Развитие нервной системы
| Эта статья является частью серии статей о |
| Развитие систем органов |
|---|
Развитие нервной системы , или нервное развитие ( нейроразвитие ), относится к процессам, которые порождают, формируют и изменяют нервную систему животных, начиная с самых ранних стадий эмбрионального развития и до взрослой жизни. Область развития нервной системы опирается как на нейробиологию, так и на биологию развития, чтобы описать и дать представление о клеточных и молекулярных механизмах, с помощью которых развиваются сложные нервные системы, от нематод и плодовых мух до млекопитающих .
Дефекты развития нервной системы могут привести к порокам развития, таким как голопрозэнцефалия , а также широкому спектру неврологических расстройств , включая парезы и параличи конечностей , нарушения равновесия и зрения, а также судороги . [1] и у людей другие расстройства, такие как синдром Ретта , синдром Дауна и умственная отсталость . [2]
Развитие мозга позвоночных
[ редактировать ]
Центральная (ЦНС) позвоночных нервная система происходит из эктодермы — самого внешнего зародышевого листка эмбриона. Часть дорсальной эктодермы специфицируется в нейральную эктодерму – нейроэктодерму , образующую нервную пластинку вдоль дорсальной стороны зародыша. [3] [4] Это часть раннего формирования паттерна эмбриона (включая эмбрионы беспозвоночных), которая также устанавливает передне-заднюю ось. [5] [6] Нервная пластинка является источником большинства нейронов и глиальных клеток ЦНС. Нервная борозда формируется вдоль длинной оси нервной пластинки, а нервная пластинка складывается, образуя нервную трубку . [7] Этот процесс известен как нейруляция . [8] Когда трубка закрыта с обоих концов, она заполняется эмбриональной спинномозговой жидкостью. [9] По мере развития эмбриона передняя часть нервной трубки расширяется и образует три первичных мозговых пузырька , которые становятся передним мозгом ( прозэнцефалон ), средним мозгом ( мезенцефалон ) и задним мозгом ( ромбэнцефалон ). Эти простые ранние пузырьки увеличиваются и далее делятся на конечный мозг (будущие кора головного мозга и базальные ганглии ), промежуточный мозг (будущий таламус и гипоталамус ), средний мозг (будущие холмики ), метэнцефалон (будущий мост и мозжечок ) и миеленцефалон (будущий мозговой мозг ). [10] Заполненная спинномозговой жидкостью центральная камера непрерывна от конечного мозга до центрального канала спинного мозга и представляет собой развивающуюся желудочковую систему ЦНС. Эмбриональная спинномозговая жидкость отличается от образующейся на более поздних стадиях развития и от спинномозговой жидкости взрослых; это влияет на поведение нейронных предшественников. [9] Поскольку нервная трубка дает начало головному и спинному мозгу, любые мутации на этой стадии развития могут привести к фатальным уродствам, таким как анэнцефалия , или пожизненным инвалидностям, таким как расщелина позвоночника . В это время стенки нервной трубки содержат нервные стволовые клетки , которые способствуют росту мозга, поскольку они многократно делятся. Постепенно некоторые клетки перестают делиться и дифференцируются в нейроны и глиальные клетки , которые являются основными клеточными компонентами ЦНС. [11] Вновь созданные нейроны мигрируют в разные части развивающегося мозга, чтобы самоорганизоваться в разные структуры мозга. Как только нейроны достигают своих региональных позиций, они расширяют аксоны и дендриты , что позволяет им общаться с другими нейронами через синапсы . Синаптическая связь между нейронами приводит к созданию функциональных нейронных цепей , которые опосредуют сенсорную и моторную обработку и лежат в основе поведения. [12]

Индукция
[ редактировать ]Во время раннего эмбрионального развития позвоночных дорсальная эктодерма становится специфической, дающей начало эпидермису и нервной системе; часть дорсальной эктодермы переходит в нервную эктодерму, образуя нервную пластинку , дающую начало нервной системе. [3] [13] Преобразование недифференцированной эктодермы в нейроэктодерму требует сигналов от мезодермы . В начале гаструляции презумптивные мезодермальные клетки перемещаются через дорсальную губу бластопора и образуют слой мезодермы между энтодермой и эктодермой. Мезодермальные клетки мигрируют вдоль дорсальной срединной линии, давая начало хорде , которая развивается в позвоночник . Нейроэктодерма, покрывающая хорду, развивается в нервную пластинку в ответ на диффузный сигнал, вырабатываемый хордой. Остальная часть эктодермы дает начало эпидермису. Способность мезодермы превращать вышележащую эктодерму в нервную ткань называется нейронной индукцией .
У раннего эмбриона нервная пластинка сгибается наружу, образуя нервную борозду . Начиная с области будущей шеи, нервные складки этой борозды смыкаются, образуя нервную трубку . Образование нервной трубки из эктодермы называется нейруляцией . Вентральная часть нервной трубки называется базальной пластинкой ; спинная часть называется крыловой пластинкой . Полая внутренняя часть называется нервным каналом , а открытые концы нервной трубки, называемые нейропорами, закрываются. [14]
Пересаженная губа бластопора может превращать эктодерму в нервную ткань и, как говорят, оказывает индуктивный эффект. Нейронные индукторы представляют собой молекулы, которые могут индуцировать экспрессию нервных генов в эксплантатах эктодермы, не индуцируя также мезодермальные гены. Нейральную индукцию часто изучают на эмбрионах Xenopus, поскольку они имеют простое строение тела и имеются хорошие маркеры, позволяющие различать нервную и ненервную ткани. Примерами нейронных индукторов являются молекулы noggin и chordin .
Когда эмбриональные эктодермальные клетки культивируются при низкой плотности в отсутствие мезодермальных клеток, они подвергаются нейральной дифференцировке (экспрессируют нервные гены), указывая тем самым, что нервная дифференцировка является судьбой эктодермальных клеток по умолчанию. В культурах эксплантатов (которые допускают прямое межклеточное взаимодействие) одни и те же клетки дифференцируются в эпидермис. Это связано с действием BMP4 ( белок семейства TGF-β ), который индуцирует дифференцировку эктодермальных культур в эпидермис. Во время нейронной индукции noggin и chordin продуцируются дорсальной мезодермой (нотохордой) и диффундируют в вышележащую эктодерму, ингибируя активность BMP4. Это ингибирование BMP4 заставляет клетки дифференцироваться в нервные клетки. Ингибирование передачи сигналов TGF-β и BMP (костного морфогенетического белка) может эффективно индуцировать нервную ткань из плюрипотентных стволовых клеток . [15]
Регионализация
[ редактировать ]На более поздней стадии развития верхняя часть нервной трубки сгибается на уровне будущего среднего мозга — среднего мозга , у мезэнцефалического изгиба или цефалического изгиба . Над средним мозгом находится прозэнцефалон (будущий передний мозг), а под ним — ромбэнцефалон (будущий задний мозг).
