Икра – это яйца и сперматозоиды , выделяемые или попадающие в воду водными животными . Глагол « нереститься » относится к процессу свободного выброса яиц и спермы в водоем (пресную или морскую); физический акт известен как нерест . Подавляющее большинство водных и амфибийных животных размножаются нерестом. К ним относятся следующие группы:
Кораллы — это живые колонии крошечных водных организмов, а не растений, как их иногда считают. Кораллы, хотя по своей природе кажутся малоподвижными или ботаническими, на самом деле нерестятся, выпуская облака сперматозоидов и яйцеклеток в толщу воды, где они смешиваются.
Говорят также, что грибы ) «нерестятся» , ( грибы когда они выделяют белое «волокнистое» вещество, образующее матрицу, из которой они растут. [ нужна ссылка ]
Существует много вариаций в способе нереста, в зависимости от половых различий в анатомии, того, как представители пола относятся друг к другу, где и как выпускается икра, а также от того, охраняется ли она впоследствии и каким образом.
Морские животные, и особенно костистые рыбы , обычно размножаются путем нереста . Это внешний метод размножения, при котором самка выпускает в воду множество неоплодотворенных яиц. В то же время самец или несколько самцов выделяют в воду много спермы, которая оплодотворяет некоторые из этих яиц. Яйца содержат каплю питательного масла, поддерживающего развитие эмбриона внутри яичной оболочки. Масло также обеспечивает плавучесть, поэтому яйца плавают и дрейфуют по течению. Стратегия выживания при массовом нересте состоит в том, чтобы расселить оплодотворенную икру, желательно подальше от побережья, в относительную безопасность открытого океана. Там личинки развиваются по мере потребления своих жировых запасов и в конечном итоге вылупляются из яйцевой капсулы в миниатюрные версии своих родителей.Чтобы выжить, они должны сами стать миниатюрными хищниками, питающимися планктоном. Рыбы в конечном итоге сталкиваются с другими представителями своего вида ( сородичами ), где они образуют скопления и учатся ходить в стаи.
Внутренне пол большинства морских животных можно определить, посмотрев на гонады . Например, семенники самцов нерестящихся рыб гладкие, белые и составляют до 12% массы рыбы, а яичники самок зернистые, оранжевого или желтого цвета и составляют до 70% массы рыбы. Самцы миног, миксин и лосося выделяют сперму в полость тела, откуда она выбрасывается через поры в брюшной полости. Самцы акул и скатов могут передавать сперму по протоку в семенной пузырек , где они хранят ее некоторое время, прежде чем она будет изгнана, в то время как костистые кости обычно используют отдельные семявыносящие протоки. [4] : 141
Внешне многие морские животные, даже во время нереста, демонстрируют небольшой половой диморфизм (разницу в форме или размере тела) или небольшую разницу в окраске . Там, где виды диморфны, например, акулы или гуппи , у самцов часто имеются интромиттивные органы, похожие на пенис , в форме видоизмененного плавника. [4] : 141
Вид считается семельродящим , если его особи нерестятся только один раз в жизни, и итерородящим, если его особи нерестятся более одного раза. Термин semelparity происходит от латинского semel — один раз и pario — рождать, а итеропарность происходит от itero — повторять и pario — рождать.
Семелпарность иногда называют воспроизводством «большого взрыва», поскольку единственное репродуктивное событие семелородящих организмов обычно бывает большим и фатальным для производителей. [5] Классическим примером семелородящего животного является тихоокеанский лосось , который много лет живет в океане, прежде чем доплыть до пресноводного потока, где он родился, нерестился, а затем погибал. К другим нерестящимся животным относятся подёнки , кальмары , осьминоги , корюшка , мойва и некоторые амфибии. [6] Семелпаритет часто ассоциируется с r-стратегами . Однако большинство рыб и других нерестящихся животных итеративные.
Когда внутренние яичники или икринные массы рыб и некоторых морских животных созревают для нереста, их называют икрой . Икра некоторых видов, таких как креветки , морские гребешки , крабы и морские ежи , считается деликатесом во многих частях мира. Икра — это название обработанной соленой икры неоплодотворенной осетровой рыбы . Термин «мягкая икра» или «белая икра» обозначает молоку рыбы . Икру лобстера называют коралловой , потому что при приготовлении она становится ярко-красной. Икру (репродуктивные органы) обычно едят сырой или недолго приготовленной.
«Репродуктивное поведение рыб чрезвычайно разнообразно: они могут быть яйцекладущими (откладывают икру), яйцеживородящими (сохраняют икру в теле до момента вылупления) или живородящими (имеют непосредственную тканевую связь с развивающимися зародышами и рождают живых детенышей). Все хрящевые рыбы — пластиножаберные (например, акулы, скаты и скаты) — используют внутреннее оплодотворение и обычно откладывают крупную икру с толстой скорлупой или рождают живую молодь. скопление полиандрических (многих самцов) гнездящихся скоплений в открытой воде и отсутствие родительской заботы..." [7]
У рыб существует два основных способа размножения. Первый метод - откладка яиц, второй - живорождение (рождение живого молодняка).
При первом способе самка рыбы откладывает икру либо на морское дно, либо на листья водного растения. Рыба-самец оплодотворяет икру, и затем обе они работают вместе, чтобы защитить икру/детенышей от опасности, пока они не смогут защитить себя.
Во втором методе самец рыбы использует свой анальный плавник для передачи спермы самке и оплодотворения икры. Позже самка рождает живых мальков.
Головорезная форель – моногамные парные производители. Удильщик Haplophryne mollis полиандричен. Эта самка тащит за собой атрофированные останки самцов, с которыми она столкнулась.
Моногамия возникает, когда один самец спаривается исключительно с одной самкой. Это еще называют парным нерестом . [9] Большинство рыб не моногамны, а если и моногамны, то часто чередуют немоногамное поведение. Моногамия может возникнуть при небольших размерах кормовых и нерестилищ, когда рыбам трудно найти партнеров или когда за молодняком ухаживают оба пола. [8] Многие тропические цихлиды , которые вместе выращивают детенышей в местах, где им приходится яростно защищаться от конкурентов и хищников, моногамны. [10] «У некоторых иглобрюхих и морских коньков развитие икры занимает много времени, прежде чем самка сможет поместить ее в выводковую сумку самца, где они оплодотворяются. Пока самец беременен, самка начинает новую порцию икры, которая готовы примерно в то же время, когда самец рождает детенышей от предыдущего спаривания. Такое близкое время развития способствует моногамии, особенно если вероятность встречи с другим потенциальным партнером низка». [8]
Полигиния возникает, когда один самец получает исключительные права на спаривание с несколькими самками. При полигинии крупный заметный самец обычно защищает самок от других самцов или защищает место размножения. [8] Самки выбирают крупных самцов, которые успешно защищают лучшие места размножения, которые самки находят привлекательными. Например, самцы бычка защищают «пещеры» под камнями, пригодные для инкубации эмбрионов.
