надзор за мРНК
Механизмы наблюдения за мРНК — это пути, используемые организмами для обеспечения точности и качества молекул информационной РНК (мРНК). Внутри клеток существует ряд механизмов наблюдения. Эти механизмы действуют на различных этапах пути биогенеза мРНК , обнаруживая и разрушая транскрипты, которые не были процессированы должным образом.
Обзор
[ редактировать ]Перевод транскриптов информационной РНК в белки является жизненно важной частью центральной догмы молекулярной биологии . Молекулы мРНК, однако, склонны к множеству ошибок точности, которые могут вызвать ошибки в трансляции мРНК в качественные белки . [ 1 ] Механизмы наблюдения за РНК — это методы, которые клетки используют для обеспечения качества и точности молекул мРНК. [ 2 ] Обычно это достигается путем маркировки аберрантной молекулы мРНК для деградации различными эндогенными нуклеазами . [ 3 ]
Наблюдение за мРНК было зарегистрировано у бактерий и дрожжей . Известно, что у эукариот эти механизмы действуют как в ядре , так и в цитоплазме . [ 4 ] Проверка точности молекул мРНК в ядре приводит к деградации неправильно обработанных транскриптов перед экспортом в цитоплазму. Транскрипты подлежат дальнейшему наблюдению после попадания в цитоплазму. Механизмы цитоплазматического надзора оценивают транскрипты мРНК на наличие или отсутствие преждевременных стоп-кодонов. [ 3 ] [ 4 ]
В настоящее время известно, что внутри клеток функционируют три механизма наблюдения : путь распада мРНК, опосредованный нонсенсом (NMD); безостановочные пути распада мРНК (NSD); и путь распада мРНК, опосредованный no-go (NGD).
Нонсенс-опосредованный распад мРНК
[ редактировать ]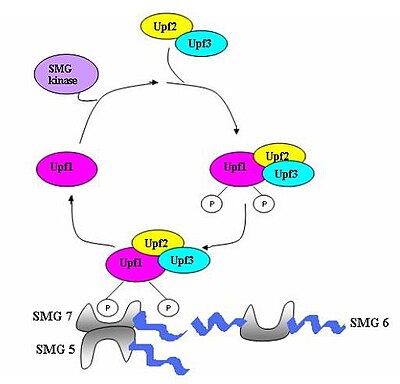
Обзор
[ редактировать ]Нонсенс-опосредованный распад участвует в обнаружении и распаде транскриптов мРНК, содержащих кодоны преждевременного терминирования (PTC). ПТК могут возникать в клетках посредством различных механизмов: зародышевых мутаций в ДНК; соматические мутации ДНК; ошибки в транскрипции ; или ошибки в посттранскрипционной обработке мРНК. [ 5 ] [ 6 ] Неспособность распознать и разрушить эти транскрипты мРНК может привести к образованию усеченных белков, которые могут быть вредными для организма. Вызывая распад на С-конце укороченных полипептидов, механизм NMD может защитить клетки от вредных доминантно -негативных эффектов и эффектов усиления функции . [ 7 ] ПТК вовлечены примерно в 30% всех наследственных заболеваний; как таковой, путь NMD играет жизненно важную роль в обеспечении общего выживания и приспособленности организма. [ 8 ] [ 9 ]
Комплекс наблюдения, состоящий из различных белков ( eRF1 , eRF3 , Upf1 , Upf2 и Upf3), собирается и сканирует мРНК на наличие преждевременных стоп-кодонов. [ 5 ] Сборка этого комплекса запускается преждевременной терминацией трансляции. Если обнаруживается преждевременный стоп-кодон, то транскрипт мРНК получает сигнал о деградации - происходит сочетание обнаружения с деградацией. [ 3 ] [ 10 ] [ 11 ]
Семь генов smg (smg1-7) и три гена UPF (Upf1-3) были идентифицированы у Saccharomyces cerevisiae и Caenorhabditis elegans как важные транс-действующие факторы, способствующие активности NMD. [ 12 ] [ 13 ] Все эти гены консервативны у Drosophila melanogaster и других млекопитающих, где они также играют решающую роль в NMD. У эукариот есть три компонента, которые сохраняются в процессе NMD. [ 14 ] Это комплексы Upf1/SMG-2, Upf2/SMG-3 и Upf3/SMG-4. Upf1/SMG-2 представляет собой фосфопротеин в многоклеточных организмах и, как полагают, способствует NMD благодаря своей активности фосфорилирования. Однако точные взаимодействия белков и их роль в NMD в настоящее время оспариваются. [ 11 ] [ 12 ] [ 14 ] [ 15 ] [ 16 ]
