Бернандер, Рольф (2011). «Деление клеток на основе Cdv и организация клеточного цикла у таумаркеона Nitrosopumilus maritimus» . Молекулярная микробиология . 82 (3): 555–566. дои : 10.1111/j.1365-2958.2011.07834.x . ПМИД 21923770 . S2CID 1202516 .
- ^ Добро, Меган Дж.; Самсон, Рэйчел Ю.; Ю, Чжихэн; Маккалоу, Джон; Дин, Х. Джейн; Чонг, Парксон Ли-Гау; Белл, Стивен Д.; Дженсен, Грант Дж. (2013). «Электронная криотомография сборок ESCRT и делящихся клеток Sulfolobus предполагает, что спиралевидные нити участвуют в разрыве мембраны» . Молекулярная биология клетки . 24 (15): 2319–2327. дои : 10.1091/mbc.E12-11-0785 . ПМЦ 3727925 . ПМИД 23761076 .
- ^ Грау-Бове, Ксавье; Себе-Педрос, Арнау; Руис-Трилло, Иньяки (2015). «Предок эукариот имел сложную убиквитиновую сигнальную систему архейного происхождения» . Молекулярная биология и эволюция . 32 (3): 726–739. дои : 10.1093/molbev/msu334 . ПМЦ 4327156 . ПМИД 25525215 .
- ^ Макарова Кира С.; Кунин, Евгений В. (2010). «Архейные убиквитиноподобные белки: функциональная универсальность и предполагаемое участие предков в модификации тРНК, выявленное с помощью сравнительного геномного анализа» . Архея . 2010 : 710303. doi : 10.1155/2010/710303 . ПМЦ 2948915 . ПМИД 20936112 .
- ^ Макарова Кира С.; Кунин Евгений Владимирович; Кельман, Цви (2012). «Комплекс CMG (CDC45/RecJ, MCM, GINS) является консервативным компонентом системы репликации ДНК у всех архей и эукариот» . Биология Директ . 7 :7. дои : 10.1186/1745-6150-7-7 . ПМЦ 3307487 . ПМИД 22329974 .
- ^ Лестница, Кортни В.; Эттема, Тейс Дж.Г. (2020). «Архейные корни эукариотического динамического актинового цитоскелета» . Современная биология . 30 (10): Р521–Р526. дои : 10.1016/j.cub.2020.02.074 . ПМИД 32428493 . S2CID 218710903 .
- ^ Jump up to: а б Фурнье, Грегори П.; Пул, Энтони М. (2018). «Кратко аргументированный случай, что асгардские археи являются частью дерева эукариот» . Границы микробиологии . 9 : 1896. doi : 10.3389/fmicb.2018.01896 . ПМК 6104171 . ПМИД 30158917 .
- ^ Йоргенсен, Штеффен Лет; Ханнисдал, Бьярте; Ланцен, Андерс; Баумбергер, Тамара; Флесланд, Кристин; Фонсека, Рита; Овреас, Лиза; Стин, Ида Х; Торсет, Ингунн Х; Педерсен, Рольф Б; Шлепер, Криста (5 сентября 2012 г.). «Корреляция профилей микробных сообществ с геохимическими данными в сильно стратифицированных отложениях Арктического Срединно-океанического хребта» . ПНАС . 109 (42): E2846–55. дои : 10.1073/pnas.1207574109 . ПМК 3479504 . ПМИД 23027979 .
- ^ Йоргенсен, Штеффен Лет; Торсет, Ингунн Х; Педерсен, Рольф Б; Баумбергер, Тамара; Шлепер, Криста (4 октября 2013 г.). «Количественное и филогенетическое изучение группы глубоководных архей в отложениях Арктического срединно-океанического спредингового хребта» . Границы микробиологии . 4 : 299. дои : 10.3389/fmicb.2013.00299 . ПМК 3790079 . ПМИД 24109477 .
- ^ Jump up to: а б Спанг, Аня; Пила, Джимми Х.; Йоргенсен, Штеффен Л.; Заремба-Недзведска, Катажина; Мартейн, Йоран; Линд, Андерс Э.; Эйк, Роэл ван; Шлепер, Криста; Гай, Лайонел (2015). «Сложные археи, заполняющие пропасть между прокариотами и эукариотами» . Природа . 521 (7551): 173–179. Бибкод : 2015Natur.521..173S . дои : 10.1038/nature14447 . ПМЦ 4444528 . ПМИД 25945739 .
