Сукцинатдегидрогеназа
| сукцинатдегидрогеназа (сукцинат-убихинон оксидоредуктаза) | |||
|---|---|---|---|
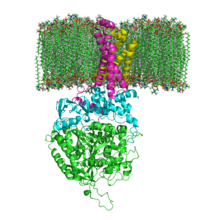 Структура SQR в фосфолипидной мембране. Сдха , СдхБ , СдхК и СдхД | |||
| Идентификаторы | |||
| Номер ЕС. | 1.3.5.1 | ||
| Номер CAS. | 9028-11-9 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| Сукцинатдегидрогеназа | |
|---|---|
| Идентификаторы | |
| Символ | Дыхательный комплекс II |
| Суперсемейство OPM | 3 |
| белок OPM | 1zoy |
| Мембраном | 656 |
Сукцинатдегидрогеназа ( SDH ) или сукцинат-коэнзим Q-редуктаза SQR ) или дыхательный комплекс II представляет собой ферментный комплекс, обнаруженный во многих бактериальных клетках и во внутренней митохондриальной мембране эукариот ( . Это единственный фермент, который участвует как в цикле лимонной кислоты , так и в цепи переноса электронов . [ 1 ] Гистохимический анализ, показывающий высокий уровень сукцинатдегидрогеназы в мышцах, демонстрирует высокое содержание митохондрий и высокий окислительный потенциал. [ 2 ]
На этапе лимонной SQR катализирует окисление 6 сукцината цикла до фумарата с восстановлением убихинона до кислоты убихинола . Это происходит во внутренней митохондриальной мембране за счет объединения двух реакций.
Структура
[ редактировать ]
Субъединицы
[ редактировать ]Митохондриальные и многие бактериальные SQR состоят из четырех структурно различных субъединиц : двух гидрофильных и двух гидрофобных . Первые две субъединицы — флавопротеин (SdhA) и железо-серный белок (SdhB) — образуют гидрофильную головку, в которой осуществляется ферментативная активность комплекса. SdhA содержит ковалентно присоединенный флавинадениндинуклеотида (FAD) кофактор и сукцината сайт связывания , а SdhB содержит три железо-серных кластера: [2Fe-2S], [4Fe-4S] и [3Fe-4S]. Вторые две субъединицы представляют собой гидрофобные субъединицы мембранного якоря, SdhC и SdhD. Митохондрии человека содержат две различные изоформы SdhA (субъединицы Fp типа I и типа II), эти изоформы также обнаружены у Ascaris suum и Caenorhabditis elegans . [ 3 ] Субъединицы образуют мембраносвязанный комплекс цитохрома b с шестью трансмембранными спиралями, содержащими одну группу гема b и сайт связывания убихинона . Две молекулы фосфолипида , одна кардиолипина и одна фосфатидилэтаноламина , также обнаружены в субъединицах SdhC и SdhD (не показаны на изображении). Они служат для того, чтобы занять гидрофобное пространство ниже гема b. Эти субъединицы показаны на прикрепленном изображении. SdhA — зеленый, SdhB — бирюзовый, SdhC — фуксия, SdhD — желтый. Вокруг SdhC и SdhD расположена фосфолипидная мембрана с межмембранным пространством вверху изображения. [ 4 ]
Таблица субъединичного состава [ 5 ]
[ редактировать ]| Нет. | Название субъединицы | Человеческий белок | Описание белка от UniProt | Семейство Pfam с человеческим белком |
|---|---|---|---|---|
| 1 | Сдха | SDHA _ЧЕЛОВЕК | Субъединица флавопротеина сукцинатдегидрогеназы [убихинон], митохондриальная | Пфам ПФ00890 , Пфам ПФ02910 |
| 2 | СдхБ | СДХБ _ЧЕЛОВЕК | Сукцинатдегидрогеназа [убихинон] железо-серная субъединица, митохондриальная | Пфам ПФ13085 , Пфам ПФ13183 |
| 3 | СдхК | C560_ЧЕЛОВЕК | Субъединица цитохрома b560 сукцинатдегидрогеназы, митохондриальная | Пфам PF01127 |
| 4 | СдхД | DHSD_HUMAN | Сукцинатдегидрогеназа [убихинон] малая субъединица цитохрома b, митохондриальная | Пфам PF05328 |
Сайт связывания убихинона
