Потенциал действия

Потенциал действия возникает, когда мембранный потенциал конкретной клетки быстро возрастает и падает. [1] Эта деполяризация затем приводит к аналогичной деполяризации соседних участков. Потенциалы действия возникают в нескольких типах возбудимых клеток , включая клетки животных , такие как нейроны и мышечные клетки , а также некоторые растительные клетки . Некоторые эндокринные клетки, такие как бета-клетки поджелудочной железы и некоторые клетки передней доли гипофиза, также являются возбудимыми клетками. [2]
В нейронах потенциалы действия играют центральную роль в межклеточной коммуникации , обеспечивая — или, что касается скачкообразной проводимости нейрона , помогая — распространению сигналов вдоль аксона к синаптическим бутонам , расположенным на концах аксона; эти сигналы могут затем соединяться с другими нейронами в синапсах или с двигательными клетками или железами. В других типах клеток их основная функция — активация внутриклеточных процессов. Например, в мышечных клетках потенциал действия является первым шагом в цепи событий, приводящих к сокращению. В бета-клетках поджелудочной железы они провоцируют выброс инсулина . [а] Потенциалы действия в нейронах также известны как « нервные импульсы » или « спайки », а временная последовательность потенциалов действия, генерируемых нейроном, называется « последовательностью спайков ». Нейрон, который излучает потенциал действия или нервный импульс, часто называют «стреляющим».
Потенциалы действия генерируются особыми типами потенциалзависимых ионных каналов, клетки встроенных в плазматическую мембрану . [б] Эти каналы закрываются, когда мембранный потенциал приближается к (отрицательному) потенциалу покоя клетки, но они быстро начинают открываться, если мембранный потенциал увеличивается до точно определенного порогового напряжения, деполяризуя трансмембранный потенциал. [б] Когда каналы открываются, они пропускают внутрь ионы натрия , что изменяет электрохимический градиент, что, в свою очередь, приводит к дальнейшему повышению мембранного потенциала до нуля. Это приводит к открытию большего количества каналов, создавая больший электрический ток через клеточную мембрану и так далее. Процесс протекает бурно до тех пор, пока не откроются все доступные ионные каналы, что приводит к значительному повышению мембранного потенциала. Быстрый приток ионов натрия вызывает изменение полярности плазматической мембраны, и ионные каналы затем быстро инактивируются. Когда натриевые каналы закрываются, ионы натрия больше не могут проникать в нейрон, а затем активно выводятся обратно из плазматической мембраны. Затем активируются калиевые каналы, и ионы калия выходят наружу, возвращая электрохимический градиент в состояние покоя. После возникновения потенциала действия происходит временный отрицательный сдвиг, называемый постгиперполяризацией .
В животных клетках существует два основных типа потенциалов действия. Один тип генерируется потенциал-управляемыми натриевыми каналами , другой — потенциал-управляемыми кальциевыми каналами . Потенциалы действия на основе натрия обычно длятся менее одной миллисекунды, но потенциалы действия на основе кальция могут длиться 100 миллисекунд или дольше. [ нужна ссылка ] В некоторых типах нейронов медленные выбросы кальция обеспечивают движущую силу для длительного выброса быстро выделяющихся выбросов натрия. С другой стороны, в клетках сердечной мышцы первоначальный быстрый всплеск натрия обеспечивает «праймер», провоцирующий быстрый всплеск кальция, который затем вызывает мышечное сокращение. [3]
Обзор
[ редактировать ]
Почти все клеточные мембраны животных, растений и грибов поддерживают разницу напряжений между внешней и внутренней частью клетки, называемую мембранным потенциалом . Типичное напряжение на мембране клеток животных составляет -70 мВ. Это означает, что внутренняя часть ячейки имеет отрицательное напряжение по отношению к внешней. В большинстве типов клеток мембранный потенциал обычно остается довольно постоянным. Однако некоторые типы клеток электрически активны в том смысле, что их напряжение колеблется с течением времени. В некоторых типах электрически активных клеток, включая нейроны и мышечные клетки, колебания напряжения часто принимают форму быстрого восходящего (положительного) скачка, за которым следует быстрое падение. Эти циклы подъемов и падений известны как потенциалы действия . В некоторых типах нейронов весь цикл подъема и опускания занимает несколько тысячных долей секунды. В мышечных клетках типичный потенциал действия длится около пятой секунды. В растительных клетках потенциал действия может длиться три секунды и более. [4]
Электрические свойства клетки определяются строением ее мембраны. Клеточная мембрана состоит из липидного бислоя молекул, в который встроены более крупные белковые молекулы. Липидный бислой обладает высокой устойчивостью к движению электрически заряженных ионов, поэтому действует как изолятор. Напротив, крупные встроенные в мембрану белки обеспечивают каналы, через которые ионы могут проходить через мембрану. Потенциалы действия управляются белками каналов, конфигурация которых переключается между закрытым и открытым состояниями в зависимости от разницы напряжений внутри и снаружи клетки. Эти чувствительные к напряжению белки известны как потенциалзависимые ионные каналы . [ нужна ссылка ]
Процесс в типичном нейроне
[ редактировать ]
Все клетки в тканях тела животных электрически поляризованы клетки – другими словами, они поддерживают разность потенциалов на плазматической мембране , известную как мембранный потенциал . Эта электрическая поляризация возникает в результате сложного взаимодействия между белковыми структурами, встроенными в мембрану, называемыми ионными насосами и ионными каналами . В нейронах типы ионных каналов в мембране обычно различаются в разных частях клетки, что придает дендритам , аксонам и телу клетки разные электрические свойства. В результате одни части мембраны нейрона могут быть возбудимыми (способными генерировать потенциалы действия), а другие — нет. Недавние исследования показали, что наиболее возбудимой частью нейрона является часть после аксонного бугорка (точка, где аксон выходит из тела клетки), которая называется начальным сегментом аксона , но в большинстве случаев аксон и тело клетки также возбудимы. случаи. [5]
Каждый возбудимый участок мембраны имеет два важных уровня мембранного потенциала: потенциал покоя , который представляет собой значение, которое мембранный потенциал поддерживает до тех пор, пока ничто не возмущает клетку, и более высокое значение, называемое пороговым потенциалом . В аксонном холмике типичного нейрона потенциал покоя составляет около –70 милливольт (мВ), а пороговый потенциал – около –55 мВ. мембраны Синаптические входы в нейрон вызывают деполяризацию или гиперполяризацию ; то есть они вызывают повышение или понижение мембранного потенциала. Потенциалы действия запускаются, когда накапливается достаточная деполяризация, чтобы довести мембранный потенциал до порогового значения. Когда срабатывает потенциал действия, мембранный потенциал резко поднимается вверх, а затем столь же резко возвращается вниз, часто заканчиваясь ниже уровня покоя, где он остается в течение некоторого периода времени. Форма потенциала действия стереотипна; это означает, что подъем и спад обычно имеют примерно одинаковую амплитуду и временной ход для всех потенциалов действия в данной клетке. (Исключения обсуждаются далее в статье). В большинстве нейронов весь процесс происходит примерно за тысячную долю секунды. Многие типы нейронов постоянно излучают потенциалы действия со скоростью до 10–100 в секунду. Однако некоторые типы гораздо тише и могут существовать в течение нескольких минут или дольше, не излучая никаких потенциалов действия.
Биофизическая основа
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( февраль 2014 г. ) |
Потенциалы действия возникают в результате присутствия в клеточной мембране особых типов потенциалзависимых ионных каналов . [6] Потенциал-управляемый ионный канал представляет собой трансмембранный белок, обладающий тремя ключевыми свойствами:
- Он способен принимать более одной конформации.
- По крайней мере, одна из конформаций создает канал через мембрану, проницаемый для определенных типов ионов.
- На переход между конформациями влияет мембранный потенциал.
Таким образом, потенциал-управляемый ионный канал имеет тенденцию быть открытым при одних значениях мембранного потенциала и закрытым при других. Однако в большинстве случаев связь между мембранным потенциалом и состоянием каналов носит вероятностный характер и включает временную задержку. Ионные каналы переключаются между конформациями в непредсказуемое время: мембранный потенциал определяет скорость переходов и вероятность каждого типа перехода в единицу времени.