Еще один способ спаривания самцов с несколькими самками — использование токов. Леки – это места, где собирается множество рыб, а самцы демонстрируются друг другу. На основе этих представлений каждая самка затем выбирает самца, которого она хочет стать своей партнершей. Например, у цихлиды Cyrtocara eucinostomus в озере Малави до 50 000 крупных и красочных самцов встречаются вместе на токе длиной четыре километра. Самки, которые высиживают яйца во рту, сами выбирают, какого самца они хотят оплодотворить. [11]
Полиандрия возникает, когда одна самка получает исключительные права на спаривание с несколькими самцами. Это происходит среди рыб, таких как рыба-клоун , которые меняют пол. Это также может произойти, когда самцы высиживают яйца, но не могут справиться со всей икрой, которую производит самка, например, у некоторых игл . [4] : 161
Самцы некоторых глубоководных удильщиков намного меньше самок. Когда они находят самку, они вгрызаются в ее кожу, выделяя фермент , который переваривает кожу их рта и ее тела и сплавляет пару до уровня кровеносных сосудов. Затем самец медленно атрофируется , теряя сначала органы пищеварения, затем мозг, сердце и глаза, превращаясь в не что иное, как пару гонад , которые выделяют сперму в ответ на гормоны в кровотоке самки, указывающие на высвобождение яйцеклеток . Это гарантирует, что, когда самка будет готова к нересту, у нее сразу же появится партнер. [12] Таким образом одна самка удильщика может «спариваться» со многими самцами.
Полигинандрия возникает, когда несколько самцов без разбора спариваются с несколькими самками. Эта взаимная распущенность — подход, наиболее часто используемый нерестящимися животными, и, возможно, это «оригинальная система спаривания рыб». [4] : 161 Распространенными примерами являются кормовые рыбы , такие как сельдь , которые образуют огромные спаривающиеся косяки на мелководье. Вода становится молочной от спермы, а дно покрывается миллионами оплодотворенных яйцеклеток. [4] : 161
Маленькие самцы синежаберных солнечных рыб ловят крупных самцов-рогоносцев, применяя кроссовок или спутников . стратегии Женщины -групперы меняют свой пол на мужской, если самца нет. Пара рыб-анемонов охраняет свою актинию . Если самка умирает, в нее вселяется молодой самец, а самец меняет пол.
Альтернативные стратегии самцов, позволяющие маленьким самцам заниматься рогоносцами, могут развиваться у видов, у которых в нересте преобладают крупные и агрессивные самцы. Рогоносство — это вариант полиандрии, который может возникать у подлых производителей (иногда называемых полосатыми производителями ). Скрытный спаунер — это самец, который бросается присоединиться к нерестовой спешке нерестящейся пары. [13] Натиск нереста происходит, когда рыба резко ускоряется, обычно на почти вертикальном наклоне, выпуская гаметы на вершине, после чего следует быстрое возвращение на озеро или морское дно или в скопление рыб. [14] Крадущиеся самцы в ухаживаниях участия не принимают. Например, у лосося и форели самцы-гнезда часто встречаются . Это маленькие серебристые самцы, которые мигрируют вверх по течению вместе со стандартными крупными крючконосыми самцами и нерестятся, пробираясь в красное (нерестовое гнездо), чтобы высвободить сперму одновременно с спаривающейся парой. Такое поведение представляет собой эволюционно стабильную стратегию воспроизводства, поскольку естественный отбор благоприятствует ему, как и «стандартной» стратегии крупных самцов. [15]
Рогоносство встречается у многих видов рыб, в том числе у драконов , рыб-попугаев и губанов на тропических рифах, а также у синежаберной солнечной рыбы в пресной воде. Самцы-кроссовки, которые становятся слишком большими, чтобы их можно было спрятать, фактически становятся самцами-спутниками . Самцы-спутники синежаберной солнечной рыбы имитируют поведение и окраску самок. Они зависают над гнездом, в котором находится пара ухаживающих рыб-лун, и постепенно опускаются, чтобы добраться до пары, когда они нерестятся. Самцам может потребоваться 6 или 7 лет, чтобы они могли выполнять функции родительских самцов, но они могут быть способны выступать в качестве самцов-кроссовок или спутников-сателлитов, когда им всего 2 или 3 года. Более мелкие самцы-спутники и кроссовки могут быть растерзаны более сильными самцами-родителями, но они нерестятся, когда они моложе, и не вкладывают энергию в родительскую заботу. [4] : 161–2 [16]
Гермафродитизм возникает, когда данная особь вида обладает как мужскими, так и женскими репродуктивными органами или может попеременно обладать то одним, то другим. Гермафродитизм часто встречается у беспозвоночных, но редко у позвоночных. Его можно противопоставить гонохоризму , при котором каждая особь вида является либо самцом, либо самкой и остается таковой на протяжении всей своей жизни. Большинство рыб — гонохористы, но известно, что гермафродитизм встречается у 14 семейств костистых рыб. [17]
Обычно гермафродиты являются последовательными , то есть могут менять пол, обычно с женского на мужской ( протогиния ). Это может произойти, если из группы самок удалить доминирующего самца. Самая крупная самка в гареме может за несколько дней сменить пол и заменить доминирующего самца. [17] Он встречается среди рыб коралловых рифов, таких как морские окуни , рыбы-попугаи и губаны . Реже самец переходит в самку ( протандрия ). [4] : 162 Например, большинство губанов являются протогинными гермафродитами в рамках гаремной системы спаривания. [18] [19] Гермафродитизм допускает сложные системы спаривания. Губаны демонстрируют три различные системы спаривания: полигинную, токообразную и беспорядочную. [20] Групповой нерест и парный нерест происходят в системах спаривания. Тип нереста зависит от размера тела самца. [19] Лаброиды обычно нерестятся широко, выделяя большое количество планктонных яиц, которые разносятся приливными течениями; взрослые губаны не взаимодействуют с потомством. [21] Губаны особой подгруппы семейства Labridae , Labrini, не имеют массового нереста.
Реже гермафродиты могут быть синхронными , то есть они одновременно обладают и яичниками, и яичками и могут функционировать как представители любого пола в любой момент времени. Черные деревушки «по очереди выпускают сперму и икру во время нереста. Поскольку такая торговля икрой выгодна обеим особям, деревни обычно моногамны в течение коротких периодов времени - необычная ситуация для рыб». [22] Пол многих рыб не фиксирован, но может меняться в зависимости от физических и социальных изменений среды, в которой обитают рыбы. [23]
Особенно среди рыб гермафродитизм может окупиться в ситуациях, когда у одного пола больше шансов выжить и размножиться, возможно, потому, что он крупнее. [24] Рыбы-анемоны — последовательные гермафродиты, которые рождаются самцами и становятся самками только в зрелом возрасте. Рыбы-анемоны моногамно живут вместе в анемоне , защищенном укусами анемона. Самцам не приходится конкурировать с другими самцами, а самки анемонов обычно крупнее. Когда самка умирает, молодь (самец) рыбы-анемоны приближается, и «резидентный самец затем превращается в самку, и репродуктивные преимущества комбинации крупной самки и маленького самца сохраняются». [25] У других рыб смена пола обратима. Например, если некоторых бычков сгруппировать по полу (самцы или самки), некоторые поменяют пол. [4] : 164 [24]
Унисексуальность возникает, когда вид состоит только из мужчин или женщин. Унисексуальность встречается у некоторых видов рыб и может принимать сложные формы. Squalius alburnoides , гольян, обитающий в нескольких речных бассейнах Португалии и Испании, по-видимому, является видом, состоящим исключительно из самцов. Существование этого вида иллюстрирует потенциальную сложность систем спаривания у рыб. Вид возник как гибрид двух видов и является диплоидным , но не гермафродитным. Могут иметь триплоидные и тетраплоидные формы, в том числе полностью женские формы, размножающиеся преимущественно посредством гибридогенеза . [26]
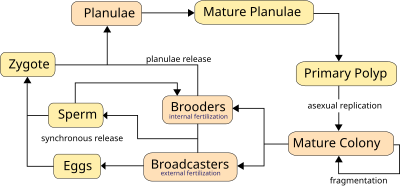
Этот раздел создан по образцу классификации нерестового поведения рыб Бэлона (1975, 1984) по репродуктивным гильдиям . Эта классификация основана на том, как оплодотворяется икра (внутренние или внешние производители), где откладывается икра ( пелагические или донные производители), а также на том, как родители ухаживают за икрой после нереста (носители, охранники и неохранники). [34]
Горечки перекладывают ответственность за заботу о молоди на мидий. Этот горчак-самец демонстрирует нерестовую окраску.