- ^ Да Кунья, Виолетт; Гайя, Морган; Гадель, Даниэле; Насир, Аршан; Фортерре, Патрик (2017). «Локиархеи являются близкими родственниками Euryarchaeota, не заполняя пропасть между прокариотами и эукариотами» . ПЛОС Генетика . 13 (6): e1006810. дои : 10.1371/journal.pgen.1006810 . ПМЦ 5484517 . ПМИД 28604769 .
- ^ Да Кунья, Виолетт; Гайя, Морган; Фортерре, Патрик (2022). «Расширяющиеся археи Асгарда и их неуловимые отношения с эукарией» . mLife . 1 (1): 3–12. дои : 10.1002/mlf2.12012 . S2CID 247689333 .
- ^ Да Кунья, Виолетт; Гайя, Морган; Насир, Аршан; Фортерре, Патрик (2018). «Асгардские археи не закрывают дискуссию о топологии универсального древа жизни» . ПЛОС Генетика . 14 (3): e1007215. дои : 10.1371/journal.pgen.1007215 . ISSN 1553-7404 . ПМЦ 5875737 . ПМИД 29596428 .
- ^ Спанг, Аня; Эме, Лаура; Пила, Джимми Х.; Касерес, Ева Ф.; Заремба-Недзведска, Катажина; Ломбард, Джонатан; Гай, Лайонел; Эттема, Тейс Дж.Г. (2018). «Асгардские археи — ближайшие прокариотические родственники эукариотов» . ПЛОС Генетика . 14 (3): e1007080. дои : 10.1371/journal.pgen.1007080 . ПМЦ 5875740 . ПМИД 29596421 .
- ^ Лю, Ян; Макарова Кира С.; Хуан, Вэнь-Цун; Вольф, Юрий И.; Никольская Анастасия Н.; Чжан, Синьсюй; Цай, Минвэй; Чжан, Цуй-Цзин; Сюй, Вэй; Ло, Чжухуа; Ченг, Лей (2021). «Расширенное разнообразие архей Асгарда и их взаимоотношения с эукариотами» . Природа . 593 (7860): 553–557. Бибкод : 2021Natur.593..553L . дои : 10.1038/s41586-021-03494-3 . ISSN 0028-0836 . ПМЦ 11165668 . ПМИД 33911286 . S2CID 233447651 .
- ^ Се, Руизе; Ван, Иньчжао; Хуан, Даньюэ; Хоу, Цзялин; Ли, Люян; Ху, Хайнин; Чжао, Сяосяо; Ван, Фэнпин (2022). «Распространение членов Асгарда во владениях архей проливает новый свет на происхождение эукариотов» . Наука Китай Науки о жизни . 65 (4): 818–829. дои : 10.1007/s11427-021-1969-6 . ПМИД 34378142 . S2CID 236977853 .
- ^ Лопес-Гарсия, Purificación; Морейра, Дэвид (2020). «Культивированные асгардские археи проливают свет на эукариогенез» . Клетка . 181 (2): 232–235. дои : 10.1016/j.cell.2020.03.058 . ПМИД 32302567 . S2CID 215798394 .
- ^ Акыл, Джанер; Робинсон, Роберт С. (2018). «Геномы архей Асгарда кодируют профилины, регулирующие актин» . Природа . 562 (7727): 439–443. Бибкод : 2018Natur.562..439A . дои : 10.1038/s41586-018-0548-6 . ПМИД 30283132 . S2CID 52917038 .
- ^ Хеннелл Джеймс, Рори; Касерес, Ева Ф.; Эскасинас, Алекс; Альхасан, Хая; Ховард, Джули А.; Дири, Майкл Дж.; Эттема, Тейс Дж.Г.; Робинсон, Николас П. (2017). «Функциональная реконструкция эукариотоподобного каскада убиквитилирования E1/E2/(RING) E3 из некультивируемого архея» . Природные коммуникации . 8 (1): 1120. Бибкод : 2017NatCo...8.1120H . дои : 10.1038/s41467-017-01162-7 . ISSN 2041-1723 . ПМЦ 5654768 . ПМИД 29066714 .