Крылья пластинка переднего мозга расширяется, образуя конечный мозг , дающий начало полушариям головного мозга , а его базальная пластинка становится промежуточным мозгом . Оптический пузырек (который в конечном итоге становится зрительным нервом, сетчаткой и радужной оболочкой) формируется в базальной пластинке переднего мозга.
Узоры
[ редактировать ]У хордовых дорсальная эктодерма образует всю нервную ткань и нервную систему. Формирование паттерна происходит из-за специфических условий окружающей среды — различной концентрации сигнальных молекул.
Дорсовентральная ось
[ редактировать ]Вентральная половина нервной пластинки контролируется хордой , которая действует как «организатор». Дорсальная половина контролируется пластинкой эктодермы , которая по бокам от нервной пластинки. [16]
Эктодерма следует стандартному пути, чтобы стать нервной тканью. Доказательством этого служат единичные культивированные клетки эктодермы, которые в дальнейшем формируют нервную ткань. Предполагается, что это связано с отсутствием BMP , которые заблокированы организатором. Организатор может продуцировать такие молекулы, как фоллистатин , ноггин и хордин, которые ингибируют BMP.
Вентральная нервная трубка имеет структуру звукового ежа (Shh) из хорды, которая действует как индуцирующая ткань. Полученный из нотохорды Shh передает сигнал пластинке дна и индуцирует экспрессию Shh в пластинке дна. Shh, происходящий из пластинки дна, впоследствии передает сигналы др. клеткам нервной трубки и важен для правильной спецификации доменов-предшественников вентральных нейронов. Потеря Shh из хорды и/или пластинки дна препятствует правильной спецификации этих доменов-предшественников. Shh связывает Patched1 , снимая опосредованное Patched ингибирование Smoothened , что приводит к активации семейства транскрипционных факторов Gli ( GLI1 , GLI2 и GLI3 ).
В этом контексте Shh действует как морфоген - он индуцирует дифференцировку клеток в зависимости от его концентрации. В низких концентрациях он образует вентральные интернейроны , в более высоких концентрациях он индуцирует развитие двигательных нейронов , а в самых высоких концентрациях он индуцирует дифференцировку пластинки дна. Нарушение Shh-модулированной дифференцировки вызывает голопрозэнцефалию .
Дорсальная нервная трубка образована BMP из эпидермальной эктодермы, фланкирующей нервную пластинку. Они индуцируют сенсорные интернейроны путем активации киназ Sr/Thr и изменения уровней транскрипционного фактора SMAD .
Рострокаудальная (переднезадняя) ось
[ редактировать ]Сигналы, которые контролируют переднезаднее развитие нейронов, включают FGF и ретиноевую кислоту , которые действуют в заднем и спинном мозге. [17] Задний мозг, например, структурирован Hox-генами , которые экспрессируются в перекрывающихся доменах вдоль переднезадней оси под контролем ретиноевой кислоты. гены 3' - (3-концевой штрих) в Hox-кластере индуцируются ретиноевой кислотой в заднем мозге, тогда как 5' - гены Hox (5-конец) не индуцируются ретиноевой кислотой и экспрессируются более сзади в спинном мозге. Hoxb-1 экспрессируется в ромбомере 4 и дает начало лицевому нерву . Без экспрессии Hoxb-1 возникает нерв, похожий на тройничный нерв .
Нейрогенез
[ редактировать ]Нейрогенез – это процесс, посредством которого нейроны образуются из нейральных стволовых клеток и клеток-предшественников . Нейроны являются «постмитотическими», что означает, что они никогда больше не будут делиться на протяжении всей жизни организма. [12]
Эпигенетические модификации играют ключевую роль в регуляции экспрессии генов при дифференцировке нервных стволовых клеток и имеют решающее значение для определения судьбы клеток в развивающемся и взрослом мозге млекопитающих. Эпигенетические модификации включают метилирование цитозина ДНК с образованием 5-метилцитозина и деметилирование 5-метилцитозина . [18] [19] Метилирование ДНК-цитозина катализируется ДНК-метилтрансферазами (DNMT) . Деметилирование метилцитозина катализируется в несколько последовательных стадий ферментами ТЕТ , которые осуществляют окислительные реакции (например, превращение 5-метилцитозина в 5-гидроксиметилцитозин ), и ферментами пути эксцизионной репарации оснований ДНК (BER). [18]
Нейрональная миграция
[ редактировать ]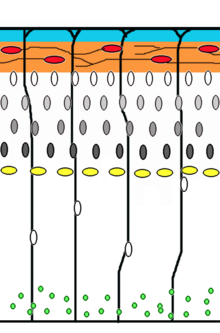
нейронов Миграция — это метод, с помощью которого нейроны перемещаются от места своего происхождения или места рождения к конечному положению в мозге. Есть несколько способов сделать это, например, радиальная миграция или тангенциальная миграция. Последовательности радиальной миграции (также известной как глиальное наведение) и сомальной транслокации были зафиксированы с помощью покадровой микроскопии . [20]

Радиальный
[ редактировать ]Нейрональные клетки-предшественники пролиферируют в желудочковой зоне развивающегося неокортекса , где основной нейральной стволовой клеткой являются радиальные глиальные клетки . Первые постмитотические клетки должны покинуть нишу стволовых клеток и мигрировать наружу, чтобы сформировать препластинку, которой суждено стать клетками Кахаля-Ретциуса и нейронами субпластинки . Эти клетки делают это путем сомальной транслокации. Нейроны, мигрирующие при таком способе передвижения, являются биполярными и прикрепляют передний край отростка к мягкой мозговой оболочке . Затем сома « транспортируется на поверхность пиальной оболочки посредством нуклеокинеза — процесса, при котором клетка» микротрубочек вокруг ядра удлиняется и сжимается вместе с центросомой , направляя ядро к конечному пункту назначения. [21] Радиальные глиальные клетки , волокна которых служат каркасом для мигрирующих клеток и средством радиальной связи, опосредованной динамической активностью кальция. [22] [23] действуют как основные возбуждающие нейрональные стволовые клетки коры головного мозга. [24] [25] или транслоцируются в кортикальную пластинку и дифференцируются либо в астроциты , либо в нейроны . [26] Сомальная транслокация может произойти в любой момент развития. [20]
Последующие волны нейронов расщепляют препластинку, мигрируя по радиальным глиальным волокнам, образуя кортикальную пластинку. Каждая волна мигрирующих клеток проходит мимо своих предшественников, образуя слои изнутри наружу, а это означает, что самые молодые нейроны находятся ближе всего к поверхности. [27] [28] Подсчитано, что миграция, управляемая глией, составляет 90% мигрирующих нейронов у человека и около 75% у грызунов. [29]
Тангенциальный
[ редактировать ]Большинство интернейронов мигрируют по касательной, используя несколько способов миграции, чтобы достичь подходящего места в коре. Примером тангенциальной миграции является перемещение интернейронов от ганглиозного возвышения к коре головного мозга. Одним из примеров продолжающейся тангенциальной миграции в зрелом организме, наблюдаемой у некоторых животных, является ростральный миграционный поток, соединяющий субвентрикулярную зону и обонятельную луковицу .