Производители открытого субстрата разбрасывают икру в окружающую среду. Обычно они нерестятся стаями без сложных ритуалов ухаживания, причем самцов больше, чем самок.
Широковещательные производители : выпускают свои гаметы (сперму и яйца) в открытую воду для внешнего оплодотворения. Никакой последующей родительской опеки нет. [35] Около 75% видов кораллов являются трансляторами, большинство из которых являются герматипическими кораллами, или кораллами, строящими рифы. [36]
Пелагические производители : тип широковещательных производителей, нерестятся в открытом море, в основном у поверхности. Обычно это пелагические рыбы, такие как тунец и сардины . Некоторые демерсальные рыбы покидают дно, чтобы нереститься в пелагическом месте, особенно рыбы коралловых рифов , такие как рыбы-попугаи и губаны . Пелагический нерест означает, что водные течения широко рассеивают молодь. Икра, зародыши и личинки пелагических производителей содержат шарики нефти или имеют высокое содержание воды. В результате они плавучие и широко разносятся течениями. Обратной стороной является высокая смертность , поскольку их легко могут съесть пелагические хищники или они могут занестись в неподходящие районы. Самки компенсируют это тем, что откладывают большое количество икры и продлевают периоды нереста. Пелагические производители, обитающие внутри коралловых рифов или вокруг них , могут выметывать небольшое количество икры почти ежедневно в течение нескольких месяцев. Эти рыбы имеют сложное поведение при размножении, включая смену пола , гаремы , токи и территориальность . [4] : 143
Бентические производители : откладывают икру на дне моря (или озера) или вблизи него. Обычно это донные рыбы, такие как треска и камбала . Эти виды обычно нерестятся без церемоний; они не участвуют в сложных ритуалах ухаживания. За каждой самкой обычно следует несколько самцов, которые оплодотворяют икру по мере ее выхода. Различные стратегии гарантируют, что яйца и эмбрионы останутся на месте и не будут дрейфовать по течению. Яйца могут прилипать к другим яйцам или к чему-либо, на что они откладываются, или яйца могут откладываться длинными нитями, обернутыми вокруг растений или камней. Некоторые яйца после того, как их выпустят, впитывают воду, поэтому их можно бросить в трещины, где они разбухнут и застрянут на месте.
Разбрасыватели яиц: разбрасывают клейкие или неклейкие яйца, чтобы они упали на субстрат, на растения или всплыли на поверхность. Эти виды не ухаживают за своим выводком и даже поедают собственные яйца. Часто это стайные рыбы , которые нерестятся группами или парами, часто откладывая большое количество мелкой икры. Мальки вылупляются быстро.
Откладчики яиц: откладывают яйца на подложку (стекло резервуара, дерево, камни, растения). Откладчики яиц обычно откладывают меньше яиц, чем разбрасыватели яиц, хотя яйца крупнее. Откладчики яиц делятся на две группы: те, кто заботится о своих яйцах, и те, кто этого не делает. Среди яйцекладчиков, которые заботятся о своей икре, есть цихлиды и некоторые сомы. Откладчиков яиц, которые заботятся о своем потомстве, можно разделить на две группы: нерестящиеся в полости и нерестящиеся в открытом состоянии.
Полостные производители: откладывают икру в пещеру или полость. Эти рыбы образуют пары и имеют расширенный уход за выводком, при котором икра защищается и очищается. Икра вылупляется через несколько дней, а мальков часто охраняют родители. различные сомы , карповые и киллифиши Большинство составляют . Полостные производители можно противопоставить открытым (укрытым) производителям, которые откладывают икру на открытую поверхность.
Спрятавшие выводок прячут яйца, но не проявляют родительской заботы после того, как спрятали их. Укрыватели выводка - это в основном донные производители, которые закапывают оплодотворенную икру. Например, у лосося и форели самка роет гнездо хвостом в гравии. Эти гнезда называются реддами . Затем самка откладывает икру, пока самец ее оплодотворяет, в то время как обе рыбы при необходимости защищают рыжину от других представителей того же вида. Затем самка закапывает гнездо, и место гнездования покидает. В Северной Америке некоторые гольяны строят гнезда из груд камней, а не роют ямы. У самцов гольяна на голове и теле есть бугорки , которые они используют, чтобы защитить место гнезда. [4] : 145
Яйцекладчики – могут обитать в пересыхающих в определенное время года водах. Примером могут служить однолетние рыбы-киллифиши , откладывающие яйца в грязь. Родители быстро взрослеют и откладывают яйца, прежде чем умереть, когда вода высыхает. Яйца остаются в состоянии покоя, пока дожди не стимулируют вылупление.
Горчаки обладают замечательной стратегией размножения, при которой родители перекладывают ответственность за заботу о потомстве на мидий . Самка выдвигает яйцеклад в мантийную полость мидии и откладывает икру между жаберными нитями. Затем самец выбрасывает свою сперму в вдыхаемый поток воды мидии, и оплодотворение происходит внутри жабр хозяина. Одна и та же самка может использовать несколько мидий, и в каждую она откладывает только одну или две желтые овальные икринки. Ранние стадии развития защищены от хищников внутри тела мидии. Через 3–4 недели личинки уплывают от хозяина, чтобы продолжить жизнь самостоятельно.
