5-HT3 - рецептор
Эта статья может быть слишком технической для понимания большинства читателей . ( Март 2024 г. ) |
Рецептор 5-HT 3 и, следовательно , принадлежит к (LGIC) Cys-петли суперсемейству лиганд -управляемых ионных каналов отличается структурно и функционально от всех других 5-HT рецепторов (5-гидрокситриптаминовых или серотониновых рецепторов), связанных с G-белком . рецепторы . [1] [2] [3] Этот ионный канал является катион -селективным и обеспечивает деполяризацию и возбуждение нейронов в центральной и периферической нервной системе . [1]
Как и другие лиганд-зависимые ионные каналы, рецептор 5-HT 3 состоит из пяти субъединиц, расположенных вокруг центральной ионпроводящей поры, которая проницаема для ионов натрия (Na), калия (K) и кальция (Ca). Связывание нейромедиатора 5 -гидрокситриптамина ( серотонина ) с рецептором 5-НТ 3 открывает канал, что, в свою очередь, приводит к возбуждающему ответу в нейронах. Быстро активирующий, десенсибилизирующий внутренний ток преимущественно переносится ионами натрия и калия . [2] Рецепторы 5-НТ3 обладают незначительной проницаемостью для анионов . [1] Они наиболее тесно связаны по гомологии с никотиновым ацетилхолиновым рецептором .
Структура
[ редактировать ]Рецептор 5-НТ3 заметно отличается по структуре и механизму от других подтипов рецепторов 5-НТ , которые все связаны с G-белком . Функциональный канал может состоять из пяти идентичных субъединиц 5-HT 3A (гомопентамерных) или смеси 5-HT 3A и одной из четырех других 5-HT 3B . [4] [5] [6] [7] 5-HT 3C , 5-HT 3D или 5-HT 3E (гетеропентамерные). Субъединицы [8] Похоже, что только субъединицы 5-HT 3A образуют функциональные гомопентамерные каналы. Все остальные подтипы субъединиц должны гетеропентамеризоваться с субъединицами 5-HT 3A для образования функциональных каналов. Кроме того, в настоящее время не обнаружено каких-либо фармакологических различий между гетеромерными 5-HT 3AC , 5-HT 3AD , 5-HT 3AE и гомомерными 5-HT 3A рецепторами. [9] N-концевое гликозилирование субъединиц рецептора имеет решающее значение для сборки субъединиц и транспорта через плазматическую мембрану. [10]
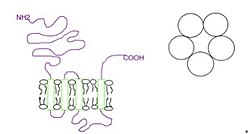
Субъединицы окружают центральный ионный канал псевдосимметрично (рис.1). Каждая субъединица содержит внеклеточный N-концевой домен, который содержит ортостерический сайт связывания лиганда; домен трансмембранный , состоящий из четырех соединенных между собой альфа-спиралей (М1-М4), при этом внеклеточная петля М2-М3 участвует в воротном механизме; большой цитоплазматический домен между М3 и М4, участвующий в транспортировке и регуляции рецепторов; и короткий внеклеточный С-конец (рис.1). [1] В то время как внеклеточный домен является местом действия агонистов и конкурентных антагонистов , трансмембранный домен содержит центральную ионную пору, рецепторные ворота и основной селективный фильтр, который позволяет ионам пересекать клеточную мембрану . [2]
Гены человека и мыши
[ редактировать ]Гены, кодирующие рецепторы 5-HT3 человека , расположены на хромосомах 11 (HTR3A, HTR3B) и 3 (HTR3C, HTR3D, HTR3E), поэтому представляется, что они возникли в результате дупликации генов . Гены HTR3A и HTR3B кодируют субъединицы 5-HT 3A и 5-HT 3B , а HTR3C , HTR3D и HTR3E кодируют субъединицы 5-HT 3C , 5-HT 3D и 5-HT 3E . HTR3C и HTR3E, по-видимому, не образуют функциональные гомомерные каналы, но при совместной экспрессии с HTR3A они образуют гетеромерный комплекс со сниженной или повышенной эффективностью 5-НТ . Патофизиологическая роль этих дополнительных субъединиц еще не определена. [11]
человека Ген рецептора 5- HT3A по структуре подобен гену мыши, который имеет 9 экзонов и занимает площадь ~13 т.п.н. Четыре его интрона находятся точно в том же положении, что и интроны в гомологичном гене рецептора α7-ацетилхолина , что ясно показывает их эволюционное родство. [12] [13]

Выражение . Гены 5-HT 3C , 5-HT 3D и 5-HT 3E имеют тенденцию проявлять периферически ограниченный характер экспрессии с высокими уровнями в кишечнике . в двенадцатиперстной кишке и желудке Например, 5-HT 3C и 5-HT 3E человека количество мРНК может быть больше, чем у 5-HT 3A и 5-HT 3B .