Механизм у млекопитающих
[ редактировать ]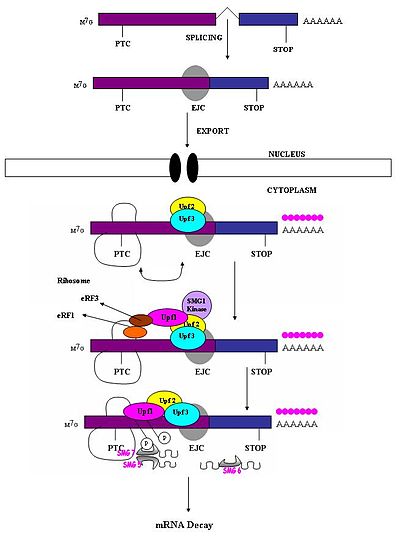
Преждевременный стоп-кодон должен быть признан отличным от нормального стоп-кодона, чтобы только первый вызывал реакцию NMD. Было замечено, что способность нонсенс-кодона вызывать деградацию мРНК зависит от его относительного расположения по отношению к следующему элементу последовательности и связанным с ним белкам. [ 1 ] Исследования показали, что нуклеотиды , расположенные более чем на 50–54 нуклеотидов выше последнего соединения экзон-экзон, могут нацеливать мРНК на распад. [ 1 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] [ 17 ] Те, кто находится ниже по течению от этого региона, не могут этого сделать. Таким образом, нонсенс-кодоны лежат более чем на 50-54 нуклеотида выше границы последнего экзона , тогда как естественные стоп-кодоны расположены внутри терминальных экзонов. [ 18 ] Комплексы экзонов (EJC) отмечают границы экзонов. EJC представляют собой мультибелковые комплексы, которые собираются во время сплайсинга в положении примерно на 20–24 нуклеотида выше места сплайсинга. [ 19 ] Именно этот EJC предоставляет информацию о положении, необходимую для отличия преждевременных стоп-кодонов от естественных стоп-кодонов. Распознавание PTCs, по-видимому, зависит от определения экзон-экзонных соединений. Это предполагает участие сплайсосомы в NMD млекопитающих. [ 17 ] [ 20 ] Исследования изучили возможность участия сплайсосом в НМД млекопитающих и определили, что это вполне вероятно. [ 18 ] Более того, было замечено, что механизмы NMD не активируются нонсенс-транскриптами, которые генерируются из генов, которые в природе не содержат интронов (например, гистона H4, Hsp70, рецептора меланокортина-4). [ 7 ]
Когда рибосома достигает PTC, факторы трансляции eRF1 и eRF3 взаимодействуют с сохранившимися комплексами EJC через мультибелковый мостик. [ 21 ] Взаимодействия UPF1 с терминирующим комплексом и с UPF2 /UPF3 сохранившихся EJCs имеют решающее значение. Именно эти взаимодействия нацелены на мРНК для быстрого распада эндогенными нуклеазами. [ 18 ] [ 21 ]

Механизм у беспозвоночных
[ редактировать ]Исследования с участием таких организмов, как S. cerevisiae , D.melanogaster и C. elegans, показали, что распознавание PTC беспозвоночными организмами не затрагивает экзон-экзонные границы. [ 20 ] Эти исследования показывают, что NMD у беспозвоночных возникает независимо от сплайсинга. В результате EJC, которые отвечают за маркировку экзон-экзонных границ, не требуются у беспозвоночных NMD. [ 3 ] Было предложено несколько моделей, объясняющих, чем PTC отличаются от нормальных стоп-кодонов у беспозвоночных. Один из них предполагает, что может существовать нижележащий элемент последовательности, который функционирует аналогично соединениям экзонов у млекопитающих. [ 11 ] Вторая модель предполагает, что широко распространенная особенность мРНК, такая как 3'-поли-А-хвост, может предоставлять позиционную информацию, необходимую для распознавания. [ 22 ] Другая модель, получившая название «модель искусственной 3'UTR», предполагает, что преждевременную терминацию трансляции можно отличить от нормальной терминации из-за внутренних особенностей, которые могут позволить ей распознавать свое присутствие в неподходящей среде. [ 3 ] Однако эти механизмы еще предстоит убедительно доказать.