Охранники защищают свою икру и потомство после нереста, осуществляя родительскую заботу (также называемую уходом за выводком ). Родительская забота - это «вложение родителей в потомство, которое увеличивает шансы потомства на выживание (и, следовательно, на размножение). У рыб родительская забота может принимать различные формы, включая охрану, строительство гнезда, обмахивание, разбрызгивание, удаление мертвых яиц, поиск заблудившихся мальков, вынашивания яиц снаружи, закапывания яиц, перемещения яиц или молодняка, эктодермального питания, высиживания через рот, внутренней беременности, вынашивания яиц в выводковой сумке и т. д.» [37]
Территориальное поведение обычно необходимо охранникам, и эмбрионы почти всегда охраняются самцами (за исключением цихлид). Необходимо соблюдать территориальность, поскольку уход за эмбрионами обычно включает защиту места, где за ними присматривают. Это также часто означает, что существует конкуренция за лучшие места для откладки яиц. Среди охранников обычно бывает тщательно продуманное ухаживание. [4] : 145
Охраняющие самцы защищают эмбрионы от хищников, поддерживают высокий уровень кислорода, раздувая потоки воды, и защищают территорию от мертвых эмбрионов и мусора. Они защищают эмбрионы до тех пор, пока они не вылупятся, а также часто присматривают за личиночными стадиями. Время, потраченное на охрану, может варьироваться от нескольких дней до нескольких месяцев. [4] : 145
Некоторые охранники строят гнезда ( производители гнезд ), а некоторые нет ( производители субстрата ), хотя разница между этими двумя группами может быть небольшой. [4] : 142 Производители субстрата счищают подходящий участок поверхности, пригодный для откладки яиц, и ухаживают за ним, но активно не строят гнездо.
Ротовые брудеры - несут во рту яйца или личинки. Ротовые брудеры могут быть овофилами или ларвофилами. Овофилы или любящие яйца инкубаторы откладывают яйца в яму, которые засасываются в рот самки. Небольшое количество крупных икринок вылупляется во рту матери, и мальки остаются там некоторое время. Оплодотворение часто происходит с помощью яйцевых пятен, представляющих собой разноцветные пятна на анальном плавнике самца. Когда самка видит эти пятна, она пытается подобрать яйцевые пятна, но вместо этого получает в рот сперму, которая оплодотворяет икру. Многие цихлиды и некоторые лабиринтовые рыбы являются яйцекладущими особями. Ларвофилы или выводки, любящие личинки, откладывают яйца на субстрат и охраняют их, пока из яиц не вылупятся яйца. После вылупления самка подбирает мальков и держит их во рту. Когда мальки смогут постоять за себя, их отпускают. Некоторые землееды — ларвофилы, вынашивающие рот.
Начало эволюционного процесса живорождения начинается с факультативного (факультативного) внутреннего вынашивания. Этот процесс происходит у некоторых видов яйцекладущих (яйцекладущих) киллифиш , которые нерестятся обычным способом на субстрате, но при этом случайно оплодотворяют икру, которую самка сохраняет и не выметывает. Эти яйца нерестятся позже, обычно не оставляя много времени для эмбрионального развития. [4] : 147
Следующим шагом в эволюции живородства является облигатное (по необходимости) внутреннее вынашивание, при котором самка сохраняет все зародыши. «Однако единственным источником питания для этих эмбрионов является яичный желток, как и в икринках, отложенных извне. Эта ситуация, также называемая яйцеживорождением , характерна для морских скальных рыб и байкальских бычков . Эта стратегия позволяет этим рыбам имеют плодовитость, приближающуюся к плодовитости пелагических рыб при внешнем оплодотворении, но это также позволяет им защищать молодь на наиболее уязвимой стадии развития, напротив, акулы и скаты, использующие эту стратегию, производят относительно небольшое количество эмбрионов и сохраняют их в течение некоторого времени. от недель до 16 месяцев и более. Более короткие промежутки времени характерны для видов, которые в конечном итоге откладывают свои эмбрионы в окружающую среду, окруженные роговой капсулой, тогда как более длительные периоды характерны для акул, которые сохраняют эмбрионы до тех пор, пока они не будут готовы к активному выходу на поверхность; плавание молодым». [4] : 147 [38]
Однако некоторые рыбы не подходят под эти категории. живородящая гамбузия крупнородничная ( Gambusia geiseri Считалось, что ) является яйцеживородящей , пока в 2001 году не было показано, что эмбрионы получают питательные вещества от матери. [39]
Нерестилища — это участки воды, где водные животные нерестятся или откладывают икру. После нереста икра может переместиться, а может и не переместиться на новые места, которые станут их нагулом. Многие виды ежегодно совершают миграции , а иногда и большие миграции, чтобы добраться до своих нерестилищ. Например, озера и речные водоразделы могут быть основными нерестилищами анадромных рыб, таких как лосось . В наши дни часто необходимо соорудить рыбные лестницы и другие обводные системы, чтобы лосось мог пройти мимо плотин гидроэлектростанций или других препятствий, таких как плотины, на пути к нерестилищам. [40] [41] Прибрежные рыбы часто используют мангровые заросли и устья рек в качестве нерестилищ, тогда как рифовые рыбы могут найти прилегающие луга с водорослями , которые являются хорошими нерестилищами. Короткоплавники угри могут преодолевать расстояние до трех-четырех тысяч километров к месту своего нереста на глубокой воде где-нибудь в Коралловом море .
Кормовые рыбы часто совершают большие миграции между местами нереста, нагула и нагула. Школы определенного типа обычно путешествуют в треугольнике между этими территориями. Например, нерестилища одного поголовья сельди находятся на юге Норвегии , места нагула — в Исландии , а места нагула — в северной Норвегии. Подобные широкие треугольные путешествия могут быть важны, поскольку кормовая рыба во время кормления не может различить собственное потомство. [42]
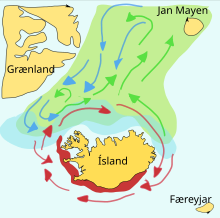
Мойва – кормовая рыба семейства корюшка , обитающая в Атлантическом и Северном Ледовитом океанах. Летом они пасутся на густых стаях планктона на краю шельфового ледника. Более крупная мойва также поедает криль и других ракообразных . Мойва большими стаями перемещается к берегу для нереста и миграции весной и летом, чтобы накормиться в богатых планктоном районах между Исландией , Гренландией и Ян-Майеном . На миграцию влияют океанские течения . Вокруг Исландии созревающая мойва весной и летом совершает крупные кормовые миграции на север. Обратная миграция происходит в сентябре-ноябре. Нерестовая миграция начинается к северу от Исландии в декабре или январе. [43]
На схеме справа показаны основные нерестилища и пути перемещения личинок . Мойва на пути к местам нагула окрашена в зеленый цвет, мойва на обратном пути – в синий, а места размножения – в красный. [43] В статье, опубликованной в 2009 году, исследователи из Исландии рассказывают о своем применении модели взаимодействующих частиц к запасам мойвы вокруг Исландии, успешно предсказав путь нерестовой миграции на 2008 год. [44]
Названный «величайшим косяком на земле», забег сардин происходит, когда миллионы сардин мигрируют со своих нерестилищ к югу от южной оконечности Африки на север вдоль береговой линии Восточного Кейпа. Чавыча совершает самую длинную пресноводную миграцию из всех лососей: более 3000 километров (1900 миль) вверх по реке Юкон к нерестилищам выше по течению от Уайтхорса , Юкон. Некоторые зеленые морские черепахи проплывают более 2600 километров (1600 миль), чтобы добраться до мест нереста.
Самки золотых рыбок откладывают (выбрасывают) икру в воду, поощряемые самцами золотой рыбки, которые одновременно выделяют сперму, которая оплодотворяет икру извне. В течение двух-трех дней из уязвимых икринок золотой рыбки вылупляются личинки , из которых быстро развиваются мальки.