Потенциал-управляемые ионные каналы способны создавать потенциалы действия, поскольку они могут вызывать петли положительной обратной связи : мембранный потенциал контролирует состояние ионных каналов, но состояние ионных каналов контролирует мембранный потенциал. Таким образом, в некоторых ситуациях повышение мембранного потенциала может привести к открытию ионных каналов, тем самым вызывая дальнейшее повышение мембранного потенциала. Потенциал действия возникает, когда этот цикл положительной обратной связи ( цикл Ходжкина ) протекает взрывно. Временная и амплитудная траектория потенциала действия определяется биофизическими свойствами потенциалзависимых ионных каналов, которые его производят. Действительно существует несколько типов каналов, способных производить положительную обратную связь, необходимую для генерации потенциала действия. Потенциал-управляемые натриевые каналы отвечают за потенциалы быстрого действия, участвующие в нервной проводимости. Более медленные потенциалы действия в мышечных клетках и некоторых типах нейронов генерируются потенциалзависимыми кальциевыми каналами. Каждый из этих типов представлен в нескольких вариантах, с разной чувствительностью к напряжению и разной временной динамикой.
Наиболее интенсивно изучаемый тип потенциалзависимых ионных каналов — натриевые каналы, участвующие в быстрой нервной проводимости. Их иногда называют натриевыми каналами Ходжкина-Хаксли, поскольку они были впервые охарактеризованы Аланом Ходжкиным и Эндрю Хаксли в их исследованиях биофизики потенциала действия, получивших Нобелевскую премию, но их удобнее называть Na V. каналами («V» означает «напряжение».) Канал Na V имеет три возможных состояния: деактивированное , активированное и инактивированное . Канал проницаем для ионов натрия только в активированном состоянии. При низком мембранном потенциале канал большую часть времени находится в деактивированном (закрытом) состоянии. Если мембранный потенциал повышается выше определенного уровня, канал демонстрирует повышенную вероятность перехода в активированное (открытое) состояние. Чем выше мембранный потенциал, тем больше вероятность активации. Как только канал активируется, он в конечном итоге перейдет на инактивированное (закрытое) состояние. Затем он имеет тенденцию оставаться инактивированным в течение некоторого времени, но если мембранный потенциал снова станет низким, канал в конечном итоге вернется в деактивированное состояние. Во время потенциала действия большинство каналов этого типа проходят цикл деактивированный → активированный → инактивированный → деактивированный . Однако это всего лишь среднее поведение населения – отдельный канал в принципе может совершить любой переход в любое время. Однако вероятность перехода канала из инактивированного состояния непосредственно в активированное состояние очень мала: канал в инактивированном состоянии является рефрактерным до тех пор, пока он не перейдет обратно в деактивированное состояние.
Результатом всего этого является то, что кинетика каналов Na V определяется матрицей перехода, скорости которой сложным образом зависят от напряжения. Поскольку эти каналы сами по себе играют важную роль в определении напряжения, глобальную динамику системы может быть довольно сложно определить. Ходжкин и Хаксли подошли к проблеме, разработав систему дифференциальных уравнений для параметров, которые управляют состояниями ионных каналов, известных как уравнения Ходжкина-Хаксли . Эти уравнения были значительно изменены в ходе более поздних исследований, но они составляют отправную точку для большинства теоретических исследований биофизики потенциала действия.
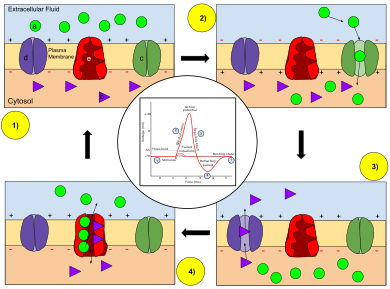
Ключ: а) Натрий (Na + ) ион. б) Калий (К + ) ион. в) Натриевый канал. г) Калиевый канал. д) Натриево-калиевый насос.
На стадиях потенциала действия изменяется проницаемость мембраны нейрона. В состоянии покоя (1) ионы натрия и калия имеют ограниченную способность проходить через мембрану, и внутри нейрона имеется суммарный отрицательный заряд. Как только срабатывает потенциал действия, деполяризация (2) нейрона активирует натриевые каналы, позволяя ионам натрия проходить через клеточную мембрану в клетку, что приводит к образованию чистого положительного заряда в нейроне относительно внеклеточной жидкости. После достижения пика потенциала действия нейрон начинает реполяризацию (3), при которой натриевые каналы закрываются, а калиевые каналы открываются, позволяя ионам калия проникать через мембрану во внеклеточную жидкость, возвращая мембранный потенциал к отрицательному значению. Наконец, наступает рефрактерный период (4), в течение которого потенциалзависимые ионные каналы инактивируются, а Na + и К + ионы возвращаются в состояние покоя, распределяясь по мембране (1), и нейрон готов повторить процесс для следующего потенциала действия.
Когда мембранный потенциал увеличивается, каналы ионов натрия проникать открываются, позволяя ионам натрия в клетку. За этим следует открытие каналов ионов калия выходить , которые позволяют ионам калия из клетки. Внутренний поток ионов натрия увеличивает концентрацию положительно заряженных катионов в клетке и вызывает деполяризацию, при которой потенциал клетки превышает потенциал покоя клетки . Натриевые каналы закрываются на пике потенциала действия, а калий продолжает покидать клетку. Отток ионов калия снижает мембранный потенциал или гиперполяризует клетку. При небольшом увеличении напряжения в состоянии покоя ток калия превышает ток натрия, и напряжение возвращается к своему нормальному значению в состоянии покоя, обычно -70 мВ. [7] [8] [9] Однако, если напряжение превышает критический порог, обычно на 15 мВ выше значения покоя, доминирует натриевый ток. Это приводит к неуправляемому состоянию, при котором положительная обратная связь от натриевого тока активирует еще больше натриевых каналов. Таким образом, клетка срабатывает , создавая потенциал действия. [7] [10] [11] [примечание 1] Частоту, с которой нейрон вызывает потенциалы действия, часто называют скоростью срабатывания или частотой возбуждения нейронов .
Токи, возникающие при открытии потенциалзависимых каналов в ходе потенциала действия, обычно значительно превышают начальный стимулирующий ток. Таким образом, амплитуда, продолжительность и форма потенциала действия во многом определяются свойствами возбудимой мембраны, а не амплитудой или продолжительностью раздражителя. Это свойство потенциала действия «все или ничего» отличает его от градуированных потенциалов , таких как рецепторные потенциалы , электротонические потенциалы , подпороговые колебания мембранного потенциала и синаптические потенциалы , которые масштабируются в зависимости от величины стимула. Во многих типах клеток и клеточных компартментах существуют различные типы потенциалов действия, что определяется типами потенциалзависимых каналов, каналов утечки , распределением каналов, концентрацией ионов, емкостью мембраны, температурой и другими факторами.
Основными ионами, участвующими в потенциале действия, являются катионы натрия и калия; ионы натрия поступают в клетку, а ионы калия выходят, восстанавливая равновесие. Относительно небольшому количеству ионов необходимо пересечь мембрану, чтобы напряжение на мембране резко изменилось. Таким образом, ионы, обменивающиеся во время потенциала действия, вносят незначительные изменения во внутреннюю и внешнюю концентрацию ионов. Те немногие ионы, которые пересекают мембрану, снова откачиваются за счет непрерывного действия натриево-калиевого насоса , который вместе с другими переносчиками ионов поддерживает нормальное соотношение концентраций ионов через мембрану. Катионы кальция и хлорида анионы участвуют в нескольких типах потенциалов действия, таких как сердечный потенциал действия и потенциал действия у одноклеточной водоросли Acetabularia соответственно.
Хотя потенциалы действия генерируются локально на участках возбудимой мембраны, возникающие в результате токи могут запускать потенциалы действия на соседних участках мембраны, ускоряя распространение по принципу домино. В отличие от пассивного распространения электрических потенциалов ( электротонический потенциал ), потенциалы действия заново генерируются вдоль возбудимых участков мембраны и распространяются без распада. [12] Миелиновые участки аксонов не возбудимы и не производят потенциалы действия, а сигнал распространяется пассивно в виде электротонического потенциала . Регулярно расположенные немиелиновые участки, называемые узлами Ранвье , генерируют потенциалы действия для усиления сигнала. Этот тип распространения сигнала, известный как скачкообразная проводимость , обеспечивает благоприятный компромисс между скоростью сигнала и диаметром аксона. Деполяризация окончаний аксонов , как правило, вызывает высвобождение нейромедиатора в синаптическую щель . Кроме того, потенциалы обратного распространения были зарегистрированы в дендритах пирамидных нейронов , которые повсеместно распространены в неокортексе. [с] Считается, что они играют роль в пластичности, зависящей от времени всплеска .