Механизм у растений
[ редактировать ]У растений существует два механизма узнавания ПТК: по удаленности от ЭОК (как у позвоночных) или от поли-А-хвоста. Механизм NMD у растений вызывает распад мРНК, содержащих 3'UTR длиной более 300 нуклеотидов, поэтому доля мРНК с более длинными 3'UTR у растений значительно ниже, чем у позвоночных. [ 23 ] [ 24 ]
Уклонение от национальной ПРО
[ редактировать ]Обычно считается, что мРНК с нонсенс-мутациями подвергаются распаду по путям NMD. Присутствие этого преждевременного стоп-кодона примерно в 50-54 нуклеотидах 5' от соединения экзонов, по-видимому, является триггером быстрого распада; однако было замечено, что некоторые молекулы мРНК с преждевременным стоп-кодоном способны избегать обнаружения и распада. [ 17 ] [ 25 ] В общем, эти молекулы мРНК обладают стоп-кодоном очень рано в рамке считывания (т.е. PTC находится проксимальнее AUG). Это, по-видимому, противоречит принятой в настоящее время модели НМД, поскольку это положение находится значительно на расстоянии 5' от экзон-экзонного соединения. [ 26 ]
Это было продемонстрировано на β-глобулине. мРНК β-глобулина, содержащие нонсенс-мутацию в начале первого экзона гена, более стабильны, чем молекулы мРНК, чувствительные к NMD. Точный механизм предотвращения обнаружения в настоящее время неизвестен. Было высказано предположение, что поли-А-связывающий белок (PABP), по-видимому, играет роль в этой стабильности. [ 27 ] В других исследованиях было продемонстрировано, что присутствие этого белка вблизи AUG-проксимальных PTC, по-видимому, способствует стабильности этих мРНК, чувствительных к NMD. Было замечено, что этот защитный эффект не ограничивается только промотором β-глобулина. [ 25 ] Это предполагает, что этот механизм предотвращения НМД может преобладать в других типах тканей для различных генов. Текущую модель НМД, возможно, придется пересмотреть после дальнейших исследований.