- ^ Клингер, Кристен М.; Спанг, Аня; Дакс, Джоэл Б.; Эттема, Тейс Дж.Г. (2016). «Отслеживание архейного происхождения строительных блоков эукариотической системы мембранного транспорта» . Молекулярная биология и эволюция . 33 (6): 1528–1541. дои : 10.1093/molbev/msw034 . ПМИД 26893300 .
- ^ Акыл, Джанер; Тран, Линь Т.; Оран-Приу, Магали; Баскаран, Йохендран; Мансер, Эдвард; Бланшуан, Лоран; Робинсон, Роберт С. (2020). «Понимание эволюции регулируемой динамики актина посредством характеристики примитивных белков гельзолина / кофилина из архей Асгарда» . Труды Национальной академии наук . 117 (33): 19904–19913. Бибкод : 2020PNAS..11719904A . дои : 10.1073/pnas.2009167117 . ПМЦ 7444086 . ПМИД 32747565 .
- ^ Лю, Ян; Макарова Кира С.; Хуан, Вэнь-Цун; Вольф, Юрий И.; Никольская Анастасия Н.; Чжан, Синьсюй; Цай, Минвэй; Чжан, Цуй-Цзин; Сюй, Вэй; Ло, Чжухуа; Ченг, Лей (2021). «Расширенное разнообразие архей Асгарда и их взаимоотношения с эукариотами» . Природа . 593 (7860): 553–557. Бибкод : 2021Natur.593..553L . дои : 10.1038/s41586-021-03494-3 . ПМЦ 11165668 . ПМИД 33911286 . S2CID 233447651 .
- ^ Гугенгольц, Филип (2002). «Изучение прокариотического разнообразия в эпоху генома» . Геномная биология . 3 (2): обзоры0003.1. doi : 10.1186/gb-2002-3-2-reviews0003 . ПМК 139013 . ПМИД 11864374 .
- ^ Орен, Аарон (2004). Годфрей, HCJ; Кнапп, С. (ред.). «Разнообразие и таксономия прокариот: современное состояние и будущие проблемы» . Философские труды Лондонского королевского общества. Серия Б: Биологические науки . 359 (1444): 623–638. дои : 10.1098/rstb.2003.1458 . ПМЦ 1693353 . ПМИД 15253349 .
- ^ Мур, Келси Р.; Магнабоско, Кара; Момпер, Лили; Голд, Дэвид А.; Босак, Таня; Фурнье, Грегори П. (2019). «Расширенная рибосомальная филогения цианобактерий поддерживает глубокое размещение пластид» . Границы микробиологии . 10 : 1612. дои : 10.3389/fmicb.2019.01612 . ISSN 1664-302X . ПМК 6640209 . ПМИД 31354692 .
- ^ Зейтц, Патрик; Блокеш, Мелани (2013). «Символы и регуляторные пути, участвующие в естественной компетентности и трансформации патогенных и экологических грамотрицательных бактерий» . Обзоры микробиологии FEMS . 37 (3): 336–363. дои : 10.1111/j.1574-6976.2012.00353.x . ПМИД 22928673 . S2CID 30861416 .
- ^ Бибб, Мервин Дж. (2013). «Понимание и управление выработкой антибиотиков актиномицетами» . Труды Биохимического общества . 41 (6): 1355–1364. дои : 10.1042/BST20130214 . ПМИД 24256223 .
- ^ Вудрафф, Х. Бойд (2014). «Селман А. Ваксман, лауреат Нобелевской премии 1952 года по физиологии и медицине» . Прикладная и экологическая микробиология . 80 (1): 2–8. Бибкод : 2014ApEnM..80....2W . дои : 10.1128/АЕМ.01143-13 . ПМК 3911012 . ПМИД 24162573 .
- ^ Шляйфер, Карл Хайнц (2009). «Классификация бактерий и архей: прошлое, настоящее и будущее» . Систематическая и прикладная микробиология . 32 (8): 533–542. дои : 10.1016/j.syapm.2009.09.002 . ПМИД 19819658 .
- ^ Адл, Сина М.; Симпсон, Аластер ГБ; Фармер, Марк А.; Андерсен, Роберт А.; Андерсон, О. Роджер; Барта, Джон Р.; Баузер, Сэмюэл С.; Брюгероль, Гай; Фенсом, Роберт А.; Фредерик, Сюзанна; Джеймс, Тимоти Ю. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . S2CID 8060916 .