[ редактировать ]два характерных убихинона сайта связывания На SDH млекопитающих можно распознать : Q P - проксимальный к матриксу и Q D - дистальный к матриксу . Сайт связывания убихинона Qp, который проявляет более высокое сродство к убихинону, расположен в пробеле, состоящем из SdhB, SdhC и SdhD. Убихинон стабилизируется боковыми цепями His207 субъединицы B, Ser27 и Arg31 субъединицы C и Tyr83 субъединицы D. Хинонное кольцо окружено Ile28 субъединицы C и Pro160 субъединицы B. Эти остатки , наряду с Il209, Trp163 , Trp164 субъединицы B и Ser27 (атом C) субъединицы C образуют гидрофобное окружение хинон -связывающего кармана Qp. [ 6 ] Напротив, сайт связывания убихинона QD , который расположен ближе к межмембранному пространству, состоит только из SdhD и имеет более низкое сродство к убихинону. [ 7 ]
Сайт связывания сукцината
[ редактировать ]обеспечивает место связывания для окисления сукцината . SdhA Боковые цепи Thr254, His354 и Arg399 субъединицы А стабилизируют молекулу , в то время как FAD окисляет и переносит электроны к первому из железо-серных кластеров [2Fe-2S]. [ 8 ] Это можно увидеть на изображении 5.
Редокс-центры
[ редактировать ]Сайт связывания сукцината и сайт связывания убихинона соединены цепочкой окислительно-восстановительных центров, включающих FAD и кластеры железо - сера . Эта цепь простирается более чем на 40 Å через фермента мономер . Все расстояния между краями между центрами меньше предполагаемого предела физиологического переноса электронов в 14 Å . [ 4 ] Этот перенос электрона продемонстрирован на изображении 8.
Сборка и созревание
[ редактировать ]Все субъединицы митохондриальной СДГ человека закодированы в ядре. После трансляции субъединица SDHA транслоцируется в виде апопротеина в митохондриальный матрикс. Впоследствии одним из первых шагов является ковалентное присоединение кофактора ( FAD ковалентное флавинилирование). Этот процесс усиливается фактором сборки сукцинатдегидрогеназы 2 ( SDHAF2 ; [ 9 ] также называемый Sdh5 у дрожжей и SdhE у бактерий), а также некоторыми промежуточными продуктами цикла Кребса. Фумарат наиболее сильно стимулирует ковалентное флавинилирование СДГК. [ 10 ] Исследования бактериальной системы показали, что механизм прикрепления FAD включает промежуточное соединение хинон:метид. [ 11 ] При митохондриальной, но не бактериальной сборке, SDHA взаимодействует со вторым фактором сборки, называемым фактором сборки сукцинатдегидрогеназы 4 (SDHAF4; у дрожжей называется Sdh8), прежде чем он вставляется в конечный комплекс. [ 7 ]
Fe-S Простетические группы субъединицы SDHB предварительно формируются в митохондриальном матриксе белковым комплексом ISU. Также считается, что комплекс способен вставлять железо-серные кластеры в SDHB во время его созревания. Исследования показывают, что внедрение кластера Fe-S предшествует образованию димера SDHA-SDHB. Такое включение требует уменьшения остатков цистеина в активном центре SDHB. Как восстановленные остатки цистеина, так и уже включенные кластеры Fe-S очень чувствительны к повреждению АФК . Еще два фактора сборки SDH, SDHAF1 (Sdh6) и SDHAF3 (Sdh7 у дрожжей), по-видимому, участвуют в созревании SDHB, защищая субъединицу или димер SDHA-SDHB от повреждения кластера Fe-S, вызванного АФК. [ 7 ]
Сборка гидрофобного якоря, состоящего из субъединиц SDHC и SDHD, остается неясной. Особенно в случае вставки гема b и даже его функции. Простетическая группа гема b, по-видимому, не является частью пути переноса электронов в комплексе II. [ 5 ] Кофактор скорее поддерживает стабильность якоря.