Полиморфизм . У пациентов, получающих химиотерапевтические препараты, определенный полиморфизм гена HTR3B может предсказать успешное противорвотное лечение. Это может указывать на то, что субъединицу рецептора 5-HTR3B можно использовать в качестве биомаркера эффективности противорвотных препаратов.
Распределение тканей
[ редактировать ]Рецептор 5-HT3 экспрессируется в центральной и периферической нервной системе и опосредует множество физиологических функций. [14] На клеточном уровне было показано, что постсинаптические 5-HT3 - рецепторы опосредуют быструю возбуждающую синаптическую передачу в неокортикальных интернейронах крыс, миндалевидном теле и гиппокампе, а также в зрительной коре хорька . [15] [16] [17] [18] Рецепторы 5-HT3 также присутствуют на пресинаптических нервных окончаниях. Имеются некоторые доказательства его роли в модуляции высвобождения нейромедиаторов. [19] [20] но доказательства неубедительны. [21]
Эффекты
[ редактировать ]Когда рецептор активируется для открытия ионного канала агонистами , наблюдаются следующие эффекты:
- ЦНС : центр тошноты и рвоты в стволе мозга , тревога, [22] а также противосудорожное средство [23] и проноцицептивная активность. [24] [25]
- ПНС : возбуждение нейронов (в вегетативных, ноцицептивных нейронах), рвота . [22]
Агонисты
[ редактировать ]Агонисты рецептора включают:
- Цереулида
- 2-метил-5-HT
- Альфа-метилтриптамин
- Буфотенин
- Хлорфенилбигуанид [22]
- Этанол
- Ибогаин
- Фенилбигуанид
- Квипазин
- RS-56812 - Мощный и селективный частичный агонист 5-HT 3 , 1000-кратная селективность по сравнению с другими рецепторами серотонина.
- SR-57227
- Варениклин [26]
- ЯМ-31636 [27]
- С 21007 [28] (SAR см. CGS-12066A )
Антагонисты
[ редактировать ]Антагонисты рецептора (отсортированные по их соответствующему терапевтическому применению) включают:
- противорвотные средства
- Гастропрокинетика
- Алосетрон
- Батаноприд
- Метоклопрамид (высокие дозы)
- Рензаприд
- Закоприд
- М1, основной активный метаболит мосаприда.
- Антидепрессанты
- Нейролептики
- Противомалярийные препараты
- Другие
- 3-тропанил индол-3-карбоксилат
- Каннабидиол (КБД)
- Дельта-9-тетрагидроканнабинол
- Ламотриджин ( эпилепсия и биполярное расстройство )
- Мемантин ( лекарство от болезни Альцгеймера )
- Ментол [29]
- Туйон
Положительные аллостерические модуляторы
[ редактировать ]Эти агенты не являются агонистами рецептора, но увеличивают сродство или эффективность рецепторов к агонисту:
- индола Производные
- 5-хлориндол [30]
- Малые органические анестетики
Открытие
[ редактировать ]Идентификация рецептора 5-HT 3 не состоялась до 1986 года из-за отсутствия селективных фармакологических инструментов. [14] Однако открытие того, что рецептор 5-НТ 3 играет заметную роль в химиотерапией и лучевой терапией , вызванной рвоте , и сопутствующая разработка селективных 5-НТ 3 антагонистов рецептора для подавления этих побочных эффектов вызвали интенсивный интерес со стороны фармацевтической промышленности. [2] [33] и поэтому быстро последовала идентификация рецепторов 5-НТ 3 в клеточных линиях и нативных тканях. [14]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Барнс Н.М., Хейлз Т.Г., Ламмис С.К., Питерс Дж.А. (январь 2009 г.). «Рецептор 5-HT3 — взаимосвязь между структурой и функцией» . Нейрофармакология . 56 (1): 273–284. doi : 10.1016/j.neuropharm.2008.08.003 . ПМК 6485434 . ПМИД 18761359 .
- ^ Jump up to: а б с д Томпсон А.Дж., Ламмис СК (2006). «5-НТ3-рецепторы» . Текущий фармацевтический дизайн . 12 (28): 3615–3630. дои : 10.2174/138161206778522029 . ПМК 2664614 . ПМИД 17073663 .