В модели емкости мембраны Ходжкина-Хаксли скорость передачи потенциала действия не была определена, и предполагалось, что соседние области деполяризуются из-за интерференции высвободившихся ионов с соседними каналами. Измерения диффузии и радиусов ионов с тех пор показали, что это невозможно. [ нужна ссылка ] Более того, противоречивые измерения изменений энтропии и времени поставили под сомнение действующую в одиночку модель емкости. [ нужна ссылка ] Альтернативно, гипотеза адсорбции Гилберта Линга утверждает, что мембранный потенциал и потенциал действия живой клетки обусловлены адсорбцией мобильных ионов на участках адсорбции клеток. [13]
Созревание электрических свойств потенциала действия
[ редактировать ]Способность нейрона процессе генерировать и распространять потенциал действия меняется в развития . Насколько изменится мембранный потенциал нейрона под действием импульса тока, зависит от входного сопротивления мембраны . По мере роста клетки к мембране добавляется больше каналов , что приводит к уменьшению входного сопротивления. Зрелый нейрон также претерпевает более короткие изменения мембранного потенциала в ответ на синаптические токи. хорька Нейроны латерального коленчатого ядра имеют более длительную постоянную времени и большее отклонение напряжения при P0, чем при P30. [14] Одним из последствий уменьшения продолжительности потенциала действия является то, что точность сигнала может сохраняться в ответ на высокочастотную стимуляцию. Незрелые нейроны более склонны к синаптической депрессии, чем к потенциации после высокочастотной стимуляции. [14]
На ранних стадиях развития многих организмов потенциал действия изначально переносится кальциевым, а не натриевым током . Кинетика открытия и закрытия кальциевых каналов во время развития медленнее, чем у потенциалзависимых натриевых каналов, которые несут потенциал действия в зрелых нейронах. Более длительное время открытия кальциевых каналов может привести к тому, что потенциалы действия будут значительно медленнее, чем у зрелых нейронов. [14] Нейроны Xenopus изначально имеют потенциалы действия, длительность которых составляет 60–90 мс. В процессе разработки это время уменьшается до 1 мс. Есть две причины такого резкого снижения. Во-первых, входящий ток в основном переносится по натриевым каналам. [15] Во-вторых, выпрямитель с задержкой , ток калиевого канала , увеличивается в 3,5 раза по сравнению с первоначальной силой. [14]
Для того чтобы произошел переход от кальций-зависимого потенциала действия к натрий-зависимому потенциалу действия, к мембране необходимо добавить новые каналы. Если нейроны Xenopus выращивать в среде с ингибиторами синтеза РНК или синтеза белка , этот переход предотвращается. [16] Даже электрическая активность самой клетки может играть роль в экспрессии каналов. Если потенциалы действия в миоцитах Xenopus блокируются, типичное увеличение плотности тока натрия и калия предотвращается или задерживается. [17]
Такое созревание электрических свойств наблюдается у разных видов. Токи натрия и калия Xenopus резко увеличиваются после того, как нейрон проходит заключительную фазу митоза . Плотность натриевого тока корковых нейронов крыс увеличивается на 600% в течение первых двух постнатальных недель. [14]
нейротрансмиссия
[ редактировать ]Анатомия нейрона
[ редактировать ]
Несколько типов клеток поддерживают потенциал действия, например, растительные клетки, мышечные клетки и специализированные клетки сердца (в которых возникает сердечный потенциал действия ). Однако основной возбудимой клеткой является нейрон , который также имеет простейший механизм формирования потенциала действия. [ нужна ссылка ]
Нейроны представляют собой электрически возбудимые клетки, состоящие, как правило, из одного или нескольких дендритов, одной сомы , одного аксона и одного или нескольких окончаний аксона . Дендриты — это клеточные отростки, основная функция которых — получение синаптических сигналов. Их выступы, известные как дендритные шипы , предназначены для захвата нейротрансмиттеров, выделяемых пресинаптическим нейроном. Они имеют высокую концентрацию лиганд-управляемых ионных каналов . Эти шипы имеют тонкую шейку, соединяющую выпуклый выступ с дендритом. Это гарантирует, что изменения, происходящие внутри позвоночника, с меньшей вероятностью затронут соседние позвонки. Дендритный шип может, за редким исключением (см. LTP ), действовать как самостоятельная единица. Дендриты отходят от сомы, в которой находится ядро и многие «нормальные» эукариотические органеллы. В отличие от шипов, поверхность сомы заполнена ионными каналами, активируемыми напряжением. Эти каналы помогают передавать сигналы, генерируемые дендритами. Из сомы выходит аксонный бугорок. . Эта область характеризуется очень высокой концентрацией натриевых каналов, активируемых напряжением. В целом это считается зоной инициации спайков потенциалов действия. [18] то есть триггерная зона . Множественные сигналы, генерируемые в позвоночнике и передаваемые сомой, сходятся здесь. Сразу за аксонным бугорком находится аксон. Это тонкий трубчатый выступ, отходящий от сомы. Аксон изолирован миелиновой оболочкой. Миелин состоит либо из шванновских клеток (в периферической нервной системе), либо из олигодендроцитов (в центральной нервной системе), оба из которых являются типами глиальных клеток . Хотя глиальные клетки не участвуют в передаче электрических сигналов, они взаимодействуют и обеспечивают важную биохимическую поддержку нейронов. [19] Точнее, миелин несколько раз оборачивается вокруг сегмента аксона, образуя толстый жировой слой, который предотвращает вход или выход ионов из аксона. Эта изоляция предотвращает значительное затухание сигнала, а также обеспечивает более высокую скорость передачи сигнала. Однако эта изоляция имеет ограничение: на поверхности аксона не может быть никаких каналов. Таким образом, существуют равномерно расположенные участки мембраны, не имеющие изоляции. Эти узлы Ранвье можно рассматривать как «мини-бугорки аксонов», поскольку их цель — усилить сигнал, чтобы предотвратить его значительное затухание. На самом дальнем конце аксон теряет изоляцию и начинает разветвляться на несколько окончаний аксона . Эти пресинаптические окончания, или синаптические бутоны, представляют собой специализированную область внутри аксона пресинаптической клетки, содержащую нейротрансмиттеры , заключенные в небольшие мембраносвязанные сферы, называемые синаптическими пузырьками . [ нужна ссылка ]
Инициация
[ редактировать ]Прежде чем рассматривать распространение потенциалов действия по аксонам и их прекращение в синаптических бугорках, полезно рассмотреть методы, с помощью которых потенциалы действия могут быть инициированы в аксонном холмике . Основное требование состоит в том, чтобы напряжение мембраны на бугре было выше порога срабатывания. [7] [8] [20] [21] Есть несколько способов, которыми может произойти эта деполяризация.