Безостановочный распад мРНК
[ редактировать ]Обзор
[ редактировать ]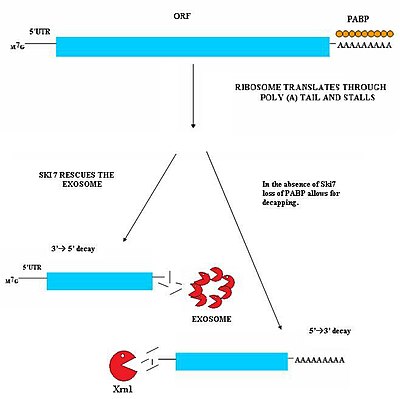
Непрерывный опосредованный распад (NSD) участвует в обнаружении и распаде транскриптов мРНК, у которых отсутствует стоп-кодон. [ 29 ] [ 30 ] Эти транскрипты мРНК могут возникать в результате множества различных механизмов, таких как сигналы преждевременного 3'-аденилирования или загадочного полиаденилирования в кодирующей области гена. [ 31 ] Отсутствие стоп-кодона представляет собой серьезную проблему для клеток. Рибосомы, транслирующие мРНК, в конечном итоге транслируются в 3'-поли-А-хвостовую область транскриптов и киосков. В результате он не может извлечь мРНК. [ 32 ] Таким образом, рибосомы могут оказаться изолированными, связанными с непрерывной мРНК, и будут недоступны для трансляции других молекул мРНК в белки. Безостановочный опосредованный распад решает эту проблему, освобождая остановившиеся рибосомы и маркируя безостановочную мРНК для деградации в клетке нуклеазами. Безостановочный опосредованный распад состоит из двух различных путей, которые, вероятно, действуют согласованно, обеспечивая безостановочный распад мРНК. [ 29 ] [ 30 ]
Трасса Ski7
[ редактировать ]Этот путь активен, когда белок Ski7 доступен в клетке. Считается, что белок Ski7 связывается с пустым А-сайтом рибосомы. Это связывание позволяет рибосоме безостановочно выбросить застрявшую молекулу мРНК – это даже освобождает рибосому и позволяет ей транслировать другие транскрипты. Ski7 теперь связан с непрерывной мРНК, и именно эта ассоциация нацелена на непрерывную мРНК для распознавания цитозольной экзосомой . Комплекс Ski7-экзосома быстро деаденилирует молекулу мРНК, что позволяет экзосоме расщеплять транскрипт по направлению от 3' до 5'. [ 29 ] [ 30 ]
Трасса, не предназначенная для катания на лыжах7
[ редактировать ]Второй тип NSD наблюдался у дрожжей. В этом механизме отсутствие Ski7 приводит к потере поли-А-хвоста, связывающего белки PABP, под действием трансляционной рибосомы. Удаление этих белков PABP затем приводит к потере защитного кэпа 5'm7G . Потеря кэпа приводит к быстрой деградации транскрипта эндогенной 5'-3' экзонуклеазой, такой как XrnI. [ 30 ]
Бесполезный распад
[ редактировать ]
No-Go распада (NGD) является последним обнаруженным механизмом наблюдения. [ 33 ] Как таковое, оно в настоящее время недостаточно изучено. Хотя подлинные мишени NGD плохо изучены, они, по-видимому, состоят в основном из транскриптов мРНК, на которых рибосомы остановились во время трансляции. Эта остановка может быть вызвана множеством факторов, включая сильные вторичные структуры , которые могут физически блокировать движение трансляционного аппарата вниз по транскрипту. [ 33 ] Dom34/Hbs1, вероятно, связывается вблизи сайта A остановленных рибосом и может способствовать рециркуляции комплексов. [ 34 ] В некоторых случаях транскрипт также расщепляется эндонуклеолитическим способом вблизи места стойла; однако личность ответственной эндонуклеазы остается спорной. Фрагментированные . молекулы мРНК затем полностью расщепляются экзосомой по направлению от 3’ до 5’ и Xrn1 по направлению от 5’ до 3’ [ 33 ] В настоящее время неизвестно, как этот процесс высвобождает мРНК из рибосом, однако Hbs1 тесно связан с белком Ski7, который играет явную роль в высвобождении рибосом при Ski7-опосредованном NSD. Предполагается, что Hbs1 может играть аналогичную роль при НГД. [ 5 ] [ 35 ]