- ^ Ривз, округ Колумбия, Ламмис, Южная Каролина (2002). «Молекулярные основы структуры и функции рецептора 5-НТ3: модельный лиганд-управляемый ионный канал (обзор)» . Молекулярная мембранная биология . 19 (1): 11–26. дои : 10.1080/09687680110110048 . ПМИД 11989819 . S2CID 36985954 .
- ^ Дэвис П.А., Пистис М., Ханна М.К., Питерс Дж.А., Ламберт Дж.Дж., Хейлз Т.Г., Киркнесс Э.Ф. (1999). «Субъединица 5-HT 3B является основным определяющим фактором функции рецептора серотонина». Природа . 397 (6717): 359–363. Бибкод : 1999Natur.397..359D . дои : 10.1038/16941 . ПМИД 9950429 . S2CID 4401851 .
- ^ Дубин А.Е., Хувар Р., Д'Андреа М.Р., Пяти Дж., Чжу Дж.И., Джой К.С., Уилсон С.Дж., Галиндо Дж.Э., Гласс К.А., Луо Л., Джексон М.Р., Ловенберг Т.В., Эрландер М.Г. (1999). «Фармакологические и функциональные характеристики рецептора серотонина 5-HT 3A специфически модифицируются субъединицей рецептора 5-HT 3B » . J Биол Хим . 274 (43): 30799–30810. дои : 10.1074/jbc.274.43.30799 . ПМИД 10521471 .
- ^ Монк С.А., Десаи К., Брэди К.А., Уильямс Дж.М., Лин Л., Принсивалле А., Хоуп А.Г., Барнс Н.М. (2001). «Получение селективного поликлонального антитела, распознающего субъединицу 5-HT 3B ; идентификация иммунореактивных клеток в гиппокампе крыс». Нейрофармакология . 41 (8): 1013–1016. дои : 10.1016/S0028-3908(01)00153-8 . ПМИД 11747906 . S2CID 10168401 .
- ^ Бойд Г.В., Лоу П., Данлоп Дж.И., Уорд М., Варди А.В., Ламберт Дж.Дж., Питерс Дж., Конолли К.Н. (2002). «Сборка и экспрессия на клеточной поверхности гомомерных и гетеромерных 5-НТ3-рецепторов: роль олигомеризации и белков-шаперонов». Мол клеточные нейроны . 21 (1): 38–50. дои : 10.1006/mcne.2002.1160 . ПМИД 12359150 . S2CID 37832903 .
- ^ Нислер Б., Вальстаб Дж., Комбринк С., Мёллер Д., Капеллер Дж., Ритдорф Дж., Бениш Х., Гётерт М., Раппольд Г., Брюсс М. (2007). «Характеристика новых субъединиц рецептора серотонина человека 5-HT 3C , 5-HT 3D и 5-HT 3E ». Мол Фармакол . 72 (28 марта): 8–17. дои : 10.1124/моль.106.032144 . ПМИД 17392525 . S2CID 40072549 .
- ^ Нислер, Беате (февраль 2011 г.). «Рецепторы 5-HT 3 : потенциал отдельных изоформ для персонализированной терапии». Современное мнение в фармакологии . 11 (1): 81–86. дои : 10.1016/j.coph.2011.01.011 . ПМИД 21345729 .
- ^ Квирк, Филипп Л.; Рао, Сума; Рот, Брайан Л.; Сигел, Рут Э. (15 августа 2004 г.). «Три предполагаемых сайта N-гликозилирования в последовательности мышиного рецептора 5-HT3A влияют на нацеливание на плазматическую мембрану, связывание лиганда и приток кальция в гетерологичные клетки млекопитающих». Журнал нейробиологических исследований . 77 (4): 498–506. дои : 10.1002/мл.20185 . ISSN 0360-4012 . ПМИД 15264219 . S2CID 25811139 .
- ^ Сэнгер Дж.Дж. (сентябрь 2008 г.). «5-гидрокситриптамин и желудочно-кишечный тракт: что дальше?». Тенденции в фармакологических науках . 29 (9): 465–471. дои : 10.1016/j.tips.2008.06.008 . ПМИД 19086255 .
- ^ Jump up to: а б с Утц, П; Абделатти, Ф; Вильярроэль, А; Раппольд, Дж .; Вайс, Б; Коенен, М (1994). «Организация гена мышиного рецептора 5-HT3 и его расположение на хромосоме 11 человека» . Письма ФЭБС . 339 (3): 302–306. Бибкод : 1994FEBSL.339..302U . дои : 10.1016/0014-5793(94)80435-4 . ПМИД 8112471 . S2CID 28979681 .