Динамика
[ редактировать ]Потенциалы действия чаще всего инициируются возбуждающими постсинаптическими потенциалами пресинаптического нейрона. [22] Обычно молекулы нейромедиатора высвобождаются пресинаптическим нейроном . Эти нейротрансмиттеры затем связываются с рецепторами постсинаптической клетки. Это связывание открывает различные типы ионных каналов . Это открытие имеет дополнительный эффект изменения местной проницаемости клеточной мембраны и, следовательно, мембранного потенциала. Если связывание увеличивает напряжение (деполяризует мембрану), синапс является возбуждающим. Однако если связывание снижает напряжение (гиперполяризует мембрану), оно является тормозным. Независимо от того, увеличивается или уменьшается напряжение, изменение пассивно распространяется на близлежащие области мембраны (как описано уравнением кабеля и его уточнениями). Обычно напряжение стимула затухает экспоненциально по мере удаления от синапса и со временем от связывания нейромедиатора. Некоторая часть возбуждающего напряжения может достичь бугорка аксона и (в редких случаях) деполяризовать мембрану настолько, чтобы спровоцировать новый потенциал действия. Чаще всего возбуждающие потенциалы нескольких синапсов должны работают вместе чтобы почти одновременно, спровоцировать новый потенциал действия. Однако их совместным усилиям могут помешать противодействующие тормозные постсинаптические потенциалы . [ нужна ссылка ]
Нейротрансмиссия также может происходить через электрические синапсы . [23] Благодаря прямой связи между возбудимыми клетками в виде щелевых контактов потенциал действия может передаваться непосредственно от одной клетки к другой в любом направлении. Свободный поток ионов между клетками обеспечивает быструю нехимическую передачу. Выпрямляющие каналы обеспечивают движение потенциалов действия только в одном направлении через электрический синапс. [ нужна ссылка ] Электрические синапсы встречаются во всех нервных системах, включая человеческий мозг, хотя их явное меньшинство. [24]
Принцип «все или ничего»
[ редактировать ]потенциала Часто полагают, что амплитуда действия не зависит от силы тока, который его вызвал. Другими словами, большие токи не создают больших потенциалов действия. Поэтому говорят, что потенциалы действия являются сигналами «все или ничего» , поскольку они либо возникают полностью, либо не возникают вообще. [д] [и] [ф] В этом отличие от рецепторных потенциалов , амплитуды которых зависят от интенсивности стимула. [25] В обоих случаях частота потенциалов действия коррелирует с интенсивностью стимула.
Несмотря на то, что классический взгляд на потенциал действия как на стереотипный, однородный сигнал, доминировавший в области нейробиологии на протяжении многих десятилетий, новые данные действительно позволяют предположить, что потенциалы действия — это более сложные события, действительно способные передавать информацию не только благодаря своей амплитуде, но и своей продолжительности и фазу, а иногда даже на расстояния, которые изначально считались невозможными. [26] [27] [28] [29]
Сенсорные нейроны
[ редактировать ]В сенсорных нейронах внешний сигнал, такой как давление, температура, свет или звук, связан с открытием и закрытием ионных каналов , которые, в свою очередь, изменяют ионную проницаемость мембраны и ее напряжение. [30] Эти изменения напряжения снова могут быть возбуждающими (деполяризующими) или тормозящими (гиперполяризующими), и в некоторых сенсорных нейронах их совокупное воздействие может деполяризовать аксонный холмик настолько, что спровоцирует потенциалы действия. Некоторые примеры у людей включают обонятельный рецепторный нейрон и тельце Мейсснера , которые имеют решающее значение для обоняния и осязания соответственно . Однако не все сенсорные нейроны преобразуют свои внешние сигналы в потенциалы действия; у некоторых даже нет аксона. [31] Вместо этого они могут преобразовать сигнал в высвобождение нейромедиатора или в непрерывные градуированные потенциалы , любой из которых может стимулировать последующие нейроны к запуску потенциала действия. Например, в человеческом волосковые ухе клетки преобразуют входящий звук в открытие и закрытие механически управляемых ионных каналов , что может вызвать нейромедиаторов высвобождение молекул . Аналогичным образом, в сетчатке человека начальные фоторецепторные клетки и следующий слой клеток (включающий биполярные клетки и горизонтальные клетки ) не производят потенциалы действия; только некоторые амакриновые клетки и третий слой, ганглиозные клетки , производят потенциалы действия, которые затем перемещаются вверх по зрительному нерву . [ нужна ссылка ]
Потенциалы кардиостимулятора
[ редактировать ]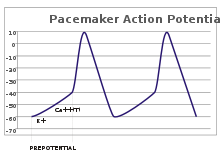
В сенсорных нейронах потенциалы действия возникают в результате внешнего стимула. Однако некоторым возбудимым клеткам такой стимул для срабатывания не требуется: они спонтанно деполяризуют свой аксонный бугорок и запускают потенциалы действия с постоянной скоростью, как внутренние часы. [32] Следы напряжения таких клеток известны как потенциалы кардиостимулятора . [33] синоатриального клетки узла сердца . кардиостимуляторные Хорошим примером являются [г] Хотя такие потенциалы кардиостимулятора имеют естественный ритм , его можно регулировать внешними раздражителями; например, частота сердечных сокращений может быть изменена фармацевтическими препаратами, а также сигналами симпатических и парасимпатических нервов. [34] Внешние стимулы не вызывают повторяющихся срабатываний клетки, а просто изменяют время их срабатывания. [33] В некоторых случаях регулирование частоты может быть более сложным, что приводит к возникновению паттернов потенциалов действия, таких как взрывы . [ нужна ссылка ]
Фазы
[ редактировать ]Ход потенциала действия можно разделить на пять частей: фазу подъема, фазу пика, фазу спада, фазу снижения и рефрактерный период. Во время фазы подъема мембранный потенциал деполяризуется (становится более положительным). Точка, в которой деполяризация прекращается, называется пиковой фазой. На этом этапе мембранный потенциал достигает максимума. После этого наступает фаза падения. На этом этапе мембранный потенциал становится более отрицательным, возвращаясь к потенциалу покоя. Фаза недолета, или послегиперполяризации , — это период, в течение которого мембранный потенциал временно становится более отрицательно заряженным, чем в состоянии покоя (гиперполяризация). Наконец, время, в течение которого последующий потенциал действия невозможно или трудно вызвать, называется рефрактерным периодом и может перекрываться с другими фазами. [35]
Ход потенциала действия определяется двумя связанными эффектами. [36] Во-первых, потенциал-чувствительные ионные каналы открываются и закрываются в ответ на изменения мембранного напряжения V m . Это изменяет проницаемость мембраны для этих ионов. [37] Во-вторых, согласно уравнению Гольдмана проницаемости изменяет равновесный потенциал Em m и, следовательно, мембранное напряжение V . , это изменение [час] Таким образом, мембранный потенциал влияет на проницаемость, которая в дальнейшем влияет на мембранный потенциал. Это создает возможность для положительной обратной связи , которая является ключевой частью фазы роста потенциала действия. [7] [10] Осложняющим фактором является то, что один ионный канал может иметь несколько внутренних «ворот», которые реагируют на изменения V m противоположным образом или с разной скоростью. [38] [я] Например, хотя повышение V m открывает большинство ворот потенциал-чувствительного натриевого канала, оно также закрывает «ворота инактивации» канала, хотя и медленнее. [39] Следовательно, когда V m внезапно повышается, натриевые каналы сначала открываются, но затем закрываются из-за более медленной инактивации.