Эволюция
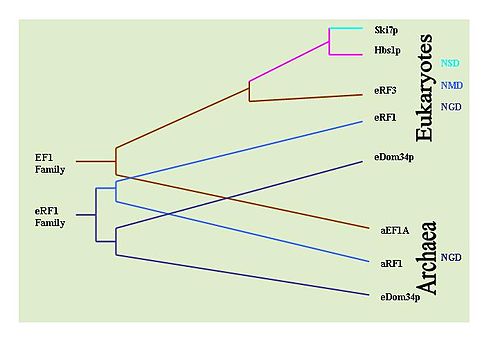
[ редактировать ]Можно определить эволюционную историю этих механизмов, наблюдая за сохранением ключевых белков, участвующих в каждом механизме. Например: Dom34/Hbs1 связаны с NGD; [ 33 ] Ski7 связан с НСД; [ 29 ] а белки eRF связаны с NMD. [ 6 ] С этой целью были проведены обширные поиски BLAST , чтобы определить распространенность белков в различных типах организмов. Установлено, что NGD Hbs1 и NMD eRF3 обнаружены только у эукариот. Однако NGD Dom34 универсален у эукариот и архей . Это указывает на то, что NGD, по-видимому, является первым развитым механизмом наблюдения за мРНК. Белок NSD Ski7, по-видимому, ограничен только видами дрожжей, что позволяет предположить, что NSD является самым последним механизмом наблюдения. Это по умолчанию оставляет НПРО вторым развитым механизмом наблюдения. [ 36 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Амрани Н., Сакс М.С., Джейкобсон А. (июнь 2006 г.). «Ранняя чепуха: распад мРНК решает проблему трансляции». Обзоры природы. Молекулярно-клеточная биология . 7 (6): 415–25. дои : 10.1038/nrm1942 . ПМИД 16723977 . S2CID 32685692 .
- ^ Мур MJ (сентябрь 2005 г.). «От рождения до смерти: сложная жизнь эукариотических мРНК». Наука . 309 (5740): 1514–8. Бибкод : 2005Sci...309.1514M . дои : 10.1126/science.1111443 . ПМИД 16141059 . S2CID 35180502 .
- ^ Jump up to: а б с д и Амрани Н., Ганесан Р., Кервестин С., Мангус Д.А., Гош С., Джейкобсон А. (ноябрь 2004 г.). «Искусственная 3'-UTR способствует аберрантному терминированию и запускает нонсенс-опосредованный распад мРНК» . Природа . 432 (7013): 112–8. Бибкод : 2004Natur.432..112A . дои : 10.1038/nature03060 . ПМИД 15525991 .
- ^ Jump up to: а б с Фаскен М.Б., Корбетт А.Х. (июнь 2005 г.). «Процесс или гибель: контроль качества биогенеза мРНК». Структурная и молекулярная биология природы . 12 (6): 482–8. дои : 10.1038/nsmb945 . ПМИД 15933735 . S2CID 28033127 .
- ^ Jump up to: а б с д и ж г Чанг Ю.Ф., Имам Дж.С., Уилкинсон М.Ф. (2007). «Путь наблюдения за распадом РНК, опосредованный нонсенсом». Ежегодный обзор биохимии . 76 : 51–74. doi : 10.1146/annurev.biochem.76.050106.093909 . ПМИД 17352659 .
- ^ Jump up to: а б с Ревинкель Дж., Раес Дж., Изаурральде Э. (ноябрь 2006 г.). «Нонсенс-опосредованный распад мРНК: гены-мишени и функциональная диверсификация эффекторов». Тенденции биохимических наук . 31 (11): 639–46. дои : 10.1016/j.tibs.2006.09.005 . ПМИД 17010613 .
- ^ Jump up to: а б с Макват Л.Е. (февраль 2004 г.). «Нонсенс-опосредованный распад мРНК: сплайсинг, трансляция и динамика мРНП». Обзоры природы. Молекулярно-клеточная биология . 5 (2): 89–99. дои : 10.1038/nrm1310 . ПМИД 15040442 . S2CID 29469668 .
- ^ Холбрук Дж.А., Ной-Йилик Г., Хентце М.В., Кулозик А.Е. (август 2004 г.). «Бессмысленный распад приближается к клинике». Природная генетика . 36 (8): 801–8. дои : 10.1038/ng1403 . ПМИД 15284851 . S2CID 23188275 .
