Клатрин представляет собой белок , который играет важную роль в формировании покрытых оболочкой везикул . Клатрин был впервые выделен Барбарой Пирс в 1976 году. [1] Он образует форму трискелиона , состоящую из трех тяжелых цепей клатрина и трех легких цепей. Когда трискелии взаимодействуют, они образуют многогранную решетку, окружающую везикулу. Название белка относится к этой решетчатой структуре, происходящей от латинского clathri, что означает «решетка». [2] Барбара Пирс назвала белок клатрином по предложению Грэма Митчисона , выбрав его из трёх возможных вариантов. [3] Белки оболочки, такие как клатрин, используются для создания небольших пузырьков для транспортировки молекул внутри клеток. Эндоцитоз . и экзоцитоз везикул позволяют клеткам общаться, переносить питательные вещества, импортировать сигнальные рецепторы, опосредовать иммунный ответ после отбора проб внеклеточного мира и очищать клеточный мусор, оставшийся в результате воспаления тканей Эндоцитарный путь может быть перехвачен вирусами и другими патогенами, чтобы проникнуть в клетку во время инфекции. [4]
Клатрин-трискелион состоит из трех тяжелых цепей клатрина, взаимодействующих на своих С-концах , каждая тяжелая цепь ~190 кДа имеет прочно связанную с ней легкую цепь ~25 кДа. Три тяжелые цепи обеспечивают структурную основу решетки клатрина, а три легкие цепи, как полагают, регулируют образование и разборку решетки клатрина. Существует две формы легких цепей клатрина, обозначенные a и b. Основная тяжелая цепь клатрина, расположенная у человека на 17-й хромосоме , обнаружена во всех клетках. Второй ген тяжелой цепи клатрина, на хромосоме 22 , экспрессируется в мышцах. [5]
Тяжелая цепь клатрина часто описывается как нога с субдоменами, представляющими стопу ( N-концевой домен), за которыми следуют домены лодыжки, дистальной части ноги, колена, проксимальной части ноги и домены тримеризации. N-концевой домен состоит из семилопастной структуры β-пропеллера. Остальные домены образуют суперспираль из коротких альфа-спиралей. Первоначально это было определено на основе структуры домена проксимальной ноги, который идентифицировал и состоит из меньшего структурного модуля, называемого повторяющимися мотивами тяжелой цепи клатрина. Легкие цепи связываются преимущественно с проксимальной частью тяжелой цепи с некоторым взаимодействием вблизи домена тримеризации. β-пропеллер у «ножки» клатрина содержит множество сайтов связывания для взаимодействия с другими белками. [5]
Клатриновая клетка с единственным трискелионом, выделенным синим цветом. Карта CryoEM EMD_5119 была визуализирована в UCSF Chimera, и был выделен один клатриновый трискелион.
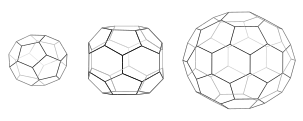
В каждой клетке 12 пятиугольников. Мини-пальто (слева) имеет 4 шестиугольника и тетраэдрическую симметрию, как в усеченном триакис-тетраэдре. Шестиугольный ствол (средний) имеет 8 шестиугольников и симметрию D6. Футбольный мяч (справа) имеет 20 шестиугольников и икосаэдрическую симметрию, как в усеченном икосаэдре.
Когда трискелии собираются вместе в растворе, они могут взаимодействовать с достаточной гибкостью, образуя 6-сторонние кольца ( шестиугольники ), образующие плоскую решетку, или 5-сторонние кольца ( пятиугольники ), необходимые для образования изогнутой решетки. Когда множество трискелионов соединяются, они могут образовывать структуру, похожую на корзину. Показанная структура состоит из 36 трискелий, одна из которых показана синим цветом. Другая распространенная сборка — усеченный икосаэдр . Чтобы вместить везикулу, в решетке должно присутствовать ровно 12 пятиугольников.
В клетке клатрин трискелион в цитоплазме связывается с белком-адаптером, который связал мембрану, одновременно соединяя одну из трех его ножек с мембраной. Клатрин не может напрямую связываться с мембраной или грузом и вместо этого использует для этого адаптерные белки. Этот трискелион связывается с другими трискелиями, прикрепленными к мембране, образуя округлую решетку из шестиугольников и пятиугольников, напоминающую панели футбольного мяча, которые втягивают мембрану в бутон. Путем построения различных комбинаций 5- и 6-сторонних колец можно собирать везикулы разных размеров. Самая маленькая клетка клатрина, которую обычно изображают, называемая мини-пальто, имеет 12 пятиугольников и только два шестиугольника. Еще более мелкие клетки с нулевыми шестиугольниками, вероятно, не образуются из нативного белка, поскольку ножки трискелии слишком громоздки. [6]
Клатрин играет решающую роль в формировании округлых везикул в цитоплазме для внутриклеточного транспорта. Покрытые клатрином везикулы (CCV) избирательно сортируют груз на клеточной мембране , в транс- сети Гольджи и в эндосомальных компартментах для множества путей мембранного транспорта. После того как везикула попадает в цитоплазму, оболочка быстро разбирается, позволяя клатрину перерабатываться, в то время как везикула транспортируется в различные места.