Механизм
[ редактировать ]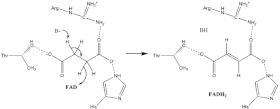
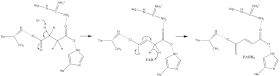
Окисление сукцината
[ редактировать ]Многое известно о сукцината окисления механизме , который включает перенос протона и гидрида. Комбинация мутагенеза и структурного анализа идентифицирует Arg-286 субъединицы SDHA ( нумерация E. coli ) как протонный челнок. Кристаллические структуры ферментов многих организмов показывают, что это хорошо подходит для этапа переноса протона. После этого существует два возможных механизма элиминации: E2 или E1cb. При исключении E2 механизм согласован. Основной остаток или кофактор депротонирует альфа-углерод , и FAD принимает гидрид от бета-углерода , окисляя связанный сукцинат до фумарата — см. изображение 6. В E1cb енолятное образуется промежуточное соединение, показанное на изображении 7, прежде чем FAD принимает гидрид . Необходимы дальнейшие исследования, чтобы определить, какой механизм элиминации сукцината осуществляется сукцинатдегидрогеназой. Окисленный фумарат , теперь свободно связанный с активным центром , может свободно покинуть белок .
Электронное туннелирование
[ редактировать ]После того, как электроны образуются в результате сукцината окисления посредством FAD , они туннелируют вдоль реле [Fe-S], пока не достигнут кластера [3Fe-4S]. Эти электроны впоследствии передаются ожидающей убихинона молекуле в активном центре . железо сера - . Система туннелирования электронов показана на изображении 9
Снижение убихинона
[ редактировать ]

O1 Карбонильный кислород убихинона в железо - ориентирован в активном центре (изображение 4) за счет взаимодействий водородной связи с Tyr83 субъединицы D. Присутствие электронов серном кластере [3Fe-4S] индуцирует движение убихинона во вторую ориентацию. Это облегчает взаимодействие второй водородной связи O4 группой между карбонильной убихинона и Ser27 субъединицы C. После первой одноэлектронного восстановления стадии семихиноновый образуется радикал. Второй электрон приходит из кластера [3Fe-4S] и обеспечивает полное восстановление убихинона до убихинола . Этот механизм восстановления убихинона показан на изображении 8.
Гемовая простетическая группа
[ редактировать ]Хотя функциональность гема в сукцинатдегидрогеназе все еще исследуется, некоторые исследования [ кем? ] утверждали, что первый электрон , доставленный к убихинону через [3Fe-4S], может туннелировать туда и обратно между гемом и промежуточным убихиноном . Таким образом, гема кофактор действует как сток электронов . Его роль заключается в предотвращении взаимодействия промежуточного продукта с молекулярным кислородом с образованием активных форм кислорода (АФК). Гемовая убихинону группа по отношению к показана на изображении 4.
Также было высказано предположение, что может существовать воротный механизм , предотвращающий электронов туннелирование непосредственно в гем из кластера [3Fe-4S]. Потенциальным кандидатом является остаток His207, расположенный непосредственно между кластером и гемом . His207 субъединицы B находится в непосредственной близости от кластера [3Fe-4S], связанного убихинона и гема ; и может модулировать поток электронов между этими окислительно-восстановительными центрами. [ 12 ]
Перенос протона
[ редактировать ]Чтобы полностью восстановить хинон в SQR, два электрона и два протона необходимы . Утверждалось, что молекула воды (HOH39) прибывает в активный центр и координируется His207 субъединицы B, Arg31 субъединицы C и Asp82 субъединицы D. Семихиноновая разновидность протонируется протонами, доставленными из HOH39, завершая убихинон. восстановление до убихинола . His207 и Asp82, скорее всего, облегчают этот процесс. что Tyr83 субъединицы D координируется с близлежащим гистидином, а также с карбонильным кислородом O1 убихинона Другие исследования утверждают , . Остаток гистидина , снижает рКа тирозина что делает его более подходящим для передачи своего протона восстановленному промежуточному убихинону .