- ^ Менделл Дж.Т., Шарифи Н.А., Мейерс Дж.Л., Мартинес-Мурильо Ф., Дитц ХК (октябрь 2004 г.). «Бессмысленный надзор регулирует экспрессию различных классов транскриптов млекопитающих и подавляет геномный шум» . Природная генетика . 36 (10): 1073–8. дои : 10.1038/ng1429 . ПМИД 15448691 .
- ^ Лежен Ф., Макват Л.Е. (июнь 2005 г.). «Механистические связи между нонсенс-опосредованным распадом мРНК и сплайсингом пре-мРНК в клетках млекопитающих». Современное мнение в области клеточной биологии . 17 (3): 309–15. дои : 10.1016/j.ceb.2005.03.002 . ПМИД 15901502 .
- ^ Jump up to: а б с Конти Э., Изаурральде Э (июнь 2005 г.). «Бессмысленный распад мРНК: молекулярные знания и механические вариации у разных видов». Современное мнение в области клеточной биологии . 17 (3): 316–25. дои : 10.1016/j.ceb.2005.04.005 . ПМИД 15901503 .
- ^ Jump up to: а б Кали Б.М., Кучма С.Л., Лэтэм Дж., Андерсон П. (февраль 1999 г.). «smg-7 необходим для наблюдения за мРНК у Caenorhabditis elegans» . Генетика . 151 (2): 605–16. дои : 10.1093/генетика/151.2.605 . ПМК 1460488 . ПМИД 9927455 .
- ^ Ямасита А., Касима И., Оно С. (декабрь 2005 г.). «Роль SMG-1 в нонсенс-опосредованном распаде мРНК». Biochimica et Biophysical Acta (BBA) - Белки и протеомика . 1754 (1–2): 305–15. дои : 10.1016/j.bbapap.2005.10.002 . ПМИД 16289965 .
- ^ Jump up to: а б Ким Ю.К., Фурик Л., Дегросейлерс Л., Макват Л.Е. (январь 2005 г.). «У млекопитающих Staufen1 рекрутирует Upf1 на специфические 3'UTR мРНК, чтобы вызвать распад мРНК» . Клетка . 120 (2): 195–208. дои : 10.1016/j.cell.2004.11.050 . ПМИД 15680326 .
- ^ Лонгман Д., Пластерк Р.Х., Джонстон И.Л., Касерес Дж.Ф. (май 2007 г.). «Механистическое понимание и идентификация двух новых факторов пути NMD C. elegans» . Гены и развитие . 21 (9): 1075–85. дои : 10.1101/gad.417707 . ПМЦ 1855233 . ПМИД 17437990 .
- ^ Гатфилд Д., Унтерхольцнер Л., Чиссарелли Ф.Д., Борк П., Изаурральде Э. (август 2003 г.). «Нонсенс-опосредованный распад мРНК у дрозофилы: на пересечении путей дрожжей и млекопитающих» . Журнал ЭМБО . 22 (15): 3960–70. дои : 10.1093/emboj/cdg371 . ПМК 169044 . ПМИД 12881430 .
- ^ Jump up to: а б с Надь Э., Макват Л.Е. (июнь 1998 г.). «Правило положения терминирующего кодона в генах, содержащих интрон: когда нонсенс влияет на обилие РНК». Тенденции биохимических наук . 23 (6): 198–9. дои : 10.1016/S0968-0004(98)01208-0 . ПМИД 9644970 .
- ^ Jump up to: а б с Игрок TJ, Mills DJ, Horton AA (июнь 1979 г.). «Перекисное окисление липидов микросомальной фракции и экстрагированные микросомальные липиды из гепатом, индуцированных DAB» . Британский журнал рака . 39 (6): 773–8. дои : 10.1128/mcb.18.9.5272 . ПМК 109113 . ПМИД 9710612 .