Золотые рыбки , как и все карповые , являются яйцекладущими. Обычно они приступают к размножению после значительного изменения температуры, часто весной. Самцы преследуют самок, побуждая их выпустить яйца, ударяя и подталкивая их. Пока самка золотой рыбки выметывает икру, самец остается рядом и оплодотворяет ее. Их яйца клейкие и прикрепляются к водной растительности. Яйца вылупляются в течение 48–72 часов. Примерно через неделю мальки начинают принимать окончательную форму, хотя может пройти год, прежде чем они приобретут зрелый цвет золотой рыбки; до тех пор они имеют металлически-коричневый цвет, как и их дикие предки. В первые недели жизни мальки растут быстро — адаптация, вызванная высоким риском быть съеденными взрослой золотой рыбкой.
Карп обычно нерестится весной и летом, в зависимости от климата и условий. Уровень кислорода в воде, наличие пищи, размер каждой рыбы, возраст, количество раз, когда рыба нерестилась раньше, и температура воды — все это факторы, которые, как известно, влияют на то, когда и сколько икринок каждый карп будет нерестить одновременно. [45]
Перед нерестом самцы сиамских бойцовских рыб строят пузырьковые гнезда на поверхности воды разного размера. Когда самец начинает интересоваться самкой, он раздувает жабры, крутит тело и расправляет плавники. Самка темнеет и изгибает тело вперед и назад. Акт нереста происходит в «брачных объятиях», когда самец обвивает свое тело вокруг самки, каждое объятие приводит к высвобождению 10–40 яиц, пока у самки не закончатся яйца. Самец со своей стороны выпускает молоки в воду и оплодотворение происходит снаружи. Во время и после нереста самец ртом извлекает тонущую икру и откладывает ее в пузырьковое гнездо (во время спаривания самка иногда помогает партнеру, но чаще она просто пожирает всю икру, которую ей удается поймать). Как только самка выпустит все яйца, ее прогоняют с территории самца, так как вполне вероятно, что она съест яйца из-за голода. [46] Затем яйца остаются на попечении самца. Он держит их в пузырьковом гнезде, следя за тем, чтобы ни один из них не упал на дно, и при необходимости ремонтирует гнездо. Инкубация длится 24–36 часов, а только что вылупившиеся личинки остаются в гнезде следующие 2–3 дня, пока их желточные мешки полностью не рассасываются. После этого мальки покидают гнездо и наступает стадия свободного плавания. [47]
Веслоногие ракообразные — это крошечные ракообразные , которые обычно размножаются либо разбросанным нерестом , либо нерестом в мешочках . Вещательные копеподы разбрасывают икру в воду, а производители мешков откладывают икру в яйцевидный мешок. Производители мешков выметывают небольшое количество, но относительно крупных икринок, которые развиваются медленно. Напротив, широковещательные производители выметывают множество маленьких икринок, которые быстро развиваются. [48] Однако более короткое время вылупления, возникающее в результате разброса, недостаточно, чтобы компенсировать более высокую смертность по сравнению с производителями мешков. Чтобы произвести заданное количество вылупившихся яиц, разбрасыватели должны выметать больше яиц, чем производители мешков. [49]
После спаривания оплодотворенные яйца калифорнийского лангуста самки переносятся на плеоподах до тех пор, пока они не вылупятся, причем одна самка переносит от 120 000 до 680 000 икринок. [50] Яйца начинаются с кораллово-красного цвета, но по мере развития становятся темно-бордовыми. [51] Когда она вынашивает яйца, о самке говорят, что она «ягодная». Яйца готовы к вылуплению через 10 недель. [50] нерест происходит с мая по август. [52] личинками филлосомы Вылупившиеся личинки (называемые . ) не похожи на взрослых особей Вместо этого это плоские, прозрачные животные длиной около 14 мм (0,55 дюйма), но тонкие, как лист бумаги. [53] Личинки питаются планктоном , [51] и вырастают через десять линек до десяти дальнейших личиночных стадий, последняя из которых имеет длину около 30–32 мм (1,2–1,3 дюйма). [53] Полная серия личиночной линьки занимает около 7 месяцев, и когда линяет последняя стадия, она превращается в пуэрулюсное состояние, которое представляет собой ювенильную форму взрослой особи, хотя и все еще прозрачную. [53] Личинки puerulus оседают на морское дно, когда температура воды приближается к максимальной, что в Нижней Калифорнии приходится на осень. [54]
омаров, несущих икру, Самки мигрируют к берегу из более глубоких вод, чтобы высидеть икру, хотя у них нет определенных нерестилищ. [55] [56] Эти миграции омаров могут происходить в виде тесных одиночных группировок «поездов омаров».
Устрицы являются распространителями нереста , то есть икра и сперма выпускаются в открытую воду, где происходит оплодотворение. Они протандрические ; в течение первого года жизни они нерестятся как самцы, выпуская сперму в воду. По мере того, как в течение следующих двух-трех лет они растут и развивают большие запасы энергии, они нерестятся как самки, выпуская яйца . Нерестятся заливные устрицы обычно к концу июня. Повышение температуры воды приводит к нересту нескольких устриц. Это вызывает нерест у остальных, замутняя воду миллионами яиц и сперматозоидов. Одна самка устрицы может производить до 100 миллионов яиц в год. Яйца оплодотворяются в воде и быстро развиваются в планктонных личинок . которые в конечном итоге находят подходящие места, например, раковину другой устрицы, на которых могут поселиться. Прикрепившиеся личинки устриц называются спатом . Спат — это устрицы длиной менее 25 миллиметров (0,98 дюйма).