- ^ Uetz, P. (1992) Ген рецептора 5HT3 мыши. Дипломная работа, Гейдельбергский университет, 143 стр.
- ^ Jump up to: а б с Якель, Дж.Л. (2000). Эндо, М; Курачи, Ю; Мишина, М (ред.). Канал рецептора 5-HT 3 : функция, активация и регуляция в фармакологии Функция ионного канала: активаторы и ингибиторы . Справочник по экспериментальной фармакологии. Том. 147. Берлин: Springer-Verlag . стр. 541–560. ISBN 3-540-66127-1 .
- ^ Ферезу И., Каули Б., Хилл Э.Л., Россье Дж., Хамель Э., Ламболес Б. (2002). «Рецепторы 5-HT 3 опосредуют серотонинергическое быстрое синаптическое возбуждение неокортикальных вазоактивных интестинальных пептидов/холецистокининовых интернейронов» . Дж. Нейроски . 22 (17): 7389–7397. doi : 10.1523/JNEUROSCI.22-17-07389.2002 . ПМК 6757992 . ПМИД 12196560 .
- ^ Казуёси Кава (1994). «Распределение и функциональные свойства рецепторов 5HT 3 в зубчатой извилине гиппокампа крысы». Журнал нейрофизиологии . 71 (5): 1935–1947. дои : 10.1152/янв.1994.71.5.1935 . ПМИД 7520482 .
- ^ Сугита С., Шен К.З., Северная Армения (1992). «5-гидрокситриптамин является быстрым возбуждающим передатчиком рецепторов 5-НТ 3 в миндалевидном теле крысы». Нейрон . 8 (1): 199–203. дои : 10.1016/0896-6273(92)90121-S . ПМИД 1346089 . S2CID 22554779 .
- ^ Рериг Б., Нельсон Д.А., Кац Л.К. (1992). «Быстрая синаптическая передача сигналов никотиновыми рецепторами ацетилхолина и серотонина 5-HT 3 в развивающейся зрительной коре» . Дж. Нейроски . 17 (21): 199–203. doi : 10.1523/JNEUROSCI.17-21-08353.1997 . ПМК 6573745 . ПМИД 9334409 .
- ^ Ронде П., Николс Р.А. (1998). «Высокая кальциевая проницаемость рецепторов серотонина 5-НТ 3 на пресинаптических нервных окончаниях полосатого тела крысы» . Дж. Нейрохем . 70 (3): 1094–1103. дои : 10.1046/j.1471-4159.1998.70031094.x . ПМИД 9489730 .
- ^ Ронде П., Николс Р.А. (1997). «Рецепторы 5-HT 3 вызывают повышение цитозольного и ядерного кальция в клетках NG108-15 посредством индуцированного кальцием высвобождения кальция». Клеточный кальций . 22 (5): 357–365. дои : 10.1016/S0143-4160(97)90020-8 . ПМИД 9448942 .
- ^ ван Хоофт Дж.А., Виджверберг Х.П. (2000). «Рецепторы 5-HT 3 и высвобождение нейромедиаторов в ЦНС: история нервных окончаний?». Тенденции нейробиологии . 23 (12): 605–610. дои : 10.1016/S0166-2236(00)01662-3 . hdl : 1874/7465 . ПМИД 11137150 . S2CID 36074796 .
- ^ Jump up to: а б с д и Ранг, HP (2003). Фармакология . Эдинбург: Черчилль Ливингстон. ISBN 0-443-07145-4 . , стр. 187.
- ^ Голипур Т., Гасеми М., Риази К., Гаффарпур М., Дехпур А.Р. (январь 2010 г.). «Изменение восприимчивости к припадкам через рецептор 5-HT (3): модуляция оксидом азота» . Захват . 19 (1): 17–22. дои : 10.1016/j.seizure.2009.10.006 . ПМИД 19942458 .
- ^ Патель, Райан; Дикенсон, Энтони Х. (сентябрь 2018 г.). «Модально-избирательная роль проноцицептивных спинальных 5-HT2A и 5-HT3-рецепторов в нормальных и нейропатических состояниях» . Нейрофармакология . 143 : 29–37. doi : 10.1016/j.neuropharm.2018.09.028 . ISSN 0028-3908 . ПМК 6277848 . ПМИД 30240783 .