Аксофильный
[ редактировать ]Многие нейроны, мигрирующие вдоль передне-задней оси тела, используют существующие аксонные для миграции пути; это называется аксофильной миграцией. Примером такого способа миграции являются нейроны, экспрессирующие ГнРГ , которые совершают долгий путь от места своего рождения в носу через передний мозг в гипоталамус. [30] Многие механизмы этой миграции были разработаны, начиная с внеклеточных сигналов. [31] которые запускают внутриклеточную передачу сигналов. Эти внутриклеточные сигналы, такие как передача сигналов кальция , приводят к образованию актина. [32] и микротрубочки [33] цитоскелетная динамика, которая создает клеточные силы, которые взаимодействуют с внеклеточной средой через белки клеточной адгезии. [34] вызвать движение этих клеток.
Многополярный
[ редактировать ]Существует также метод миграции нейронов, называемый мультиполярной миграцией . [35] [36] Это наблюдается в мультиполярных клетках, которые у человека в изобилии присутствуют в промежуточной зоне коры . Они не похожи на клетки, мигрирующие посредством локомоции или сомальной транслокации. Вместо этого эти мультиполярные клетки экспрессируют нейрональные маркеры и расширяют множество тонких отростков в различных направлениях независимо от радиальных глиальных волокон. [35]
Нейротрофические факторы
[ редактировать ]Выживание нейронов регулируется факторами выживания, называемыми трофическими факторами. Нейротрофическая гипотеза была сформулирована Виктором Гамбургером и Ритой Леви Монтальчини на основе исследований развивающейся нервной системы. Виктор Гамбургер обнаружил, что имплантация дополнительной конечности развивающемуся цыпленку приводит к увеличению количества спинальных мотонейронов. Первоначально он думал, что дополнительная конечность вызывает пролиферацию мотонейронов, но позже он и его коллеги показали, что во время нормального развития происходит значительная гибель мотонейронов, а дополнительная конечность предотвращает эту гибель клеток. Согласно нейротрофической гипотезе, растущие аксоны конкурируют за ограниченное количество целевых трофических факторов, а аксоны, которые не получают достаточной трофической поддержки, погибают в результате апоптоза. Теперь ясно, что факторы, вырабатываемые рядом источников, способствуют выживанию нейронов.
- Фактор роста нервов (NGF): Рита Леви Монтальчини и Стэнли Коэн очистили первый трофический фактор, фактор роста нервов (NGF), за что получили Нобелевскую премию. Существует три связанных с NGF трофических фактора: BDNF, NT3 и NT4, которые регулируют выживаемость различных популяций нейронов. Белки Trk действуют как рецепторы для NGF и связанных с ним факторов. Trk представляет собой рецепторную тирозинкиназу. Димеризация и фосфорилирование Trk приводит к активации различных внутриклеточных сигнальных путей, включая пути киназы MAP, Akt и PKC.
- CNTF: Цилиарный нейротрофический фактор — это еще один белок, который действует как фактор выживания мотонейронов. CNTF действует через рецепторный комплекс, включающий CNTFRα, GP130 и LIFRβ. Активация рецептора приводит к фосфорилированию и привлечению киназы JAK, которая, в свою очередь, фосфорилирует LIFR β. LIFRβ действует как место стыковки транскрипционных факторов STAT. JAK-киназа фосфорилирует белки STAT, которые диссоциируют от рецептора и перемещаются в ядро, регулируя экспрессию генов.
- GDNF: нейротрофический фактор глиального происхождения является членом семейства белков TGFb и является мощным трофическим фактором для нейронов полосатого тела. Функциональный рецептор представляет собой гетеродимер, состоящий из рецепторов типа 1 и типа 2. Активация рецептора 1 типа приводит к фосфорилированию белков Smad, которые перемещаются в ядро и активируют экспрессию генов.
Образование синапсов
[ редактировать ]Нервно-мышечный переход
[ редактировать ]Большая часть нашего понимания формирования синапсов основана на исследованиях нервно-мышечных соединений. Передатчиком в этом синапсе является ацетилхолин. Рецептор ацетилхолина (AchR) присутствует на поверхности мышечных клеток до образования синапсов. Прибытие нерва вызывает скопление рецепторов в синапсе. МакМахан и Сейнс показали, что синаптогенный сигнал концентрируется в базальной пластинке . Они также показали, что синаптогенный сигнал вырабатывается нервом, и идентифицировали этот фактор как Агрин . Агрин индуцирует кластеризацию AchR на поверхности мышц, и образование синапсов нарушается у мышей, нокаутированных по агрину. Агрин преобразует сигнал через рецептор MuSK в рапсин . Фишбах и его коллеги показали, что субъединицы рецептора избирательно транскрибируются из ядер рядом с синаптическим участком. Это опосредуется нейрегулинами.