Ингибиторы
[ редактировать ]Существует два различных класса ингибиторов (SDHI) комплекса II: те, которые связываются в сукцинатном кармане, и те, которые связываются в убихиноновом кармане. Ингибиторы убихинонового типа включают карбоксин и теноилтрифторацетон . Ингибиторы-аналоги сукцината включают синтетическое соединение малонат, а также промежуточные соединения цикла ТСА, малат и оксалоацетат . Действительно, оксалоацетат является одним из наиболее мощных ингибиторов Комплекса II. Почему обычное промежуточное соединение цикла ТСА ингибирует Комплекс II, не совсем понятно, хотя оно может играть защитную роль в минимизации опосредованного обратным переносом электронов образования супероксида Комплексом I. [ 13 ] Атпенин 5а является высокоэффективным ингибитором комплекса II, имитирующим связывание убихинона.
Ингибиторы убихинонового типа используются в качестве фунгицидов в сельском хозяйстве с 1960-х годов. Карбоксин в основном использовался для борьбы с болезнями, вызываемыми базидиомицетами, такими как стеблевая ржавчина и болезни ризоктонии . В 1980-х годах было обнаружено, что простые бензанилиды обладают активностью, сравнимой с карбоксином, и некоторые из них поступили в продажу, включая беноданил , флутоланил и мепронил . [ 14 ] Совсем недавно были разработаны другие соединения с более широким спектром действия против ряда патогенов растений, включая боскалид , флуопирам , флуксапироксад , пидифлуметофен и седаксан . [ 15 ] [ 14 ] Некоторые важные для сельского хозяйства грибы не чувствительны к ингибиторам нового поколения убихинонового типа. [ 16 ]
У FRAC есть рабочая группа [ 17 ] для SDHI и рекомендует методы управления резистентностью . [ 18 ]
Роль в болезни
[ редактировать ]Фундаментальная роль сукцинат-коэнзима Q-редуктазы в цепи переноса электронов митохондрий эмбриональной делает его жизненно важным для большинства многоклеточных организмов стадии у мышей . . Было также показано, что удаление этого фермента из генома приводит к летальному исходу на
Все мутации SDHx могут приводить к онкогенезу в хромаффинных клетках , вызывая нейроэндокринные опухоли, такие как параганглиома , феохромоцитома , карцинома почки и стромальная опухоль желудочно-кишечного тракта (ГИСО). [ 19 ] SDHA, SDHB, SDHD и SDHAF1 могут вызывать дефицит митохондриального комплекса II , что может привести к синдрому Лея , митохондриальной энцефалопатии и атрофии зрительного нерва . Мутации SDHB являются наиболее проникающими для параганглиом и феохромоцитом. [ 20 ] и мутации SDHD и SDHAF2 имеют некоторые эффекты материнского импринтинга. [ 21 ]
Сукцинатдегидрогеназа млекопитающих участвует не только в выработке энергии в митохондриях , но также играет роль в чувствительности к кислороду и опухолей подавлении ; и, следовательно, является объектом постоянных исследований.
Снижение уровня митохондриального фермента сукцинатдегидрогеназы (СДГ), основного элемента комплекса II, наблюдается посмертно в мозге пациентов с болезнью Гентингтона, а дефекты энергетического обмена выявляются как у предсимптомных, так и у симптоматических пациентов с ГБ. [ 22 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Оедотун К.С., Лемир Б.Д. (март 2004 г.). «Четвертичная структура сукцинатдегидрогеназы Saccharomyces cerevisiae. Моделирование гомологии, докинг кофакторов и исследования молекулярной динамики» . Журнал биологической химии . 279 (10): 9424–9431. дои : 10.1074/jbc.M311876200 . ПМИД 14672929 .
- ^ веб-мастер (04.03.2009). «Использование гистохимии для определения свойств мышц» . Сукцинатдегидрогеназа: определение окислительного потенциала . Калифорнийский университет, Сан-Диего . Архивировано из оригинала 10 октября 2018 г. Проверено 27 декабря 2017 г.