Напряжения и токи потенциала действия во всех его фазах были точно смоделированы Аланом Ллойдом Ходжкиным и Эндрю Хаксли в 1952 году. [я] за что им была присуждена Нобелевская премия по физиологии и медицине в 1963 году. [нижнегреческий 2] Однако их модель учитывает только два типа потенциал-чувствительных ионных каналов и делает о них несколько предположений, например, что их внутренние ворота открываются и закрываются независимо друг от друга. На самом деле существует множество типов ионных каналов. [40] и они не всегда открываются и закрываются самостоятельно. [Дж]
Фаза стимуляции и подъема
[ редактировать ]Типичный потенциал действия начинается на аксонном холмике. [41] при достаточно сильной деполяризации, например, раздражителе, увеличивающем V m . Эта деполяризация часто вызывается введением дополнительных катионов в клетку натрия; эти катионы могут поступать из самых разных источников, таких как химические синапсы , сенсорные нейроны или потенциалы кардиостимулятора . [ нужна ссылка ]
Для нейрона в состоянии покоя во внеклеточной жидкости наблюдается высокая концентрация ионов натрия и хлора по сравнению с внутриклеточной жидкостью , а во внутриклеточной жидкости - высокая концентрация ионов калия по сравнению с внеклеточной жидкостью. Разница в концентрациях, которая заставляет ионы перемещаться от высокой концентрации к низкой , и электростатические эффекты (притяжение противоположных зарядов) ответственны за движение ионов в нейрон и из него. Внутренняя часть нейрона имеет отрицательный заряд по сравнению с внешней частью клетки из-за движения K. + из клетки. Мембрана нейрона более проницаема для К. + чем к другим ионам, что позволяет этому иону выборочно выходить из клетки по градиенту ее концентрации. Этот градиент концентрации вместе с каналами утечки калия , присутствующими на мембране нейрона, вызывает отток ионов калия, делая потенциал покоя близким к E K ≈ –75 мВ. [42] Поскольку На + ионы находятся в более высоких концентрациях вне клетки, разница в концентрации и напряжении приводит их в клетку, когда Na + каналы открываются. Деполяризация открывает как натриевые, так и калиевые каналы в мембране, позволяя ионам поступать в аксон и выходить из него соответственно. Если деполяризация невелика (скажем, при увеличении V m с -70 мВ до -60 мВ), внешний ток калия подавляет входящий ток натрия, и мембрана реполяризуется обратно к своему нормальному потенциалу покоя около -70 мВ. [7] [8] [9] Однако, если деполяризация достаточно велика, входящий ток натрия увеличивается больше, чем внешний ток калия, и возникает состояние неуправляемого движения ( положительная обратная связь ): чем больше входящий ток, тем больше увеличивается V m , что, в свою очередь, еще больше увеличивает входящий ток. текущий. [7] [10] Достаточно сильная деполяризация (увеличение V м ) вызывает открытие потенциал-чувствительных натриевых каналов; увеличение проницаемости для натрия приближает V m к равновесному напряжению натрия E Na ≈ +55 мВ. Увеличение напряжения, в свою очередь, вызывает открытие еще большего количества натриевых каналов, что подталкивает V m еще больше к E Na . Эта положительная обратная связь продолжается до тех пор, пока натриевые каналы не откроются полностью и V m не приблизится к E Na . [7] [8] [20] [21] Резкое повышение V м и натриевой проницаемости соответствует фазе нарастания потенциала действия. [7] [8] [20] [21]
Критическое пороговое напряжение для этого состояния неконтролируемого состояния обычно составляет около -45 мВ, но оно зависит от недавней активности аксона. Клетка, только что запустившая потенциал действия, не может немедленно запустить другой, поскольку Na + каналы не восстановились из неактивного состояния. Период, в течение которого не может быть запущен новый потенциал действия, называется периодом абсолютной рефрактерности . [43] [44] [45] В более длительные периоды времени, после того как некоторые, но не все ионные каналы восстановились, аксон можно стимулировать для создания другого потенциала действия, но с более высоким порогом, требующим гораздо более сильной деполяризации, например, до -30 мВ. Период, в течение которого потенциалы действия необычайно трудно вызвать, называется относительным рефрактерным периодом . [43] [44] [45]
Пиковая фаза
[ редактировать ]Положительная обратная связь восходящей фазы замедляется и прекращается, когда каналы ионов натрия становятся максимально открытыми. На пике потенциала действия проницаемость натрия максимальна, а мембранное напряжение V m почти равно равновесному напряжению натрия E Na . Однако то же самое повышенное напряжение, которое первоначально открыло натриевые каналы, также медленно их закрывает, закрывая их поры; натриевые каналы инактивируются . [39] Это снижает проницаемость мембраны для натрия по сравнению с калием, возвращая мембранное напряжение к значению покоя. В то же время повышенное напряжение открывает чувствительные к потенциалу калиевые каналы; увеличение проницаемости мембраны для калия приводит к тому, что m приближается к EK V . [39] В совокупности эти изменения проницаемости для натрия и калия вызывают быстрое падение V m , реполяризуя мембрану и вызывая «фазу падения» потенциала действия. [43] [46] [21] [47]
Послегиперполяризации
[ редактировать ]Деполяризованное напряжение открывает дополнительные зависимые от напряжения калиевые каналы, и некоторые из них не закрываются сразу, когда мембрана возвращается к своему нормальному напряжению покоя. Кроме того, дополнительные калиевые каналы открываются в ответ на приток ионов кальция во время потенциала действия. Внутриклеточная концентрация ионов калия временно необычайно низка, что делает мембранное напряжение V m еще ближе к равновесному напряжению калия E K . Мембранный потенциал становится ниже мембранного потенциала покоя. Следовательно, возникает недостаточная или гиперполяризация , называемая постгиперполяризацией , которая сохраняется до тех пор, пока проницаемость мембраны для калия не вернется к своему обычному значению, восстанавливая мембранный потенциал до состояния покоя. [48] [46]
Рефрактерный период
[ редактировать ]За каждым потенциалом действия следует период рефрактерности , который можно разделить на период абсолютной рефрактерности , в течение которого невозможно вызвать другой потенциал действия, и затем период относительной рефрактерности , в течение которого требуется более сильный, чем обычно, стимул. [43] [44] [45] Эти два рефрактерных периода обусловлены изменениями состояния молекул натриевых и калиевых каналов. При закрытии после потенциала действия натриевые каналы переходят в «инактивированное» состояние , в котором их нельзя заставить открыться независимо от мембранного потенциала — это приводит к периоду абсолютной рефрактерности. Даже после того, как достаточное количество натриевых каналов вернулось в состояние покоя, часто случается, что часть калиевых каналов остается открытой, что затрудняет деполяризацию мембранного потенциала и тем самым приводит к возникновению относительного рефрактерного периода. Поскольку плотность и подтипы калиевых каналов могут сильно различаться у разных типов нейронов, продолжительность относительного рефрактерного периода сильно варьирует. [ нужна ссылка ]
Абсолютный рефрактерный период в значительной степени ответственен за однонаправленное распространение потенциалов действия по аксонам. [49] В любой момент участок аксона позади активно активирующей части является рефрактерным, но участок перед ним, не активированный в последнее время, способен стимулироваться деполяризацией от потенциала действия.
Распространение
[ редактировать ]Потенциал действия, генерируемый на аксонном холмике, распространяется волнообразно вдоль аксона. [50] Токи, текущие внутрь в определенной точке аксона во время потенциала действия, распространяются вдоль аксона и деполяризуют соседние участки его мембраны. Если эта деполяризация достаточно сильна, она вызывает аналогичный потенциал действия на соседних участках мембраны. Этот основной механизм был продемонстрирован Аланом Ллойдом Ходжкиным в 1937 году. После раздавливания или охлаждения нервных сегментов и, таким образом, блокировки потенциалов действия, он показал, что потенциал действия, приходящий на одну сторону блока, может спровоцировать другой потенциал действия на другой, при условии, что заблокированный сегмент был достаточно коротким. [к]
Как только на участке мембраны возник потенциал действия, этому участку мембраны требуется время для восстановления, прежде чем он сможет снова сработать. На молекулярном уровне этот период абсолютной рефрактерности соответствует времени, необходимому для восстановления активируемых напряжением натриевых каналов после инактивации, т. е. для возвращения в закрытое состояние. [44] В нейронах существует множество типов калиевых каналов, активируемых напряжением. Некоторые из них инактивируются быстро (токи типа А), другие инактивируются медленно или не инактивируются вообще; эта изменчивость гарантирует, что всегда будет доступный источник тока для реполяризации, даже если некоторые из калиевых каналов инактивируются из-за предшествующей деполяризации. С другой стороны, все нейрональные натриевые каналы, активируемые напряжением, инактивируются в течение нескольких миллисекунд во время сильной деполяризации, что делает последующую деполяризацию невозможной до тех пор, пока значительная часть натриевых каналов не вернется в свое закрытое состояние. Хоть это и ограничивает частоту стрельбы, [51] период абсолютной рефрактерности гарантирует, что потенциал действия движется только в одном направлении вдоль аксона. [49] Токи, поступающие благодаря потенциалу действия, распространяются в обоих направлениях вдоль аксона. [52] Однако только неактивированная часть аксона может ответить потенциалом действия; часть, которая только что сработала, не реагирует до тех пор, пока потенциал действия не выйдет за пределы допустимого диапазона и не сможет рестимулировать эту часть. При обычной ортодромной проводимости потенциал действия распространяется от аксонного бугорка к синаптическим выступам (концам аксона); распространение в противоположном направлении, известное как антидромная проводимость , встречается очень редко. [53] Однако если лабораторный аксон стимулировать в его середине, обе половины аксона будут «свежими», т. е. необожженными; тогда будут генерироваться два потенциала действия: один движется к аксонному бугорку, а другой - к синаптическим выступам.