В зрелом синапсе каждое мышечное волокно иннервируется одним мотонейроном. Однако во время развития многие волокна иннервируются несколькими аксонами. Лихтман и его коллеги изучили процесс устранения синапсов. [37] Это событие, зависящее от активности. Частичная блокада рецептора приводит к ретракции соответствующих пресинаптических терминалей. Позднее они применили коннектомный подход, т. е. проследив все связи между мотонейронами и мышечными волокнами, чтобы охарактеризовать элиминацию синапсов развития на уровне полного контура. Анализ подтвердил масштабную перестройку, 10-кратное уменьшение количества синапсов, которая происходит, когда аксоны сокращают свои двигательные единицы, но добавляют больше синаптических областей в НМС, с которыми они остаются в контакте. [38]
синапсы ЦНС
[ редактировать ]Агрин, по-видимому, не является центральным медиатором образования синапсов ЦНС, и существует активный интерес к идентификации сигналов, которые опосредуют синаптогенез ЦНС. Нейроны в культуре развивают синапсы, подобные тем, которые образуются in vivo, что позволяет предположить, что синаптогенные сигналы могут правильно функционировать in vitro. Исследования синаптогенеза ЦНС были сосредоточены в основном на глутаматергических синапсах. Эксперименты с визуализацией показывают, что дендриты очень динамичны во время развития и часто инициируют контакт с аксонами. За этим следует рекрутирование постсинаптических белков в место контакта. Стивен Смит и его коллеги показали, что контакт, инициированный дендритными филоподиями, может перерасти в синапсы.
Индукция образования синапсов глиальными факторами: Баррес и его коллеги сделали наблюдение, что факторы в кондиционированных глией средах индуцируют образование синапсов в культурах ганглиозных клеток сетчатки. Образование синапсов в ЦНС коррелирует с дифференцировкой астроцитов, что позволяет предположить, что астроциты могут обеспечивать синаптогенный фактор. Идентичность астроцитарных факторов пока неизвестна.
Нейролигины и SynCAM как синаптогенные сигналы: Судхоф, Серафини, Шайффеле и коллеги показали, что нейролигины и SynCAM могут действовать как факторы, индуцирующие пресинаптическую дифференцировку. Нейролигины концентрируются в постсинаптическом участке и действуют через нейрексины, сконцентрированные в пресинаптических аксонах. SynCAM — это молекула клеточной адгезии, которая присутствует как в пре-, так и в постсинаптических мембранах.
Сборка нейронных цепей
[ редактировать ]Обычно считается, что процессы миграции , дифференцировки и направления аксонов нейронов являются независимыми от активности механизмами и основаны на запрограммированных генетических программах в самих нейронах. Однако результаты исследований указывают на роль механизмов, зависящих от активности, в опосредовании некоторых аспектов этих процессов, таких как скорость миграции нейронов, [39] аспекты дифференцировки нейронов [40] и поиск пути аксонов. [41] Зависимые от активности механизмы влияют на развитие нейронных цепей и имеют решающее значение для построения ранних карт связей и дальнейшего совершенствования синапсов, которое происходит во время развития. [42] Существует два различных типа нейронной активности, которые мы наблюдаем в развивающихся цепях: ранняя спонтанная активность и сенсорно-вызванная активность. Спонтанная активность возникает на ранних стадиях развития нейронных цепей , даже когда сенсорная информация отсутствует, и наблюдается во многих системах, таких как развивающаяся зрительная система , [43] [44] слуховая система , [45] [46] двигательная система , [47] гиппокамп , [48] мозжечок [49] и неокортекс . [50]
Экспериментальные методы, такие как прямая электрофизиологическая регистрация, флуоресцентная визуализация с использованием индикаторов кальция и оптогенетические методы, пролили свет на природу и функцию этих ранних всплесков активности. [51] [52] В процессе развития они имеют четкие пространственные и временные закономерности. [53] и их абляция во время развития, как известно, приводит к дефициту совершенствования сети в зрительной системе. [54] В незрелой сетчатке волны спонтанных потенциалов действия возникают из ганглиозных клеток сетчатки и распространяются по поверхности сетчатки в первые несколько недель после рождения. [55] Эти волны опосредуются нейромедиатором ацетилхолином на начальной стадии, а затем глутаматом . [56] Считается, что они инструктируют формирование двух сенсорных карт — ретинотопической карты и специфической для глаз сегрегации. [57] Уточнение ретинотопической карты происходит в нижних зрительных мишенях головного мозга — верхнем холмике (SC) и дорсально -латеральном коленчатом ядре (LGN). [58] Фармакологические нарушения и модели на мышах, у которых отсутствует субъединица β2 никотинового рецептора ацетилхолина, показали, что отсутствие спонтанной активности приводит к заметным дефектам ретинотопии и специфической для глаз сегрегации. [57]
Недавние исследования подтверждают, что микроглия , резидентная иммунная клетка головного мозга, устанавливает прямые контакты с телами клеток развивающихся нейронов и посредством этих связей регулирует нейрогенез, миграцию, интеграцию и формирование нейронных сетей в зависимости от активности. [59]
В развивающейся слуховой системе развивающаяся улитка генерирует всплески активности, которые распространяются на внутренние волосковые клетки и нейроны спиральных ганглиев , которые передают слуховую информацию в мозг. [60] Высвобождение АТФ из поддерживающих клеток запускает потенциалы действия во внутренних волосковых клетках . [61] Считается, что в слуховой системе спонтанная активность участвует в формировании тонотопической карты путем разделения аксонов улитковых нейронов, настроенных на высокие и низкие частоты. [60] В двигательной системе периодические всплески спонтанной активности вызываются возбуждающими ГАМК и глутаматом на ранних стадиях и ацетилхолином и глутаматом на более поздних стадиях. [62] В развивающемся рыбок данио спинном мозге ранняя спонтанная активность необходима для формирования все более синхронных чередующихся всплесков между ипсилатеральными и контралатеральными областями спинного мозга и для интеграции новых клеток в цепь. [63] Считается, что мотонейроны, иннервирующие одни и те же сокращающиеся мышечные волокна, поддерживают синхронную активность, что позволяет обоим нейронам оставаться в контакте с мышечными волокнами во взрослом возрасте. [38] В коре головного мозга ранние волны активности наблюдались в мозжечке и корковых срезах. [64] Как только сенсорный стимул становится доступным, окончательная точная настройка карт сенсорного кодирования и совершенствование схемы начинают все больше и больше полагаться на сенсорно-вызванную активность, как демонстрируют классические эксперименты о последствиях сенсорной депривации в критические периоды . [64]
Современные методы диффузионно-взвешенной МРТ также могут раскрыть макроскопический процесс развития аксонов. Коннектом u можно построить на основе данных диффузионной МРТ : вершины графа соответствуют анатомически помеченным областям серого вещества, а две такие вершины, скажем, и v , соединяются ребром, если на трактографическом этапе обработки данных обнаруживается аксональное волокно. который соединяет две области, соответствующие u и v .