- ^ Сузуки, Ри; Рахман, Вахида; Риг, Ларс Дж; Уэббер, Марк; Хант, Стивен П.; Дикенсон, Энтони Х (октябрь 2005 г.). «Спинно-супраспинальные серотонинергические цепи, регулирующие нейропатическую боль, и ее лечение габапентином». Боль . 117 (3): 292–303. дои : 10.1016/j.pain.2005.06.015 . ISSN 0304-3959 . ПМИД 16150546 . S2CID 6060490 .
- ^ Минёр Ю.С., Пиччотто М.Р. (декабрь 2010 г.). «Никотиновые рецепторы и депрессия: пересмотр и пересмотр холинергической гипотезы» . Тренды Фармакол. Наука . 31 (12): 580–586. дои : 10.1016/j.tips.2010.09.004 . ПМЦ 2991594 . ПМИД 20965579 .
- ^ Иманиши, Н.; Иваока, К.; Косио, Х.; Нагашима, Ю.Ю.; Казута, К.И.; Охта, М.; Сакамото, С.; Ито, Х.; Акузава, С.; Кисо, Т.; Цукамото, СИ; Мейс, Т. (2003). «Новые производные тиазола как мощные и селективные агонисты рецептора 5-гидрокситриптамина 3 (5-HT3) для лечения запоров». Биоорганическая и медицинская химия . 11 (7): 1493–1502. дои : 10.1016/S0968-0896(02)00557-6 . ПМИД 12628674 .
- ^ Делагранж, Филипп; Почетный М.Борис; Мерахи, Насера; Авраам, Кристина; Морен, Филипп; Раулт, Сильвен; Ренар, Пьер; Пфайффер, Бруно; Гвардиола-Леметр, Беатрис; Хамон, Мишель (1996). «Взаимодействие S 21007 с 5-НТ3-рецепторами. Характеристика in vitro и in vivo». Европейский журнал фармакологии . 316 (2–3): 195–203. дои : 10.1016/S0014-2999(96)00680-2 . ISSN 0014-2999 . ПМИД 8982686 .
- ^ Ашур, А.; Нордман, Дж.; Велтри, Д.; Сьюзан Янг, К.-Х.; Шуба Ю.; Аль Кури, Л.; Садек, Б.; Ховарт, ФК; Шеху, А.; Каббани, Н.; Оз, М. (2013). «Ментол ингибирует токи, опосредованные рецептором 5-Ht3». Журнал фармакологии и экспериментальной терапии . 347 (2): 398–409. дои : 10.1124/jpet.113.203976 . ПМИД 23965380 . S2CID 111928 .
- ^ Ньюман, А.С.; Батис, Н.; Графтон, Дж; Капуто, Ф; Брейди, Калифорния; Ламберт, Джей-Джей; Питерс, Дж. А.; Гордон, Дж; Мозг, КЛ; Пауэлл, AD; Барнс, Нью-Мексико (2013). «5-Хлороиндол: мощный аллостерический модулятор рецептора 5-HT3» . Британский журнал фармакологии . 169 (6): 1228–1238. дои : 10.1111/bph.12213 . ПМЦ 3831704 . ПМИД 23594147 .
- ^ Дэвис, Пол А. (2011). «Аллостерическая модуляция рецептора 5-HT3» . Современное мнение в фармакологии . 11 (1). Эльзевир Б.В.: 75–80. дои : 10.1016/j.coph.2011.01.010 . ISSN 1471-4892 . ПМЦ 3072441 . ПМИД 21342788 .
- ^ Jump up to: а б с Солт, Кен; Стивенс, Ренна Дж.; Дэвис, Пол А.; Рейнс, Дуглас Э. (4 августа 2005 г.). «Усиление открытия каналов рецепторов 5-гидрокситриптамина типа 3, индуцированное общим наркозом, зависит от состава субъединиц рецептора». Журнал фармакологии и экспериментальной терапии . 315 (2). Американское общество фармакологии и экспериментальной терапии (ASPET): 771–776. дои : 10.1124/jpet.105.090621 . ISSN 0022-3565 . ПМИД 16081679 . S2CID 22050514 .
- ^ Томпсон А.Дж., Ламмис СК (2007). «Рецептор 5-НТ3 как терапевтическая мишень» . Экспертное мнение. Цели . 11 (4): 527–540. дои : 10.1517/14728222.11.4.527 . ЧВК 1994432 . ПМИД 17373882 .
Внешние ссылки
[ редактировать ]- Рецептор 5-HT3 + в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)