Миелин и сальтаторная проводимость
[ редактировать ]
Чтобы обеспечить быструю и эффективную передачу электрических сигналов в нервной системе, аксоны определенных нейронов покрыты миелиновыми оболочками. Миелин представляет собой многослойную мембрану, которая окружает аксон сегментами, разделенными промежутками, известными как перехваты Ранвье . Его продуцируют специализированные клетки: шванновские клетки исключительно периферической нервной системы и олигодендроциты исключительно центральной нервной системы . Миелиновая оболочка уменьшает емкость мембраны и увеличивает сопротивление мембраны в межузловых интервалах, тем самым обеспечивая быстрое скачкообразное движение потенциалов действия от узла к узлу. [л] [м] [н] Миелинизация встречается главным образом у позвоночных , но аналогичная система обнаружена у некоторых беспозвоночных, например у некоторых видов креветок . [the] Не все нейроны у позвоночных миелинизированы; например, аксоны нейронов, составляющих автономную нервную систему, как правило, не миелинизированы.
Миелин предотвращает попадание ионов в аксон или выход из него вдоль миелинизированных сегментов. Как правило, миелинизация увеличивает скорость проведения потенциалов действия и делает их более энергоэффективными. Независимо от того, скачет ли он или нет, средняя скорость проведения потенциала действия колеблется от 1 метра в секунду (м/с) до более 100 м/с и, как правило, увеличивается с увеличением диаметра аксона. [п]
Потенциалы действия не могут распространяться через мембрану в миелинизированных сегментах аксона. Однако ток переносится цитоплазмой, которой достаточно для деполяризации первого или второго последующего узла Ранвье . Вместо этого ионный ток от потенциала действия в одном узле Ранвье провоцирует другой потенциал действия в следующем узле; это кажущееся «перескакивание» потенциала действия от узла к узлу известно как скачкообразная проводимость . Хотя механизм скачкообразной проводимости был предложен в 1925 году Ральфом Лилли, [д] Первые экспериментальные доказательства скачкообразной проводимости были получены Ичиджи Тасаки. [р] и Тайджи Такеучи [с] [54] и от Эндрю Хаксли и Роберта Стэмпфли. [т] Напротив, в немиелинизированных аксонах потенциал действия провоцирует другой потенциал действия в непосредственно прилегающей мембране и непрерывно движется вниз по аксону, как волна.

Миелин имеет два важных преимущества: высокая скорость проводимости и энергоэффективность. Для аксонов, диаметр которых превышает минимальный (примерно 1 микрометр ), миелинизация увеличивает скорость проведения потенциала действия, обычно в десять раз. [v] И наоборот, при заданной скорости проводимости миелиновые волокна меньше, чем их немиелинизированные аналоги. Например, потенциалы действия движутся примерно с одинаковой скоростью (25 м/с) в миелинизированном аксоне лягушки и немиелинизированном аксоне гигантского кальмара , но аксон лягушки имеет примерно в 30 раз меньший диаметр и в 1000 раз меньшую площадь поперечного сечения. . Кроме того, поскольку ионные токи ограничиваются узлами Ранвье, гораздо меньше ионов «просачиваются» через мембрану, экономя метаболическую энергию. Такая экономия является существенным селективным преимуществом , поскольку нервная система человека использует примерно 20% метаболической энергии организма. [v]
Длина миелинизированных сегментов аксонов важна для успеха скачкообразной проводимости. Они должны быть как можно более длинными, чтобы максимизировать скорость проведения, но не настолько длинными, чтобы поступающий сигнал был слишком слабым, чтобы спровоцировать потенциал действия в следующем узле Ранвье. В природе миелиновые сегменты обычно достаточно длинные, чтобы пассивно распространяемый сигнал мог пройти как минимум два узла, сохраняя при этом достаточную амплитуду для запуска потенциала действия во втором или третьем узле. Таким образом, коэффициент безопасности скачкообразной проводимости высок, что позволяет в случае травмы осуществлять передачу в обход узлов. Однако потенциалы действия могут преждевременно прекращаться в определенных местах, где коэффициент безопасности низкий, даже в безмиелиновых нейронах; распространенным примером является точка ветвления аксона, где он делится на два аксона. [56]
Некоторые заболевания разрушают миелин и нарушают сальтаторную проводимость, снижая скорость проведения потенциалов действия. [В] Наиболее известным из них является рассеянный склероз , при котором разрушение миелина нарушает координацию движений. [57]
Теория кабеля
[ редактировать ]
Поток токов внутри аксона можно количественно описать с помощью кабельной теории. [58] и ее разработки, такие как компартментальная модель. [59] Теория кабеля была разработана в 1855 году лордом Кельвином для моделирования трансатлантического телеграфного кабеля. [х] и было показано, что оно имеет отношение к нейронам Ходжкиным и Раштоном в 1946 году. [и] В простой теории кабеля нейрон рассматривается как электрически пассивный, идеально цилиндрический передающий кабель, который можно описать уравнением в частных производных. [58]

где V ( x , t ) — напряжение на мембране в момент времени t и в позиции x по длине нейрона, и где λ и τ — характерные масштабы длины и времени, в которых эти напряжения затухают в ответ на стимул. . Ссылаясь на принципиальную схему справа, эти масштабы можно определить по сопротивлениям и емкостям на единицу длины. [60]


Эти шкалы времени и длины можно использовать для понимания зависимости скорости проводимости от диаметра нейрона в безмиелиновых волокнах. Например, временной масштаб τ увеличивается как с увеличением сопротивления мембраны r m, так и с увеличением емкости c m . По мере увеличения емкости необходимо передать больше заряда для создания заданного трансмембранного напряжения (по уравнению Q = CV ); по мере увеличения сопротивления в единицу времени передается меньше заряда, что замедляет уравновешивание. Аналогичным образом, если внутреннее сопротивление на единицу длины r i меньше в одном аксоне, чем в другом (например, из-за того, что радиус первого больше), длина пространственного распада λ становится больше и скорость проводимости потенциала действия должно увеличиться. Если трансмембранное сопротивление r m увеличивается, это снижает средний ток «утечки» через мембрану, что также приводит к λ увеличению , увеличивая скорость проводимости.
Прекращение действия
[ редактировать ]Химические синапсы
[ редактировать ]В общем, потенциалы действия, достигающие синаптических бугорков, вызывают нейромедиатора в синаптическую щель. высвобождение [С] Нейротрансмиттеры — это небольшие молекулы, которые могут открывать ионные каналы в постсинаптической клетке; большинство аксонов имеют один и тот же нейромедиатор на всех своих концах. Прибытие потенциала действия открывает потенциал-чувствительные кальциевые каналы в пресинаптической мембране; приток кальция заставляет пузырьки, наполненные нейротрансмиттером, мигрировать на поверхность клетки и высвобождать свое содержимое в синаптическую щель . [аа] Этот сложный процесс ингибируется нейротоксинами тетаноспазмином и ботулотоксином , которые ответственны за столбняк и ботулизм соответственно. [аб]

Электрические синапсы
[ редактировать ]Некоторые синапсы обходятся без «посредника» нейромедиатора и соединяют пресинаптические и постсинаптические клетки вместе. [и] Когда потенциал действия достигает такого синапса, ионные токи, текущие в пресинаптическую клетку, могут преодолеть барьер двух клеточных мембран и войти в постсинаптическую клетку через поры, известные как коннексоны . [объявление] Таким образом, ионные токи пресинаптического потенциала действия могут непосредственно стимулировать постсинаптическую клетку. Электрические синапсы обеспечивают более быструю передачу, поскольку они не требуют медленной диффузии нейротрансмиттеров через синаптическую щель. Следовательно, электрические синапсы используются всякий раз, когда решающее значение имеют быстрая реакция и координация времени, например, в бегства , сетчатке позвоночных рефлексах и сердце .
Нервно-мышечные соединения
[ редактировать ]Особым случаем химического синапса является мышечный переход , в котором аксон мотонейрона нервно - заканчивается на мышечном волокне . [но] В таких случаях высвобождаемым нейромедиатором является ацетилхолин , который связывается с рецептором ацетилхолина, интегральным мембранным белком в мембране ( сарколемме ) мышечного волокна. [из] Однако ацетилхолин не остается связанным; скорее, он диссоциирует и гидролизуется ферментом ацетилхолинэстеразой , расположенным в синапсе. Этот фермент быстро снижает раздражение мышц, что позволяет деликатно регулировать степень и время мышечного сокращения. Некоторые яды инактивируют ацетилхолинэстеразу, чтобы предотвратить этот контроль, например нервно-паралитические вещества зарин и табун . [в] и инсектициды диазинон и малатион . [ах]
Другие типы клеток
[ редактировать ]Сердечные потенциалы действия
[ редактировать ]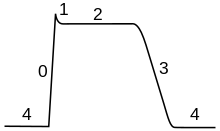
Сердечный потенциал действия отличается от потенциала действия нейронов наличием расширенного плато, при котором мембрана удерживается под высоким напряжением в течение нескольких сотен миллисекунд, прежде чем она, как обычно, реполяризуется током калия. [есть] Это плато возникает из-за более медленного открытия кальциевых каналов и удержания мембранного напряжения вблизи равновесного потенциала даже после инактивации натриевых каналов.