Многочисленные мозговые диаграммы, рассчитанные в рамках проекта Human Connectome, можно загрузить с сайта http://braingraph.org . Динамика консенсусного коннектома (CCD) — это замечательное явление, которое было обнаружено путем постоянного уменьшения минимального параметра достоверности в графическом интерфейсе эталонного сервера коннектома в Будапеште . [65] [66] Будапештский эталонный сервер коннектома ( http://connectome.pitgroup.org ) отображает церебральные связи n=418 субъектов с частотным параметром k: Для любого k=1,2,...,n можно просмотреть график ребер, присутствующих не менее чем в k коннектомах. Если параметр k уменьшать по одному от k=n до k=1, то в графе появляется все больше и больше ребер, поскольку условие включения ослабляется. Удивительным наблюдением является то, что внешний вид краев далеко не случайен: он напоминает растущую сложную структуру, например, дерево или кустарник (изображено на анимации слева).
Предполагается, что в [67] что растущая структура копирует аксональное развитие человеческого мозга : самые ранние развивающиеся связи (аксональные волокна) являются общими у большинства испытуемых, а развивающиеся впоследствии связи имеют все большую дисперсию, поскольку их дисперсии накапливаются в процессе аксонального развития. разработка.
Удаление синапсов
[ редактировать ]За каждое нервно-мышечное соединение конкурируют несколько мотонейронов, но до взрослого возраста доживает только один. [37] Было показано, что конкуренция in vitro включает ограниченное количество нейротрофического вещества, которое высвобождается, или что нервная активность дает преимущество сильным постсинаптическим связям, обеспечивая устойчивость к токсину, также высвобождаемому при стимуляции нерва. In vivo предполагается, что мышечные волокна выбирают самый сильный нейрон посредством ретроградного сигнала или что зависящие от активности механизмы устранения синапсов определяют идентичность «победившего» аксона на концевой пластинке мотора. [38]
Картирование
[ редактировать ]Картирование мозга может показать, как мозг животного меняется на протяжении всей его жизни. По состоянию на 2021 год ученые нанесли на карту и сравнили весь мозг восьми червей C. elegans по их развитию на нейрональном уровне. [68] [69] и полная проводка одной мышцы млекопитающего от рождения до взрослой жизни. [38]
Взрослый нейрогенез
[ редактировать ]Нейрогенез также происходит в определенных частях мозга взрослого человека.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Неврологические признаки и заболевания» . 2 ноября 2016 г. Архивировано из оригинала 2 ноября 2016 г. Проверено 1 мая 2020 г.
- ^ «Дефекты нервной трубки» . Проверено 6 декабря 2011 г.
- ^ Jump up to: а б Гилберт С. (2006). Биология развития (8-е изд.). Издательство Sinauer Associates. стр. 373–379 . ISBN 978-0-87893-250-4 .
- ^ Чжоу Ю, Сун Х, Мин Г.Л. (январь 2024 г.). «Генетика развития мозга человека» . Обзоры природы. Генетика . 25 (1): 26–45. дои : 10.1038/s41576-023-00626-5 . ПМЦ 10926850 . ПМИД 37507490 .
- ^ Вулперт Л. (2015). Принципы развития (Пятое изд.). Издательство Оксфордского университета. ISBN 978-0-19-967814-3 . OCLC 914509705 .
- ^ Вулперт 2015 , стр. 522–526.
- ^ Саладин К. (2011). Анатомия и физиология. Единство формы и функции . Нью-Йорк: МакГроу Хилл. п. 514. ИСБН 978-0-07-337825-1 .
- ^ Шенвольф Г.К., Смит Дж.Л. (2000). «Механизмы нейруляции». В Туан RS, Ло CW (ред.). Протоколы биологии развития: Том II . Методы молекулярной биологии. Том. 136. Тотова, Нью-Джерси: Humana Press. стр. 125–134. дои : 10.1385/1-59259-065-9:125 . ISBN 978-1-59259-065-0 . ПМИД 10840705 .
- ^ Jump up to: а б Гато А., Алонсо М.И., Мартин С., Батчер Е., Моро Х.А., Де ла Мано А. и др. (август 2014 г.). «Эмбриональная спинномозговая жидкость в развитии мозга: контроль нейрональных предшественников» . Хорватский медицинский журнал . 55 (4): 299–305. дои : 10.3325/cmj.2014.55.299 . ПМЦ 4157377 . ПМИД 25165044 .
- ^ Гилберт С. (2013). Биология развития (Десятое изд.). Sinauer Associates Inc. ISBN 978-1-60535-192-6 . [ нужна страница ]
- ^ Чжоу Ю, Сун Х, Мин Г.Л. (январь 2024 г.). «Генетика развития мозга человека» . Обзоры природы. Генетика . 25 (1): 26–45. дои : 10.1038/s41576-023-00626-5 . ПМЦ 10926850 . ПМИД 37507490 .
- ^ Jump up to: а б Кандель Э.Р. (2006). Принципы нейронауки (5-е изд.). Эпплтон и Ланге: МакГроу Хилл. ISBN 978-0-07-139011-8 . [ нужна страница ]
- ^ Вулперт 2015 , стр. 163.
- ^ Эстомих Мтуи, Грегори Грюнер (2006). Клиническая нейроанатомия и неврология . Филадельфия: Сондерс. п. 1. ISBN 978-1-4160-3445-2 .
- ^ Чемберс С.М., Фазано К.А., Папапетру Э.П., Томишима М., Саделен М., Студер Л. (март 2009 г.). «Высокоэффективное нейронное преобразование ES и iPS клеток человека путем двойного ингибирования передачи сигналов SMAD» . Природная биотехнология . 27 (3): 275–280. дои : 10.1038/nbt.1529 . ПМЦ 2756723 . ПМИД 19252484 .
- ^ Джесселл, Томас М., Кандел, Эрик Р., Шварц, Джеймс Х. (2000). «Глава 55». Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-0-8385-7701-1 .
- ^ Дустер Дж. (сентябрь 2008 г.). «Синтез ретиноевой кислоты и передача сигналов на раннем органогенезе» . Клетка . 134 (6): 921–931. дои : 10.1016/j.cell.2008.09.002 . ПМЦ 2632951 . ПМИД 18805086 .