- ^ Томицука Э., Хираваке Х., Гото Ю., Таниваки М., Харада С., Кита К. (август 2003 г.). «Прямое свидетельство существования двух различных форм флавопротеиновой субъединицы митохондриального комплекса II человека (сукцинат-убихинонредуктаза)». Журнал биохимии . 134 (2): 191–195. дои : 10.1093/jb/mvg144 . ПМИД 12966066 .
- ^ Перейти обратно: а б Янковская В., Хорсфилд Р., Торнрот С., Луна-Чавес С., Миёши Х., Леже С. и др. (январь 2003 г.). «Архитектура сукцинатдегидрогеназы и генерации активных форм кислорода». Наука . 299 (5607): 700–704. Бибкод : 2003Sci...299..700Y . дои : 10.1126/science.1079605 . ПМИД 12560550 . S2CID 29222766 .
- ^ Перейти обратно: а б Сунь Ф, Хо X, Чжай Ю, Ван А, Сюй Дж, Су Д и др. (июль 2005 г.). «Кристаллическая структура белкового комплекса дыхательной мембраны митохондрий II» . Клетка . 121 (7): 1043–1057. дои : 10.1016/j.cell.2005.05.025 . ПМИД 15989954 .
- ^ Хорсфилд Р., Янковская В., Секстон Г., Уиттингем В., Шиоми К., Омура С. и др. (март 2006 г.). «Структурный и вычислительный анализ хинонсвязывающего сайта комплекса II (сукцинат-убихинон оксидоредуктаза): механизм переноса электрона и протонной проводимости при восстановлении убихинона» . Журнал биологической химии . 281 (11): 7309–7316. дои : 10.1074/jbc.M508173200 . ПМИД 16407191 .
- ^ Перейти обратно: а б с Ван Вранкен Дж. Г., На У, Винге Д. Р., Раттер Дж. (декабрь 2014 г.). «Белково-опосредованная сборка сукцинатдегидрогеназы и ее кофакторов» . Критические обзоры по биохимии и молекулярной биологии . 50 (2): 168–180. дои : 10.3109/10409238.2014.990556 . ПМЦ 4653115 . ПМИД 25488574 .
- ^ Кенни WC (апрель 1975 г.). «Реакция N-этилмалеимида в активном центре сукцинатдегидрогеназы» . Журнал биологической химии . 250 (8): 3089–3094. дои : 10.1016/S0021-9258(19)41598-6 . ПМИД 235539 .
- ^ Шарма П., Маклашина Е., Чеккини Дж., Айверсон Т.М. (сентябрь 2020 г.). «Роль SDHAF2 и дикарбоксилата в ковалентном флавинилировании SDHA, флавопротеина комплекса II человека» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (38): 23548–23556. Бибкод : 2020PNAS..11723548S . дои : 10.1073/pnas.2007391117 . ПМЦ 7519310 . ПМИД 32887801 .
- ^ Маклашина Е, Айверсон Т.М., Чеккини Дж. (октябрь 2022 г.). «Как фактор сборки усиливает ковалентное прикрепление FAD к субъединице флавопротеина комплекса II» . Журнал биологической химии . 298 (10): 102472. doi : 10.1016/j.jbc.2022.102472 . ПМЦ 9557727 . ПМИД 36089066 .
- ^ Шарма П., Маклашина Е., Чеккини Дж., Айверсон Т.М. (январь 2018 г.). «Кристаллическая структура промежуточного продукта сборки дыхательного комплекса II» . Природные коммуникации . 9 (1): 274. Бибкод : 2018NatCo...9..274S . дои : 10.1038/s41467-017-02713-8 . ПМЦ 5773532 . ПМИД 29348404 .
- ^ Тран К.М., Ротери Р.А., Маклашина Е., Чеккини Г., Вайнер Дж.Х. (октябрь 2006 г.). «Участок связывания хинона в сукцинатдегидрогеназе Escherichia coli необходим для переноса электрона на гем b» . Журнал биологической химии . 281 (43): 32310–32317. дои : 10.1074/jbc.M607476200 . ПМИД 16950775 .