Потенциал действия сердца играет важную роль в координации сокращений сердца. [есть] Сердечные клетки синоатриального узла обеспечивают кардиостимуляторный потенциал , который синхронизирует работу сердца. Потенциалы действия этих клеток распространяются через атриовентрикулярный узел (АВ-узел), который обычно является единственным путем проводимости между предсердиями и желудочками . Потенциалы действия от АВ-узла проходят через пучок Гиса и оттуда к волокнам Пуркинье . [примечание 2] И наоборот, аномалии сердечного потенциала действия — будь то из-за врожденной мутации или травмы — могут привести к патологиям человека, особенно аритмиям . [есть] Некоторые антиаритмические препараты действуют на сердечный потенциал действия, например хинидин , лидокаин , бета-блокаторы и верапамил . [также]
Мышечные потенциалы действия
[ редактировать ]Потенциал действия в нормальной клетке скелетных мышц аналогичен потенциалу действия в нейронах. [61] Потенциалы действия возникают в результате деполяризации клеточной мембраны ( сарколеммы ), которая открывает чувствительные к напряжению натриевые каналы; они инактивируются, и мембрана реполяризуется за счет направленного наружу тока ионов калия. Потенциал покоя перед потенциалом действия обычно составляет -90 мВ, что несколько более отрицательно, чем у типичных нейронов. Потенциал действия мышцы длится примерно 2–4 мс, абсолютный рефрактерный период — примерно 1–3 мс, скорость проводимости по мышце — примерно 5 м/с. Потенциал действия высвобождает ионы кальция , которые высвобождают тропомиозин и позволяют мышцам сокращаться. Мышечные потенциалы действия провоцируются появлением пресинаптического потенциала действия нейронов в нервно-мышечном соединении , которое является общей мишенью для нейротоксинов . [в]
Потенциалы действия растений
[ редактировать ]растений и грибов Клетки [и] также являются электрически возбудимыми. Принципиальное отличие от потенциалов действия животных состоит в том, что деполяризация в растительных клетках осуществляется не за счет поглощения положительных ионов натрия, а за счет высвобождения отрицательных ионов хлора . [ал] [являюсь] [ан] В 1906 году Дж. К. Бозе опубликовал первые измерения потенциалов действия у растений, которые ранее были открыты Бердоном-Сандерсоном и Дарвином. [62] Увеличение цитоплазматических ионов кальция может быть причиной выброса анионов в клетку. Это делает кальций предшественником движения ионов, такого как приток отрицательных ионов хлорида и отток положительных ионов калия, как это наблюдается в листьях ячменя. [63]
Первоначальный приток ионов кальция также вызывает небольшую клеточную деполяризацию, вызывая открытие потенциалзависимых ионных каналов и позволяя ионам хлорида распространяться полной деполяризации.
Некоторые растения (например, Dionaea muscipula ) используют натриевые каналы для управления движениями растений и «подсчета» событий стимуляции, чтобы определить, достигнут ли порог движения. Dionaea muscipula , также известная как венерина мухоловка, встречается в субтропических водно-болотных угодьях Северной и Южной Каролины. [64] Когда почва бедна питательными веществами, мухоловка питается насекомыми и животными. [65] Несмотря на исследования этого растения, отсутствует понимание молекулярной основы венериных мухоловок и хищных растений в целом. [66]
Тем не менее, было проведено множество исследований потенциалов действия и того, как они влияют на движение и часовой механизм венерианской мухоловки. Начнем с того, что мембранный потенциал покоя венерианской мухоловки (-120 мВ) ниже, чем у клеток животных (обычно от -90 мВ до -40 мВ). [66] [67] Более низкий потенциал покоя облегчает активацию потенциала действия. Таким образом, когда насекомое попадает в ловушку растения, оно активирует волосоподобный механорецептор. [66] Затем этот рецептор активирует потенциал действия, который длится около 1,5 мс. [68] Это вызывает увеличение поступления положительных ионов кальция в клетку, слегка деполяризуя ее. Однако мухоловка не закрывается после одного срабатывания. Вместо этого требуется активация двух или более волос. [65] [66] Если срабатывает только один волос, активация игнорируется как ложное срабатывание. Далее второй волос должен быть активирован в течение определенного интервала времени (0,75–40 с), чтобы он зарегистрировался при первой активации. [66] Таким образом, начинается накопление кальция, который затем медленно падает после первого триггера. Когда в течение заданного интервала времени активируется второй потенциал действия, он достигает кальциевого порога и деполяризует клетку, закрывая ловушку на жертве за долю секунды. [66]
Вместе с последующим высвобождением положительных ионов калия потенциал действия у растений включает осмотическую потерю соли (KCl). В то время как потенциал действия животных осмотически нейтрален, поскольку равные количества входящего натрия и выходящего калия осмотически компенсируют друг друга. Взаимодействие электрических и осмотических связей в растительных клетках. [к] по-видимому, возникло в результате осмотической функции электрической возбудимости у общих одноклеточных предков растений и животных в условиях изменяющейся солености. Кроме того, нынешняя функция быстрой передачи сигналов рассматривается как новое достижение клеток многоклеточных животных в более стабильной осмотической среде. [69] Вероятно, известная сигнальная функция потенциалов действия у некоторых сосудистых растений (например, Mimosa pudica ) возникла независимо от таковой в возбудимых клетках многоклеточных животных.
В отличие от фазы подъема и пика, фаза спада и постгиперполяризация, по-видимому, зависят в первую очередь от катионов, не являющихся кальцием. Чтобы инициировать реполяризацию, клетке требуется перемещение калия из клетки посредством пассивного транспорта на мембране. В этом отличие от нейронов, поскольку движение калия не доминирует над снижением мембранного потенциала. Для полной реполяризации растительной клетке требуется энергия в виде АТФ, которая помогает высвободить водород из клетки, используя транспортер, называемый протонной АТФазой . [70] [66]
Таксономическое распространение и эволюционные преимущества
[ редактировать ]Потенциалы действия обнаруживаются во всех многоклеточных организмах , включая растения , беспозвоночных, таких как насекомые , и позвоночных, таких как рептилии и млекопитающие . [ап] Губки, по-видимому, являются основным типом многоклеточных эукариот , который не передает потенциалы действия, хотя некоторые исследования предполагают, что эти организмы также обладают формой электрической передачи сигналов. [ак] Потенциал покоя, а также размер и продолжительность потенциала действия не сильно изменились в ходе эволюции, хотя скорость проводимости резко меняется в зависимости от диаметра аксона и миелинизации.
| Животное | Тип ячейки | Потенциал покоя (мВ) | Увеличение АД (мВ) | Длительность AP (мс) | Скорость проводимости (м/с) |
|---|---|---|---|---|---|
| Кальмар ( Лолиго ) | Гигантский аксон | −60 | 120 | 0.75 | 35 |
| Дождевой червь ( Lumbricus ) | Среднее гигантское волокно | −70 | 100 | 1.0 | 30 |
| Таракан ( Перипланета ) | Гигантское волокно | −70 | 80–104 | 0.4 | 10 |
| Лягушка ( Рана ) | Аксон седалищного нерва | от −60 до −80 | 110–130 | 1.0 | 7–30 |
| Cat (FelisКот | Спинномозговой мотонейрон | от −55 до −80 | 80–110 | 1–1.5 | 30–120 |
Учитывая его сохранение на протяжении всей эволюции, потенциал действия, по-видимому, дает эволюционные преимущества. Одной из функций потенциалов действия является быстрая передача сигналов на большие расстояния внутри организма; скорость проводимости может превышать 110 м/с, что составляет одну треть скорости звука . Для сравнения: молекула гормона, переносимая в кровотоке, движется в крупных артериях со скоростью примерно 8 м/с. Частью этой функции является тесная координация механических процессов, таких как сокращение сердца. Вторая функция — это вычисления, связанные с ее генерацией. Будучи сигналом «все или ничего», который не затухает с расстоянием передачи, потенциал действия имеет те же преимущества, что и цифровая электроника . Интеграция различных дендритных сигналов на аксонном холмике и их пороговая обработка для формирования сложной последовательности потенциалов действия — это еще одна форма вычислений, которая использовалась биологически для формирования центральных генераторов паттернов и имитировалась в искусственных нейронных сетях .