- ^ Jump up to: а б Ван З, Тан Б, Хэ Ю, Джин П (март 2016 г.). «Динамика метилирования ДНК в нейрогенезе» . Эпигеномика . 8 (3): 401–414. дои : 10.2217/эпи.15.119 . ПМЦ 4864063 . ПМИД 26950681 .
- ^ Ноак Ф, Патаскар А, Шнайдер М, Бухгольц Ф, Тивари ВК, Калегари Ф (апрель 2019 г.). «Оценка и сайт-специфическое манипулирование (гидрокси-) метилированием ДНК во время кортикогенеза у мышей» . Альянс наук о жизни . 2 (2): e201900331. дои : 10.26508/lsa.201900331 . ПМК 6394126 . ПМИД 30814272 .
- ^ Jump up to: а б Надараджа Б., Брунстрем Дж. Э., Грутцендлер Дж., Вонг Р.О., Перлман А.Л. (февраль 2001 г.). «Два режима радиальной миграции в раннем развитии коры головного мозга». Природная неврология . 4 (2): 143–150. дои : 10.1038/83967 . ПМИД 11175874 . S2CID 6208462 .
- ^ Сэмюэлс Б.А., Цай Л.Х. (ноябрь 2004 г.). «Нуклеокинез освещен». Природная неврология . 7 (11): 1169–1170. дои : 10.1038/nn1104-1169 . ПМИД 15508010 . S2CID 11704754 .
- ^ Ракич П. (май 1972 г.). «Режим миграции клеток в поверхностные слои неокортекса плода обезьяны». Журнал сравнительной неврологии . 145 (1): 61–83. дои : 10.1002/cne.901450105 . ПМИД 4624784 . S2CID 41001390 .
- ^ Раш Б.Г., Акман Дж.Б., Ракич П. (февраль 2016 г.). «Двунаправленная радиальная активность Ca(2+) регулирует нейрогенез и миграцию во время раннего формирования кортикального столба» . Достижения науки . 2 (2): e1501733. Бибкод : 2016SciA....2E1733R . дои : 10.1126/sciadv.1501733 . ПМЦ 4771444 . ПМИД 26933693 .
- ^ Ноктор С.С., Флинт А.С., Вайсман Т.А., Даммерман Р.С., Кригштейн А.Р. (февраль 2001 г.). «Нейроны, происходящие из радиальных глиальных клеток, создают радиальные единицы в неокортексе». Природа . 409 (6821): 714–720. Бибкод : 2001Natur.409..714N . дои : 10.1038/35055553 . ПМИД 11217860 . S2CID 3041502 .
- ^ Тамамаки Н., Накамура К., Окамото К., Канеко Т. (сентябрь 2001 г.). «Радиальная глия является прародителем неокортикальных нейронов в развивающейся коре головного мозга». Неврологические исследования . 41 (1): 51–60. дои : 10.1016/S0168-0102(01)00259-0 . ПМИД 11535293 . S2CID 2539488 .
- ^ Мията Т., Кавагути А., Окано Х., Огава М. (сентябрь 2001 г.). «Асимметричное наследование радиальных глиальных волокон кортикальными нейронами» . Нейрон . 31 (5): 727–741. дои : 10.1016/S0896-6273(01)00420-2 . ПМИД 11567613 .
- ^ Надараджа Б., Парнавелас Дж.Г. (июнь 2002 г.). «Режимы миграции нейронов в развивающейся коре головного мозга». Обзоры природы. Нейронаука . 3 (6): 423–432. дои : 10.1038/nrn845 . ПМИД 12042877 . S2CID 38910547 .
- ^ Ракич П. (май 1972 г.). «Режим миграции клеток в поверхностные слои неокортекса плода обезьяны». Журнал сравнительной неврологии . 145 (1): 61–83. дои : 10.1002/cne.901450105 . ПМИД 4624784 . S2CID 41001390 .
- ^ Летинич К., Зончу Р., Ракич П. (июнь 2002 г.). «Происхождение ГАМКергических нейронов в неокортексе человека». Природа . 417 (6889): 645–649. Бибкод : 2002Natur.417..645L . дои : 10.1038/nature00779 . ПМИД 12050665 . S2CID 4349070 .
- ^ Рэй С. (июль 2010 г.). «От носа к мозгу: развитие нейронов гонадотропин-рилизинг-гормона-1» . Журнал нейроэндокринологии . 22 (7): 743–753. дои : 10.1111/j.1365-2826.2010.02034.x . ПМЦ 2919238 . ПМИД 20646175 .
- ^ Джакобини П., Мессина А., Рэй С., Джампьетро С., Крепальди Т., Кармелиет П. и др. (январь 2007 г.). «Фактор роста гепатоцитов действует как мотоген и направляющий сигнал для миграции нейронов, высвобождающих гонадотропин-рилизинг-гормон-1» . Журнал неврологии . 27 (2): 431–445. doi : 10.1523/JNEUROSCI.4979-06.2007 . ПМК 6672060 . ПМИД 17215404 .
- ^ Хатчинс Б.И., Кленке У., Рэй С. (июль 2013 г.). «Зависимый от высвобождения кальция поток актина в ведущем процессе опосредует аксофильную миграцию» . Журнал неврологии . 33 (28): 11361–11371. doi : 10.1523/JNEUROSCI.3758-12.2013 . ПМЦ 3724331 . ПМИД 23843509 .
- ^ Хатчинс Б.И., Рэй С. (2014). «Захват плюс-концов микротрубочек в актиновой коре способствует аксофильной миграции нейронов за счет усиления натяжения микротрубочек в ведущем отростке» . Границы клеточной нейронауки . 8 : 400. дои : 10.3389/fncel.2014.00400 . ПМК 4245908 . ПМИД 25505874 .
- ^ Паркаш Дж., Чимино И., Феррарис Н., Касони Ф., Рэй С., Кэппи Х. и др. (ноябрь 2012 г.). «Подавление β1-интегрина в клетках, высвобождающих гонадотропин, нарушает миграцию и расширение аксонов, что приводит к серьезным репродуктивным изменениям» . Журнал неврологии . 32 (47): 16992–17002. doi : 10.1523/JNEUROSCI.3057-12.2012 . ПМК 5238668 . ПМИД 23175850 .
- ^ Jump up to: а б Табата Х., Накадзима К. (ноябрь 2003 г.). «Мультиполярная миграция: третий способ радиальной миграции нейронов в развивающейся коре головного мозга» . Журнал неврологии . 23 (31): 9996–10001. doi : 10.1523/JNEUROSCI.23-31-09996.2003 . ПМК 6740853 . ПМИД 14602813 .