- ^ Ной-Йилик Г., Геринг Н.Х., Терманн Р., Фреде У., Хентце М.В., Кулозик А.Е. (февраль 2001 г.). «Сплайсинг и образование 3'-конца в определении нонсенс-опосредованного распада компетентных мРНП человеческого бета-глобина» . Журнал ЭМБО . 20 (3): 532–40. дои : 10.1093/emboj/20.3.532 . ПМЦ 133467 . ПМИД 11157759 .
- ^ Jump up to: а б Бем-Ансмант И., Гатфилд Д., Ревинкель Дж., Хильгерс В., Изаурральде Э. (март 2007 г.). «Консервативная роль цитоплазматического поли(А)-связывающего белка 1 (PABPC1) в нонсенс-опосредованном распаде мРНК» . Журнал ЭМБО . 26 (6): 1591–601. дои : 10.1038/sj.emboj.7601588 . ПМЦ 1829367 . ПМИД 17318186 .
- ^ Jump up to: а б Касима И, Ямасита А, Идзуми Н, Катаока Н, Моришита Р, Хосино С, Оно М, Дрейфус Г, Оно С (февраль 2006 г.). «Связывание нового комплекса SMG-1-Upf1-eRF1-eRF3 (SURF) с комплексом соединения экзонов запускает фосфорилирование Upf1 и нонсенс-опосредованный распад мРНК» . Гены и развитие . 20 (3): 355–67. дои : 10.1101/gad.1389006 . ПМК 1361706 . ПМИД 16452507 .
- ^ Паланисвами В., Мораес К.С., Вилуш С.Дж., Вилуш Дж. (май 2006 г.). «Нуклеофосмин избирательно откладывается на мРНК во время полиаденилирования» . Структурная и молекулярная биология природы . 13 (5): 429–35. дои : 10.1038/nsmb1080 . ПМК 2811576 . ПМИД 16604083 .
- ^ Шварц А.М., Комарова Т.В., Скулачев М.В., Зверева А.С., Дорохов И., Атабеков Ю.Г. (декабрь 2006 г.). «Стабильность мРНК растений зависит от длины 3'-нетранслируемой области». Биохимия. Биохимия . 71 (12): 1377–84. дои : 10.1134/s0006297906120145 . ПМИД 17223792 . S2CID 1527080 .
- ^ Ньико Т., Кереньи Ф., Шабадкай Л., Бенкович А.Х., Майор П., Сонколи Б., Мерай З., Барта Е., Ниемец Е., Куфель Дж., Силхави Д. (июль 2013 г.). «Распад мРНК растений, опосредованный нонсенсом, контролируется различными схемами ауторегуляции и может быть индуцирован EJC-подобным комплексом» . Исследования нуклеиновых кислот . 41 (13): 6715–28. дои : 10.1093/нар/gkt366 . ПМЦ 3711448 . ПМИД 23666629 .
- ^ Jump up to: а б Инасиу А., Силва А.Л., Пинту Дж., Джи Икс, Моргадо А., Алмейда Ф., Фаустино П., Лавинья Дж., Либхабер С.А., Ромао Л. (июль 2004 г.). «Нонсенс-мутации в непосредственной близости от инициирующего кодона не могут вызвать полный нонсенс-опосредованный распад мРНК» . Журнал биологической химии . 279 (31): 32170–80. дои : 10.1074/jbc.m405024200 . ПМИД 15161914 .
- ^ Сильва А.Л., Перейра Ф.Дж., Моргадо А., Конг Дж., Мартинс Р., Фаустино П., Либхабер С.А., Ромао Л. (декабрь 2006 г.). «Канонический UPF1-зависимый нонсенс-опосредованный распад мРНК ингибируется в транскриптах, несущих короткую открытую рамку считывания, независимо от контекста последовательности» . РНК . 12 (12): 2160–70. дои : 10.1261/rna.201406 . ПМЦ 1664719 . ПМИД 17077274 .