Тихоокеанская устрица обычно раздельнополая. Их пол можно определить путем исследования гонад , и он может меняться из года в год, обычно в зимние месяцы. В определенных условиях окружающей среды один пол имеет преимущество перед другим. Протандрия предпочтительна в районах с высоким изобилием пищи, а протогиния возникает в областях с низким изобилием пищи. В местах обитания с большим запасом пищи соотношение полов во взрослом населении имеет тенденцию отдавать предпочтение самкам, а в районах с низким изобилием пищи, как правило, наблюдается большая доля взрослых самцов. Нерест тихоокеанских устриц происходит при температуре 20 ° C (68 ° F). Этот вид очень плодовит : самки выпускают около 50–200 миллионов икринок через равные промежутки времени (со скоростью 5–10 раз в минуту) за один нерест. После выхода из гонад яйца проходят через наджаберные камеры ( жабры ), затем через жаберные отверстия попадают в мантийную камеру и, наконец, выбрасываются в воду, образуя небольшое облако. У самцов сперма выделяется на противоположном конце устрицы вместе с нормальной выдыхаемой струей воды. [57] Считается, что повышение температуры воды является основным сигналом к началу нереста, поскольку повышение температуры воды летом приводит к более раннему нересту тихоокеанских устриц. [58]
Личинки тихоокеанской устрицы планктотрофны , их размер на стадии продиссоконха 1 составляет около 70 мкм. Личинки перемещаются по толще воды с помощью личиночной стопы, чтобы найти подходящие места для поселения. На этой фазе они могут провести несколько недель, что зависит от температуры воды, солености и наличия пищи. За эти недели личинки могут рассеиваться на большие расстояния с помощью потоков воды, прежде чем они метаморфизируются и оседают в виде небольших брызг. Как и у других видов устриц, как только личинки тихоокеанских устриц находят подходящую среду обитания, они прикрепляются к ней навсегда, используя цемент, выделяемый железой на их стопе. После расселения личинки превращаются в молодь. Темпы роста очень высоки в оптимальных условиях окружающей среды, а объем рынка может быть достигнут за 18–30 месяцев. [59]
Головоногие моллюски , такие как кальмары и осьминоги, имеют выдающуюся голову и набор рук ( щупалец ), видоизмененных из примитивной стопы моллюсков. Все головоногие моллюски имеют половой диморфизм . Однако у них отсутствуют внешние половые признаки, поэтому они используют цветовое общение. Ухаживающий самец приближается к вероятно выглядящей партнерше, сверкая своими самыми яркими цветами, часто в виде ряби. Если другое головоногие - самка и восприимчива, ее кожа изменит цвет и станет бледной, и произойдет спаривание. Если другое головоногие моллюски остаются ярко окрашенными, это воспринимается как предупреждение. [60]
Все головоногие моллюски размножаются нерестовой икрой . Большинство головоногих моллюсков используют полувнутреннее оплодотворение, при котором самец помещает свои гаметы в мантийную полость самки , чтобы оплодотворить яйцеклетку самки в единственном яичнике . [61] «Пенис» у большинства самцов головоногих моллюсков представляет собой длинный и мускулистый конец гонодода, используемый для переноса сперматофоров в модифицированную несущую сперму руку, называемую гектокотилем . Это, в свою очередь, используется для передачи сперматофоров самке. У видов, у которых гектокотиль отсутствует, «пенис» длинный, может выходить за пределы мантийной полости и переносить сперматофоры непосредственно самке. У многих головоногих спаривание происходит «голова к голове», и самец может просто передать сперму самке. Другие могут отсоединить руку, несущую сперму, и оставить ее прикрепленной к самке. Глубоководные кальмары имеют самую большую известную длину пениса по отношению к размеру тела среди всех подвижных животных, уступая только некоторым сидячим ракушкам во всем животном мире . Удлинение пениса у большого крючковатого кальмара может привести к тому, что длина пениса будет равна длине мантии, головы и рук вместе взятых. [62] [63]
Некоторые виды высиживают оплодотворенные яйца: самки бумажных наутилусов строят убежища для детенышей, а гонатиидные кальмары несут нагруженную личинками оболочку с крючков на своих руках. [64] Другие головоногие откладывают детенышей под камни и проветривают их вылупляющимися щупальцами. В основном яйца предоставлены сами себе; Многие кальмары откладывают яйцеобразные сгустки, похожие на колбаски, в расщелинах или иногда на морском дне. Каракатицы откладывают икру отдельно в чехлы и прикрепляют их к коралловым или водорослевым листьям. [65] Как и тихоокеанский лосось, головоногие моллюски в основном являются семеплавильными , выметывая за одну порцию множество мелких икринок, а затем умирают. Головоногие моллюски обычно живут быстро и умирают молодыми. Большая часть энергии, получаемой из их пищи, используется для роста, и они быстро созревают до взрослого размера. Некоторые набирают до 12% массы тела каждый день. Большинство из них живут один-два года, размножаются, а затем вскоре умирают. [66] [67]
Иглокожие – морские животные, широко распространенные во всех океанах, но не встречающиеся в пресных водах. Прямо под их кожей находится эндоскелет, состоящий из известковых пластинок или косточек .
Морские ежи — это колючие иглокожие со сферическими телами, которые обычно содержат пять гонад . Они передвигаются медленно, питаются в основном морскими водорослями и играют важную роль в рационе каланов . Морские ежи раздельнополы , имеют отдельные мужской и женский пол, хотя, как правило, отличить их нелегко. Гонады покрыты мышцами под брюшиной , что позволяет животному выдавливать свои гаметы через проток в окружающую морскую воду, где происходит оплодотворение. [68] Их икра (мужские и женские гонады) мягкая и тающая, ее цвет варьируется от оранжевого до бледно-желтого, и во многих частях мира ее используют как человеческий деликатес.
Морские огурцы — кожистые иглокожие с удлиненным телом, содержащим одну разветвленную гонаду. Они встречаются на морском дне по всему миру и в большом количестве встречаются на глубоководном морском дне, где они часто составляют большую часть биомассы животных . [69] Они питаются планктоном и разлагающимися органическими отходами, обнаруженными на морском дне, ловя протекающую мимо пищу открытыми щупальцами или просеивая донные отложения . Как и морские ежи, большинство морских огурцов размножаются путем выпуска спермы и яйцеклеток в океанскую воду. В зависимости от условий один организм может производить тысячи гамет .
Морские огурцы обычно раздельнополые , с отдельными мужскими и женскими особями. Репродуктивная система состоит из единственной гонады , состоящей из скопления канальцев, впадающих в один проток, открывающийся на верхней поверхности животного, рядом со щупальцами. [68] Многие виды оплодотворяют яйца внутренне. Оплодотворенная яйцеклетка развивается в сумке на теле взрослой особи и в конечном итоге вылупляется в виде молодого морского огурца. [70] Некоторые виды высиживают детенышей внутри полости тела, рожая через небольшой разрыв в стенке тела рядом с анусом. Остальные виды развивают яйца в свободно плавающую личинку обычно примерно через три дня развития. Эта личинка плавает с помощью длинного пучка ресничек, обернутого вокруг ее тела. По мере роста личинка превращается в бочкообразное тело с тремя-пятью отдельными кольцами ресничек. Щупальца обычно появляются первыми у взрослых особей, прежде чем обычные трубчатые ножки. [68]
Земноводные успешно решили большинство проблем, связанных с воздействием воздуха. Но их репродуктивная система была и остается связана с водой и остается очень похожей на рыбью. Почти все земноводные нерестятся в воде и откладывают большое количество мелких яиц, из которых быстро вылупляются плавающие личинки. Яйца не нуждаются в какой-либо комплексной защите от высыхания, поскольку при высыхании среды личинки обречены так же, как и яйца. Таким образом, отбор способствовал выбору подходящих мест для откладки яиц, а не подходящих устройств для защиты яиц. И рыбы, и земноводные могут мигрировать на большие расстояния в поисках нереста, и предпочтительные места часто активно оспариваются.
Обыкновенные лягушки сортируют свою икру Икра лягушки вблизи Развитие лягушачьей икры
Земноводные обитают в пресноводных озерах и прудах и вокруг них, но не в морской среде. Примерами являются лягушки и жабы, саламандры , тритоны и червяги (напоминающие змей). Это хладнокровные животные, которые превращаются из юной вододышащей формы, обычно во взрослую воздушно-дышащую форму, хотя грязевые щенки сохраняют молодые жабры и во взрослом возрасте.