- ^ Мюллер Ф.Л., Лю Ю., Абдул-Гани М.А., Люстгартен М.С., Бхаттачарья А., Джанг Ю.К. и др. (январь 2008 г.). «Высокие темпы продукции супероксида в митохондриях скелетных мышц, дышащих как на комплексных I-, так и на комплексных II-связанных субстратах». Биохимический журнал . 409 (2): 491–499. дои : 10.1042/BJ20071162 . ПМИД 17916065 .
- ^ Перейти обратно: а б Уолтер Х (2016). «Фунгицидные карбоксамиды, ингибирующие сукцинатдегидрогеназу». Классы биоактивных карбоновых соединений: фармацевтика и агрохимия . стр. 405–425. дои : 10.1002/9783527693931.ch31 . ISBN 978-3-527-33947-1 .
- ^ Авенот Х.Ф., Михаилидис Т.Дж. (2010). «Прогресс в понимании молекулярных механизмов и эволюции устойчивости к фунгицидам, ингибирующим сукцинатдегидрогеназу (SDHI), у фитопатогенных грибов». Защита урожая . 29 (7): 643–651. Бибкод : 2010CrPro..29..643A . дои : 10.1016/j.cropro.2010.02.019 .
- ^
- Лукас Дж.А., Хокинс, Нью-Джерси, Фраайе Б.А. (2015). Эволюция устойчивости к фунгицидам . Достижения прикладной микробиологии. Полный. 90. стр. 29–92. дои : 10.1016/bs.aambs.2014.09.001 . ISBN 978-0-12-802275-7 . ПМИД 25596029 .
- Дюбос Т., Паскуали М., Погода Ф., Казанова А., Хоффманн Л., Бейер М. (январь 2013 г.). «Различия между последовательностями сукцинатдегидрогеназы чувствительных к изопиразаму штаммов Zymoseptoria tritici и нечувствительных штаммов Fusarium graminearum». Биохимия и физиология пестицидов . 105 (1): 28–35. Бибкод : 2013PBioP.105...28D . дои : 10.1016/j.pestbp.2012.11.004 . ПМИД 24238287 .
- ^ «Рабочая группа по фунгицидам SDHI» . FRAC ( Комитет по борьбе с устойчивостью к фунгицидам ) . 31 января 2020 г. Проверено 5 июля 2022 г.
- ^ «Рекомендации по SDHI» . ФРАК . Март 2020 года . Проверено 5 июля 2022 г.
- ^ Барлетта Дж. А., Хорник Дж. Л. (июль 2012 г.). «Опухоли с дефицитом сукцинатдегидрогеназы: достижения диагностики и клинические последствия». Достижения анатомической патологии . 19 (4): 193–203. дои : 10.1097/PAP.0b013e31825c6bc6 . ПМИД 22692282 . S2CID 32088940 .
- ^ Райкен Дж.А., Нимейер Н.Д., Йонкер М.А., Эйкеленкамп К., Янсен Дж.К., ван Беркель А. и др. (январь 2018 г.). «Пенетрантность параганглиомы и феохромоцитомы у носителей зародышевой мутации SDHB». Клиническая генетика . 93 (1): 60–66. дои : 10.1111/cge.13055 . ПМИД 28503760 .
- ^ Байсал Б.Е. (май 2013 г.). «Митохондриальный комплекс II и геномный импринтинг при наследовании опухолей параганглиомы». Биохимика и биофизика Acta . 1827 (5): 573–577. дои : 10.1016/j.bbabio.2012.12.005 . ПМИД 23291190 .
- ^ Скиллингс Э.А., Мортон А.Дж. (2016). «Отсроченное начало и снижение когнитивных нарушений посредством предварительного кондиционирования с помощью 3-нитропропионовой кислоты зависит от пола и длины повтора CAG в мышиной модели болезни Хантингтона R6/2». Журнал болезни Хантингтона . 5 (1): 19–32. дои : 10.3233/JHD-160189 . ПМИД 27031731 .