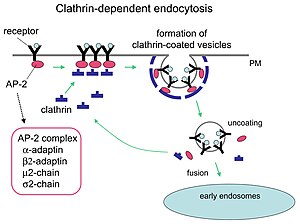
Молекулы-адаптеры отвечают за самосборку и рекрутирование. Двумя примерами адаптерных белков являются AP180. [7] и эпсин . [8] [9] [10] AP180 используется при образовании синаптических пузырьков . Он привлекает клатрин к мембранам, а также способствует его полимеризации . Эпсин также привлекает клатрин к мембранам и способствует его полимеризации, а также может способствовать деформации мембраны, и, таким образом, везикулы, покрытые клатрином, могут отпочковываться. В клетке трискелион, плавающий в цитоплазме, связывается с белком-адаптером, поочередно соединяя одну из его ножек с мембраной. Ножка трискелиона связывается с другими, прикрепленными к мембране, образуя многогранную решетку, ножку трискелиона, которая втягивает мембрану в почку. Ножка не связывается непосредственно с мембраной, а связывается с белками-адаптерами, распознающими молекулы на поверхности мембраны.
Клатрин выполняет еще одну функцию, помимо покрытия органелл . В неделящихся клетках образование везикул, покрытых клатрином, происходит непрерывно. , прекращается образование везикул, покрытых клатрином В клетках, подвергающихся митозу . Во время митоза клатрин связывается с веретенообразным аппаратом в комплексе с двумя другими белками: TACC3 и ch-TOG/ CKAP5 . Клатрин способствует съезду хромосом путем стабилизации кинетохорных волокон митотического веретена . Аминоконцевой домен тяжелой цепи клатрина и домен TACC TACC3 образуют поверхность связывания микротрубочек для TACC3/ch-TOG/клатрина для связывания с митотическим веретеном . Стабилизация кинетохорных волокон требует тримерной структуры клатрина для сшивания микротрубочек. [11] [12] [13]
Клатрин-опосредованный эндоцитоз (CME) регулирует многие клеточные физиологические процессы, такие как интернализация факторов роста и рецепторов, проникновение патогенов и синаптическая передача. Считается, что клеточные захватчики используют питательный путь, чтобы получить доступ к механизмам репликации клетки. Определенные сигнальные молекулы открывают путь питательным веществам. [1] Два химических соединения под названием Pitstop 1 и Pitstop 2, селективные ингибиторы клатрина, могут препятствовать патогенной активности и, таким образом, защищать клетки от инвазии. Эти два соединения избирательно блокируют ассоциацию эндоцитарного лиганда с концевым доменом клатрина in vitro. [14] Однако специфичность этих соединений в блокировании клатрин-опосредованного эндоцитоза подвергается сомнению. [15] Однако в более поздних исследованиях было подтверждено, что специфичность Pitstop 2 зависит от клатрина. [16]
^ МакМахон ХТ. «Галерея Эпсин 1 ЭМ» . МРЦ Лаборатория молекулярной биологии. Архивировано из оригинала 2 января 2009 г. Проверено 17 апреля 2009 г. микрофотографии почкования пузырьков
^ Роль терминального домена клатрина в регуляции динамики покрытых ямок, выявленная с помощью ингибирования малых молекул | Cell, том 146, выпуск 3, 471–484, 5 августа 2011 г. Аннотация. Архивировано 19 января 2012 г. в Wayback Machine.
^ Робертсон М.Дж., Хорачек А., Зауэр С., фон Кляйст Л., Бейкер Дж.Р., Штальшмидт В. и др. (ноябрь 2016 г.). «5-Арил-2-(нафта-1-ил)сульфонамидотиазол-4(5H)-оны как ингибиторы клатрина». Органическая и биомолекулярная химия . 14 (47): 11266–11278. дои : 10.1039/C6OB02308H . ПМИД 27853797 .
Arc.Ask3.Ru Номер скриншота №: dc55668c3848dcdfb95c7797f6da2331__1714814100 URL1:https://arc.ask3.ru/arc/aa/dc/31/dc55668c3848dcdfb95c7797f6da2331.html Заголовок, (Title) документа по адресу, URL1: Clathrin - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)