Самки лягушек и жаб обычно выметывают студенистые яичные массы, содержащие тысячи икринок в воде. Различные виды откладывают яйца разными и узнаваемыми способами. Например, американская жаба откладывает длинные ниточки яиц. Яйца очень уязвимы для хищников , поэтому лягушки разработали множество методов, обеспечивающих выживание следующего поколения. В более холодных регионах эмбрион черный, чтобы поглощать больше солнечного тепла, что ускоряет развитие. Чаще всего это предполагает синхронное воспроизведение . Многие особи будут размножаться одновременно, подавляя действия хищников; большая часть потомства все равно умрет из-за хищников, но есть больше шансов, что некоторые выживут. Еще один способ, с помощью которого некоторые виды избегают хищников и яиц болезнетворных микроорганизмов, которым подвергаются яйца в прудах, - это откладывать яйца на листья над прудом с желеобразным покрытием, предназначенным для удержания влаги. У этих видов головастики после вылупления падают в воду. Яйца некоторых видов, отложенные из воды, могут улавливать вибрации находящихся поблизости хищных ос или змей и вылупляются рано, чтобы их не съели. [72]
Хотя продолжительность стадии яйца зависит от вида и условий окружающей среды, водные яйца обычно вылупляются в течение одной недели. В отличие от саламандр и тритонов, лягушки и жабы никогда не достигают половой зрелости, находясь еще на личиночной стадии. Вылупившиеся яйца продолжают жизнь в виде головастиков , которые обычно имеют овальные тела и длинные, сплюснутые по вертикали хвосты. Как правило, свободноживущие личинки полностью водны. У них отсутствуют веки, они имеют хрящевой скелет, систему боковой линии , жабры для дыхания (сначала наружные, позже внутренние) и хвост с дорсальными и брюшными складками кожи для плавания. [73] У них быстро развивается жаберный мешок, закрывающий жабры и передние ноги; легкие также развиваются на ранней стадии как вспомогательный орган дыхания. У некоторых видов, которые проходят метаморфоз внутри яйца и вылупляются в маленьких лягушек, никогда не развиваются жабры; вместо этого есть специальные участки кожи, которые отвечают за дыхание. У головастиков также отсутствуют настоящие зубы, но челюсти у большинства видов обычно имеют два удлиненных параллельных ряда небольших ороговевших структур, называемых керадонтами, в верхней челюсти, тогда как нижняя челюсть имеет три ряда керадонтов, окруженных роговым клювом , но количество рядов может быть ниже (иногда ноль) или значительно выше. [74] Головастики питаются водорослями , в том числе диатомовыми водорослями , фильтрующимися из воды через жабры . Некоторые виды на стадии головастика плотоядны и поедают насекомых, более мелких головастиков и рыбу. каннибализм Среди головастиков наблюдался . Ранние развивающиеся особи, получившие ноги, могут быть съедены другими, поэтому поздно расцветшие выживут дольше. [75]
Морские черепахи — земноводные рептилии, но не земноводные . Рептилии относятся к классу Рептилии , а земноводные — к классу Амфибии . Это две отдельные таксономические группы. У рептилий чешуя и кожистая кожа, а у амфибий кожа гладкая и пористая. В отличие от лягушек, яйца морских черепах имеют прочную кожистую скорлупу, которая позволяет им выживать на суше, не высыхая.
Некоторые морские черепахи мигрируют на большие расстояния между местами кормления и нереста. Зеленые черепахи имеют места кормления вдоль побережья Бразилии. Каждый год тысячи этих черепах мигрируют на расстояние около 2300 километров (1400 миль) к месту своего нереста, острову Вознесения в Атлантике, острову шириной всего 11 километров (6,8 миль). Каждый год возвращающиеся черепахи выкапывают от 6 000 до 15 000 гнезд, часто возвращаясь на тот же пляж, откуда они вылупились. Самки обычно спариваются каждые два-четыре года. С другой стороны, самцы каждый год посещают места размножения, пытаясь спариваться. [76] Спаривание зеленых морских черепах похоже на спаривание других морских черепах. Самки черепах контролируют процесс. Некоторые популяции практикуют полиандрию , хотя это, похоже, не приносит пользы птенцам. [77] После спаривания в воде самка поднимается над линией прилива на пляже, выкапывает яму задними ластами и откладывает икру. Размер помета зависит от возраста самки и вида, но количество кладок зеленой черепахи колеблется от 100 до 200. Затем она засыпает гнездо песком и возвращается в море. [78]
Примерно через 45–75 дней яйца вылупляются ночью, и птенцы инстинктивно направляются прямо в воду. Это самое опасное время в жизни черепахи. Пока они идут, их хватают хищники, такие как чайки и крабы . Значительный процент никогда не добирается до океана. Мало что известно о первоначальной истории жизни только что вылупившихся морских черепах. [79] Молодые особи проводят от трех до пяти лет в открытом океане, прежде чем еще незрелыми молодыми особями освоятся и начнут вести постоянный образ жизни на мелководье. [80] [81] Предполагается, что им требуется от двадцати до пятидесяти лет, чтобы достичь половой зрелости . Особи живут до восьмидесяти лет в дикой природе. [78] Они относятся к числу самых крупных морских черепах, многие из которых имеют длину более метра и вес до 300 килограммов (660 фунтов). [82]
водные насекомые Нерестятся и . Подёнки «славятся своей короткой взрослой жизнью. У некоторых видов есть меньше часа, чтобы спариваться и откладывать яйца, прежде чем они умрут. Их предвзрослая стадия, известная как субимаго, может быть еще короче — возможно, она длится всего несколько минут, прежде чем они умрут. линяют во взрослую форму, поэтому подёнка проводит большую часть своей жизни в виде нимфы, спрятанной от глаз под водой». [83]
Кораллы могут быть как гонохорическими (однополыми), так и гермафродитными , каждый из которых может размножаться половым и бесполым путем. Размножение также позволяет кораллам заселять новые территории.
Кораллы размножаются преимущественно половым путем . 25% герматипических кораллов (каменистых кораллов) образуют однополые ( гонохористические ) колонии, остальные — гермафродиты . [36] Около 75% всех герматипических кораллов «транслируют икру», выпуская гаметы — яйца и сперму — в воду для распространения потомства. Гаметы сливаются во время оплодотворения, образуя микроскопическую личинку , называемую планулой , обычно розового цвета и эллиптической формы. Типичная колония кораллов образует несколько тысяч личинок в год, чтобы преодолеть препятствия к образованию новой колонии. [84]
Планулы демонстрируют положительный фототаксис , плывя к свету, чтобы достичь поверхностных вод, где они дрейфуют и растут, прежде чем спуститься в поисках твердой поверхности, к которой они могут прикрепиться и основать новую колонию. Они также демонстрируют положительный сонотаксис , двигаясь навстречу звукам, исходящим от рифа, и вдали от открытой воды. [85] Высокая частота неудач затрагивает многие стадии этого процесса, и хотя каждая колония высвобождает миллионы гамет, образуется очень мало новых колоний. Время от нереста до заселения обычно составляет 2–3 дня, но может достигать 2 месяцев. [86] Личинка превращается в полип и в конечном итоге становится коралловой головой в результате бесполого почкования и роста.