- ^ Силва А.Л., Рибейру П., Инасиу А., Либхабер С.А., Ромао Л. (март 2008 г.). «Близость поли(А)-связывающего белка к кодону преждевременной терминации ингибирует нонсенс-опосредованный распад мРНК млекопитающих» . РНК . 14 (3): 563–76. дои : 10.1261/rna.815108 . ПМК 2248256 . ПМИД 18230761 .
- ^ Гарно Н.Л., Вилуш Дж., Вилуш CJ (февраль 2007 г.). «Шоссе и обходные пути распада мРНК». Обзоры природы. Молекулярно-клеточная биология . 8 (2): 113–26. дои : 10.1038/nrm2104 . ПМИД 17245413 . S2CID 14107577 .
- ^ Jump up to: а б с д ван Хуф А., Фришмейер П.А., Дитц Х.К., Паркер Р. (март 2002 г.). «Экзосомо-опосредованное распознавание и деградация мРНК, не имеющих терминирующего кодона». Наука . 295 (5563): 2262–4. Бибкод : 2002Sci...295.2262V . дои : 10.1126/science.1067272 . ПМИД 11910110 . S2CID 35345349 .
- ^ Jump up to: а б с д Фришмейер П.А., ван Хоф А., О'Доннелл К., Геррерио А.Л., Паркер Р., Дитц ХК (март 2002 г.). «Механизм надзора за мРНК, который устраняет транскрипты, не имеющие терминирующих кодонов». Наука . 295 (5563): 2258–61. Бибкод : 2002Sci...295.2258F . дои : 10.1126/science.1067338 . ПМИД 11910109 . S2CID 40843312 .
- ^ Темперли Р.Дж., Сенека С.Х., Тонска К., Бартник Э., Биндофф Л.А., Лайтаулерс Р.Н., Хшановска-Лайтаулерс З.М. (сентябрь 2003 г.). «Исследование патогенной микроделеции мтДНК выявило зависимый от трансляции путь распада деаденилирования в митохондриях человека» . Молекулярная генетика человека . 12 (18): 2341–8. дои : 10.1093/hmg/ddg238 . ПМИД 12915481 .
- ^ Карзай А.В., Рош Э.Д., Зауэр РТ (июнь 2000 г.). «Система SsrA-SmpB для мечения белков, направленной деградации и спасения рибосом». Структурная биология природы . 7 (6): 449–55. дои : 10.1038/75843 . ПМИД 10881189 .
- ^ Jump up to: а б с д Дома МК, Паркер Р. (март 2006 г.). «Эндонуклеолитическое расщепление эукариотических мРНК с остановкой элонгации трансляции» . Природа . 440 (7083): 561–4. Бибкод : 2006Natur.440..561D . дои : 10.1038/nature04530 . ПМЦ 1839849 . ПМИД 16554824 .
- ^ Кобаяши К., Кикуно И., Куроха К., Сайто К., Ито К., Ишитани Р., Инада Т., Нуреки О. (октябрь 2010 г.). «Структурная основа наблюдения за мРНК архей Pelota и GTP-связанным комплексом EF1α» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (41): 17575–9. Бибкод : 2010PNAS..10717575K . дои : 10.1073/pnas.1009598107 . ПМЦ 2955123 . ПМИД 20876129 .
- ^ Грайль М., Шайе М., ван Тилберг Х. (март 2008 г.). «Структура дрожжей Dom34: белок, связанный с фактором терминации трансляции Erf1 и участвующий в распаде No-Go» . Журнал биологической химии . 283 (11): 7145–54. дои : 10.1074/jbc.M708224200 . ПМИД 18180287 .
- ^ Jump up to: а б Аткинсон Г.К., Балдауф С.Л., Гаврилюк В. (октябрь 2008 г.). «Эволюция безостановочного, бездействующего и бессмысленного распада мРНК и их компонентов, производных от фактора терминации» . Эволюционная биология BMC . 8 (1): 290. Бибкод : 2008BMCEE...8..290A . дои : 10.1186/1471-2148-8-290 . ПМЦ 2613156 . ПМИД 18947425 .