- ^ Надараджа Б., Алифрагис П., Вонг Р.О., Парнавелас Дж.Г. (июнь 2003 г.). «Миграция нейронов в развивающейся коре головного мозга: наблюдения, основанные на визуализации в реальном времени». Кора головного мозга . 13 (6): 607–611. дои : 10.1093/cercor/13.6.607 . ПМИД 12764035 .
- ^ Jump up to: а б Терни С.Г., Лихтман Дж.В. (26 июня 2012 г.). «Изменение результата устранения синапсов при развитии нервно-мышечных соединений in vivo: доказательства синаптической конкуренции и ее механизма» . ПЛОС Биология . 10 (6): e1001352. дои : 10.1371/journal.pbio.1001352 . ПМЦ 3383738 . ПМИД 22745601 .
- ^ Jump up to: а б с д Мейрович Ю., Канг К., Драфт Р.В., Паварино Э.К., Энао Эчеверри М.Ф., Ян Ф. и др. (сентябрь 2021 г.). «Нейромышечные коннектомы на протяжении всего развития раскрывают правила синаптического упорядочивания». биоRxiv . дои : 10.1101/2021.09.20.460480 . S2CID 237598181 .
- ^ Комуро Х., Ракич П. (август 1996 г.). «Внутриклеточные колебания Ca2+ модулируют скорость миграции нейронов» . Нейрон . 17 (2): 275–285. дои : 10.1016/s0896-6273(00)80159-2 . ПМИД 8780651 .
- ^ Гу X, Олсон EC, Спитцер, Северная Каролина (ноябрь 1994 г.). «Спонтанные всплески и волны кальция в нейронах во время ранней дифференцировки» . Журнал неврологии . 14 (11, ч. 1): 6325–6335. doi : 10.1523/JNEUROSCI.14-11-06325.1994 . ПМК 6577261 . ПМИД 7965039 .
- ^ Хэнсон М.Г., Милнер Л.Д., Ландмессер Л.Т. (январь 2008 г.). «Спонтанная ритмическая активность спинного мозга ранних цыплят влияет на четкие решения по поиску путей моторных аксонов» . Обзоры исследований мозга . 57 (1): 77–85. дои : 10.1016/j.brainresrev.2007.06.021 . ПМК 2233604 . ПМИД 17920131 .
- ^ Киркби Л.А., Сак Г.С., Фирл А., Феллер М.Б. (декабрь 2013 г.). «Роль коррелированной спонтанной активности в сборке нейронных цепей» . Нейрон . 80 (5): 1129–1144. дои : 10.1016/j.neuron.2013.10.030 . ПМК 4560201 . ПМИД 24314725 .
- ^ Губерман А.Д. (февраль 2007 г.). «Механизмы развития зрительных цепей, специфичных для глаза». Современное мнение в нейробиологии . 17 (1): 73–80. дои : 10.1016/j.conb.2007.01.005 . ПМИД 17254766 . S2CID 19418882 .
- ^ Мейстер М., Вонг Р.О., Бэйлор Д.А., Шац С.Дж. (май 1991 г.). «Синхронные всплески потенциалов действия в ганглиозных клетках развивающейся сетчатки млекопитающих». Наука . 252 (5008): 939–943. Бибкод : 1991Sci...252..939M . дои : 10.1126/science.2035024 . ПМИД 2035024 .
- ^ Липпе В.Р. (март 1994 г.). «Ритмическая спонтанная деятельность развивающейся слуховой системы птиц» . Журнал неврологии . 14 (3, часть 2): 1486–1495. doi : 10.1523/JNEUROSCI.14-03-01486.1994 . ПМК 6577532 . ПМИД 8126550 .
- ^ Джонс Т.А., Джонс С.М., Пэггетт К.С. (октябрь 2001 г.). «Изначальные ритмические взрывы в эмбриональных ганглиозных клетках улитки» . Журнал неврологии . 21 (20): 8129–8135. doi : 10.1523/JNEUROSCI.21-20-08129.2001 . ПМК 6763868 . ПМИД 11588185 .
- ^ О'Донован, MJ (февраль 1999 г.). «Происхождение спонтанной активности в развивающихся сетях нервной системы позвоночных». Современное мнение в нейробиологии . 9 (1): 94–104. дои : 10.1016/s0959-4388(99)80012-9 . ПМИД 10072366 . S2CID 37387513 .
- ^ Крепель В., Аронов Д., Хоркера И., Репреса А., Бен-Ари Ю., Коссарт Р. (апрель 2007 г.). «Связанный с родами образец несинаптической когерентной активности в развивающемся гиппокампе» . Нейрон . 54 (1): 105–120. дои : 10.1016/j.neuron.2007.03.007 . ПМИД 17408581 .
- ^ Ватт А.Дж., Кунц Х., Мори М., Нуссер З., Шёстрем П.Дж., Хойссер М. (апрель 2009 г.). «Бегущие волны в развитии коры мозжечка, опосредованные асимметричной связью клеток Пуркинье» . Природная неврология . 12 (4): 463–473. дои : 10.1038/nn.2285 . ПМЦ 2912499 . ПМИД 19287389 .
- ^ Корлью Р., Босма М.М., Муди У.Дж. (октябрь 2004 г.). «Спонтанная синхронная электрическая активность в корковых нейронах новорожденных мышей» . Журнал физиологии . 560 (Часть 2): 377–390. дои : 10.1113/jphysicalol.2004.071621 . ПМЦ 1665264 . ПМИД 15297578 .
- ^ Феллер М.Б. (апрель 1999 г.). «Спонтанная коррелированная активность в развитии нейронных цепей» . Нейрон . 22 (4): 653–656. дои : 10.1016/s0896-6273(00)80724-2 . ПМИД 10230785 .
- ^ О'Донован М.Дж., Чаб Н., Веннер П. (октябрь 1998 г.). «Механизмы спонтанной активности в развитии спинальных сетей». Журнал нейробиологии . 37 (1): 131–145. doi : 10.1002/(sici)1097-4695(199810)37:1<131::aid-neu10>3.0.co;2-h . ПМИД 9777737 .
- ^ Стаффорд Б.К., Шер А., Литке А.М., Фельдхайм Д.А. (октябрь 2009 г.). «Пространственно-временные паттерны ретинальных волн, лежащие в основе зависящего от активности уточнения ретинофугальных проекций» . Нейрон . 64 (2): 200–212. дои : 10.1016/j.neuron.2009.09.021 . ПМК 2771121 . ПМИД 19874788 .