Синхронный нерест очень типичен для коралловых рифов, и часто, даже если несколько видов присутствует , все кораллы нерестятся в одну и ту же ночь. Эта синхронность необходима для того, чтобы мужские и женские гаметы могли встретиться. Кораллам приходится полагаться на сигналы окружающей среды, варьирующиеся от вида к виду, чтобы определить подходящее время для выпуска гамет в воду. Сигналы включают лунные изменения, время заката и, возможно, химические сигналы. [36] Синхронный нерест может образовывать гибриды и, возможно, участвует в видообразовании кораллов . [87] В некоторых местах нерест может быть визуально эффектным: обычно прозрачная вода замутняется гаметами, обычно ночью.
Кораллы используют два метода полового размножения, которые различаются тем, выделяются ли женские гаметы:
Вещатели , большинство из которых массово нерестятся, в значительной степени полагаются на сигналы окружающей среды, поскольку они выпускают в воду как сперму, так и яйцеклетки. Кораллы используют долгосрочные сигналы, такие как продолжительность дня , температура воды и/или скорость изменения температуры. Краткосрочным сигналом чаще всего является лунный цикл с закатом, сигнализирующим об освобождении. [36] Около 75% видов кораллов являются трансляторами, большинство из которых являются герматипическими кораллами, или кораллами, строящими рифы. [36] Положительно плавучие гаметы всплывают к поверхности, где в результате оплодотворения образуются личинки планулы . Личинки плывут к поверхности света и попадают в течения, где обычно остаются в течение двух дней, но иногда и до трех недель, а в одном известном случае - два месяца. [86] после чего они расселяются и превращаются в полипы и образуют колонии.
Брудеры чаще всего являются агерматипическими (постройками без рифов) в районах с сильным течением или волнением. Брудеры выделяют только сперму, которая имеет отрицательную плавучесть и может содержать неоплодотворенные икринки в течение нескольких недель, что снижает потребность в массовом синхронном нересте, который иногда случается. [36] После оплодотворения кораллы выпускают личинки планулы , готовые к оседанию. [88]
Сбор урожая вешенки Pleurotus ostreatus , выращенной с использованием икры, помещенной в смесь опилок , помещенную в пластиковые контейнеры.
Грибы не являются растениями, и для оптимального роста им требуются разные условия. Растения развиваются посредством фотосинтеза – процесса, который превращает углекислый газ из атмосферы в углеводы , особенно в целлюлозу . В то время как солнечный свет является источником энергии для растений, грибы получают всю свою энергию и материалы для роста из среды роста посредством биохимического разложения процессов . Это не означает, что свет является ненужным требованием, поскольку некоторые грибы используют свет как сигнал, вызывающий плодоношение. Однако все материалы для роста уже должны присутствовать в питательной среде. Вместо семян грибы размножаются половым путем при подземном росте и бесполым путем спорами . Любой из них может быть заражен переносимыми по воздуху микроорганизмами, которые будут мешать росту грибов и препятствовать получению здорового урожая. Мицелий , или активно растущая грибная культура, помещается на субстрат для посева или внесения грибов для выращивания на субстрате . Это также известно как прививка, нерест или добавление икры. Его основные преимущества заключаются в снижении вероятности заражения и придании грибам твердого начала. [89] [90]
^ "Нерест". Британская энциклопедия. Британская энциклопедия Интернет. Британская энциклопедия, 2011. Интернет. 03 февраля 2011 г. < «Нерест | биология | Британника» . Архивировано из оригинала 2 ноября 2012 г. Проверено 5 февраля 2011 г. >
^ Гросс МР (1982). «Кроссовки, спутники и родители: стратегии полиморфного спаривания североамериканских солнечных рыб». Журнал психологии животных . 60 :1-26. дои : 10.1111/j.1439-0310.1982.tb01073.x .
^ Jump up to: а б Шапиро Д.Ю. (1984) «Реверсия пола и социально-демографические процессы у коралловых рифовых рыб», страницы 103–116 в Г.В. Поттсе и Р.К. Вутуне, ред., Размножение рыб: стратегии и тактика , Academic Press.
^ Робертсон, доктор медицинских наук; Р. Р. Уорнер (1978). «Половые особенности лаброидов Западного Карибского бассейна II: рыбы-попугаи (Scaridae)». Смитсоновский вклад в зоологию . 255 (255): 1–26. дои : 10.5479/si.00810282.255 .
^ Чан СТХ и Юнг ВСБ (1983) «Контроль над сексом и смена пола у рыб в естественных условиях». Страницы 171–222 в WS Hoar, DJ Randall и EM Donaldson, ред., Физиология рыб 9B: Размножение, поведение и контроль фертильности . Академическая пресса.
^ Jump up to: а б Вильялмссон, Х (2002). «Мойва (Mallotus villosus) в экосистеме Исландия – Восточная Гренландия – Ян-Майен». Журнал морских наук ICES . 59 (5): 870–883. дои : 10.1006/jmsc.2002.1233 .
^ Бранч Джордж, Бранч, Марго и Баннистер, Энтони (1981). Живые берега Южной Африки . Кейптаун: К. Струик. ISBN 978-0-86977-115-0 . {{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка )
Arc.Ask3.Ru Номер скриншота №: 2201954d9d793df880264c4d3fee8472__1721276880 URL1:https://arc.ask3.ru/arc/aa/22/72/2201954d9d793df880264c4d3fee8472.html Заголовок, (Title) документа по адресу, URL1: Spawn (biology) - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)












 Детеныши райской рыбки , только что вылупившиеся, собраны под поверхностью пузырькового гнезда.
Детеныши райской рыбки , только что вылупившиеся, собраны под поверхностью пузырькового гнезда. Рыба-анемона гнездится в анемоне . Здесь самец охраняет икру, произведенную партнершей.
Рыба-анемона гнездится в анемоне . Здесь самец охраняет икру, произведенную партнершей.

Аппетитная тилапия — YouTube






 Сиамские бойцовые рыбы строят пузырьковые гнезда разного размера.
Сиамские бойцовые рыбы строят пузырьковые гнезда разного размера. Пара сиамских бойцовских рыб, нерестящихся под своим пузырьковым гнездом.
Пара сиамских бойцовских рыб, нерестящихся под своим пузырьковым гнездом. Однодневные личинки сиамских боевых рыб в пузырьковом гнезде — их желточные мешки еще не рассосались
Однодневные личинки сиамских боевых рыб в пузырьковом гнезде — их желточные мешки еще не рассосались 15-дневный малек сиамской бойцовой рыбы, свободно плавающий.
15-дневный малек сиамской бойцовой рыбы, свободно плавающий.


















 Нерест хрупкой звезды
Нерест хрупкой звезды Голова самки криля с выводковым мешком
Голова самки криля с выводковым мешком Нерест нерки
Нерест нерки Мертвый лосось после нереста
Мертвый лосось после нереста Эти лагуны, соединенные с рекой Тис, обеспечивают тихую заводь для нереста рыбы и убежища во время паводка.
Эти лагуны, соединенные с рекой Тис, обеспечивают тихую заводь для нереста рыбы и убежища во время паводка. В середине этой плотины находится рыбная лестница , которая позволяет форели и лососю пройти через плотину и подняться вверх по реке на нерест.
В середине этой плотины находится рыбная лестница , которая позволяет форели и лососю пройти через плотину и подняться вверх по реке на нерест.