- ^ Торборг КЛ, Феллер МБ (июль 2005 г.). «Спонтанная закономерность активности сетчатки и уточнение проекций сетчатки». Прогресс нейробиологии . 76 (4): 213–235. doi : 10.1016/j.pneurobio.2005.09.002 . ПМИД 16280194 . S2CID 24563014 .
- ^ Галли Л., Маффей Л. (октябрь 1988 г.). «Спонтанная импульсная активность ганглиозных клеток сетчатки крысы во внутриутробном периоде». Наука . 242 (4875): 90–91. Бибкод : 1988Sci...242...90G . дои : 10.1126/science.3175637 . ПМИД 3175637 .
- ^ Форд К.Дж., Феллер М.Б. (январь 2012 г.). «Сборка и разборка холинергической сети сетчатки» . Визуальная нейронаука . 29 (1): 61–71. дои : 10.1017/S0952523811000216 . ПМЦ 3982217 . ПМИД 21787461 .
- ^ Jump up to: а б Киркби Л.А., Сак Г.С., Фирл А., Феллер М.Б. (декабрь 2013 г.). «Роль коррелированной спонтанной активности в сборке нейронных цепей» . Нейрон . 80 (5): 1129–1144. дои : 10.1016/j.neuron.2013.10.030 . ПМК 4560201 . ПМИД 24314725 .
- ^ Акман Дж.Б., Бербридж Т.Дж., Крейр MC (октябрь 2012 г.). «Волны сетчатки координируют структурную активность всей развивающейся зрительной системы» . Природа . 490 (7419): 219–225. Бибкод : 2012Natur.490..219A . дои : 10.1038/nature11529 . ПМЦ 3962269 . ПМИД 23060192 .
- ^ Череп С., Шварц А.Д., Посфаи Б., Ласло З.И., Келлермайер А., Корней З. и др. (сентябрь 2022 г.). «Микроглиальный контроль развития нейронов через соматические пуринергические соединения» . Отчеты по ячейкам . 40 (12): 111369. doi : 10.1016/j.celrep.2022.111369 . ПМЦ 9513806 . ПМИД 36130488 . S2CID 252416407 .
- ^ Jump up to: а б Кандлер К., пункт А, Но Дж. (июнь 2009 г.). «Тонотопическая реорганизация развивающихся слуховых стволовых цепей» . Природная неврология . 12 (6): 711–717. дои : 10.1038/nn.2332 . ПМК 2780022 . ПМИД 19471270 .
- ^ Трич Н.С., Родригес-Контрерас А., Кринс Т.Т., Ван Х.К., Борст Х.Г., Берглс Д.Е. (сентябрь 2010 г.). «Потенциалы действия кальция в волосковых клетках определяют активность слуховых нейронов до начала слуха» . Природная неврология . 13 (9): 1050–1052. дои : 10.1038/nn.2604 . ПМЦ 2928883 . ПМИД 20676105 .
- ^ Момосе-Сато Ю, Сато К (2013). «Крупномасштабная синхронизированная активность эмбрионального ствола мозга и спинного мозга» . Границы клеточной нейронауки . 7:36 . дои : 10.3389/fncel.2013.00036 . ПМЦ 3625830 . ПМИД 23596392 .
- ^ Варп Э., Агарвал Г., Вайарт С., Фридман Д., Олдфилд К.С., Коннер А. и др. (январь 2012 г.). «Появление закономерной активности в развивающемся спинном мозге рыбок данио» . Современная биология . 22 (2): 93–102. Бибкод : 2012CBio...22...93W . дои : 10.1016/j.cub.2011.12.002 . ПМК 3267884 . ПМИД 22197243 .
- ^ Jump up to: а б Санес Д., Рех Т., Харрис В. (2012). Развитие нервной системы (Третье изд.). Берлингтон Массачусетс: Эльзевир. ISBN 978-0-12-374539-2 . OCLC 827948474 . [ нужна страница ]
- ^ Салкай Б, Керепеши С, Варга Б, Гролмуш В (май 2015 г.). «Будапештский эталонный сервер Connectome v2.0». Письма по неврологии . 595 : 60–62. arXiv : 1412.3151 . дои : 10.1016/j.neulet.2015.03.071 . ПМИД 25862487 . S2CID 6563189 .
- ^ Салкай Б, Керепеши С, Варга Б, Гролмуш В (февраль 2017 г.). «Параметрируемые консенсусные коннектомы из проекта Human Connectome: Будапештский эталонный сервер коннектома v3.0» . Когнитивная нейродинамика . 11 (1): 113–116. arXiv : 1602.04776 . дои : 10.1007/s11571-016-9407-z . ПМК 5264751 . ПМИД 28174617 .
- ^ Керепеши С, Салкай Б, Варга Б, Гролмуш В (30 июня 2016 г.). «Как направлять края коннектомов: динамика консенсусных коннектомов и развитие связей в мозгу человека» . ПЛОС ОДИН . 11 (6): e0158680. arXiv : 1509.05703 . Бибкод : 2016PLoSO..1158680K . дои : 10.1371/journal.pone.0158680 . ПМЦ 4928947 . ПМИД 27362431 .
- ^ «Почему развитие мозга крошечного червяка может пролить свет на человеческое мышление» . физ.орг . Дуглас, остров Мэн, Великобритания: Научно-исследовательский институт им. К. Луненфельда-Таненбаума. Архивировано из оригинала 20 июня 2022 года . Проверено 21 сентября 2021 г.
- ^ Витвлит Д., Малкахи Б., Митчелл Дж.К., Мейрович Ю., Бергер Д.Р., Ву Ю. и др. (август 2021 г.). «Конектомы в процессе развития раскрывают принципы созревания мозга» . Природа . 596 (7871): 257–261. Бибкод : 2021Natur.596..257W . bioRxiv 10.1101/2020.04.30.066209v3 . дои : 10.1038/s41586-021-03778-8 . ПМЦ 8756380 . ПМИД 34349261 . S2CID 236927815 .
Внешние ссылки
[ редактировать ]- Neural Development (рецензируемый журнал открытого доступа).
- Трансляция времени нейроразвития у разных видов млекопитающих
- Развивающийся мозг ребенка
- Развитие мозга
- Как бедность может изменить мозг
- Подростковый мозг

| Базы данных органов управления : Национальные |
|---|