Считается, что общий предок прокариот/эукариот, живший примерно четыре миллиарда лет назад, имел потенциалзависимые каналы. Эта функциональность, вероятно, в какой-то более поздний момент стала перекрестной для обеспечения механизма связи. Даже современные одноклеточные бактерии могут использовать потенциалы действия для связи с другими бактериями в той же биопленке . [72]
Экспериментальные методы
[ редактировать ]
Изучение потенциалов действия потребовало разработки новых экспериментальных методов. Первоначальная работа, до 1955 года, была выполнена в основном Аланом Ллойдом Ходжкиным и Эндрю Филдингом Хаксли , которые вместе с Джоном Кэрью Экклсом были удостоены Нобелевской премии по физиологии и медицине 1963 года за вклад в описание ионной основы нервов. проводимость. Он сосредоточился на трех целях: изолировать сигналы от отдельных нейронов или аксонов, разработать быструю и чувствительную электронику и уменьшить электроды настолько, чтобы можно было записать напряжение внутри одной клетки.
Первая проблема была решена путем изучения гигантских аксонов, обнаруженных в нейронах кальмара ( Loligo forbesii и Doryteuthis pealeii , в то время классифицировавшихся как Loligo pealeii ). [с] Эти аксоны настолько велики в диаметре (примерно 1 мм, что в 100 раз больше, чем у типичного нейрона), что их можно увидеть невооруженным глазом, что позволяет легко их извлекать и манипулировать ими. [я] [как] Однако они не являются репрезентативными для всех возбудимых клеток, и было изучено множество других систем с потенциалом действия.
Вторая проблема была решена с помощью решающей разработки фиксатора напряжения . [в] что позволило экспериментаторам изучать ионные токи, лежащие в основе потенциала действия, изолированно, и устранило ключевой источник электронного шума — ток I C , связанный с емкостью C мембраны. [74] Поскольку ток равен C, умноженному на скорость изменения трансмембранного напряжения V m , решением было разработать схему, которая поддерживала бы V m фиксированным (нулевая скорость изменения) независимо от токов, протекающих через мембрану. Таким образом, ток, необходимый для поддержания фиксированного значения V m, является прямым отражением тока, протекающего через мембрану. Другие достижения в области электроники включали использование клеток Фарадея и электроники с высоким входным сопротивлением , так что само измерение не влияло на измеряемое напряжение. [75]
Третья проблема — получение электродов, достаточно маленьких для регистрации напряжения внутри одного аксона, не нарушая его, — была решена в 1949 году с изобретением стеклянного электрода для микропипеток. [В] который был быстро принят другими исследователями. [из] [оу] Усовершенствования этого метода позволяют производить наконечники электродов толщиной до 100 Å (10 нм ), что также обеспечивает высокий входной импеданс. [76] Потенциалы действия также можно регистрировать с помощью небольших металлических электродов, расположенных рядом с нейроном, с помощью нейрочипов, содержащих EOSFET , или оптически с помощью красителей, чувствительных к Ca. 2+ или к напряжению. [топор]

В то время как стеклянные электроды-микропипетки измеряют сумму токов, проходящих через множество ионных каналов, изучение электрических свойств одного ионного канала стало возможным в 1970-х годах с разработкой патч-зажима Эрвином Неером и Бертом Сакманном . За это открытие они были удостоены Нобелевской премии по физиологии и медицине в 1991 году. [нижнегреческий 3] Зажим патчей подтвердил, что ионные каналы имеют дискретные состояния проводимости, такие как открытое, закрытое и инактивированное.
В последние годы были разработаны технологии оптической визуализации для измерения потенциалов действия либо посредством одновременной многозонной записи, либо с ультрапространственным разрешением. С помощью чувствительных к напряжению красителей оптически зарегистрировали потенциалы действия на крошечном участке мембраны кардиомиоцитов . [является]
Нейротоксины
[ редактировать ]
Некоторые нейротоксины , как природные, так и синтетические, блокируют потенциал действия. Тетродотоксин из рыбы-фугу и сакситоксин из Gonyaulax ( род динофлагеллят, ответственный за « красные приливы ») блокируют потенциалы действия, ингибируя потенциал-чувствительный натриевый канал; [the] Точно так же дендротоксин черной змеи мамбы ингибирует чувствительный к напряжению калиевый канал. Такие ингибиторы ионных каналов служат важной исследовательской цели, позволяя ученым «отключать» определенные каналы по своему желанию, изолируя таким образом вклад других каналов; они также могут быть полезны при очистке ионных каналов с помощью аффинной хроматографии или при определении их концентрации. Однако такие ингибиторы также создают эффективные нейротоксины и рассматривались для использования в качестве химического оружия . Нейротоксины, направленные на ионные каналы насекомых, оказались эффективными инсектицидами ; Одним из примеров является синтетический перметрин , который продлевает активацию натриевых каналов, участвующих в потенциалах действия. Ионные каналы насекомых настолько отличаются от своих аналогов у человека, что у человека возникает мало побочных эффектов.
История
[ редактировать ]
Роль электричества в нервной системе животных впервые наблюдал у рассеченных лягушек Луиджи Гальвани , изучавший его с 1791 по 1797 год. [нет] Результаты Гальвани вдохновили Алессандро Вольту на разработку Вольтова батареи — самой ранней из известных электрических батарей , — с помощью которой он изучал электричество животных (таких как электрические угри ) и физиологические реакции на приложенное постоянного тока напряжение . [бб]
В 19 веке ученые изучали распространение электрических сигналов в целых нервах (т. е. пучках нейронов ) и продемонстрировали, что нервная ткань состоит из клеток , а не из взаимосвязанной сети трубок ( сеточки ). [77] Карло Маттеуччи продолжил исследования Гальвани и продемонстрировал, что поврежденные нервы и мышцы лягушек могут производить постоянный ток . Работа Маттеуччи вдохновила немецкого физиолога Эмиля дю Буа-Реймона , который в 1843 году обнаружил, что стимуляция этих мышечных и нервных препаратов приводит к заметному уменьшению их токов покоя, что сделало его первым исследователем, определившим электрическую природу потенциала действия. [78] Скорость проводимости потенциалов действия была затем измерена в 1850 году другом Дюбуа-Реймона Германом фон Гельмгольцем . [79] После этого прогресс в электрофизиологии застопорился из-за ограничений химической теории и экспериментальной практики. Чтобы установить, что нервная ткань состоит из отдельных клеток, испанский врач Сантьяго Рамон-и-Кахаль и его ученики использовали краску, разработанную Камилло Гольджи, чтобы выявить бесчисленные формы нейронов, которые они кропотливо воспроизвели. За свои открытия Гольджи и Рамон-и-Кахаль были удостоены Нобелевской премии по физиологии 1906 года . [нижнегреческий 4] Их работа разрешила давний спор в нейроанатомии XIX века; Сам Гольджи выступал за сетевую модель нервной системы.

В XX веке произошел значительный прорыв в электрофизиологии. В 1902 и 1912 годах Юлиус Бернштейн выдвинул гипотезу о том, что потенциал действия возникает в результате изменения проницаемости аксональной мембраны для ионов. [до н. э.] [80] Гипотеза Бернштейна была подтверждена Кеном Коулом и Говардом Кертисом, которые показали, что проводимость мембраны увеличивается во время потенциала действия. [бд] В 1907 году Луи Лапик предположил, что потенциал действия генерируется при пересечении порога. [быть] то, что позже будет показано как продукт динамических систем ионной проводимости. В 1949 году Алан Ходжкин и Бернард Кац уточнили гипотезу Бернштейна, посчитав, что аксональная мембрана может иметь разную проницаемость для разных ионов; в частности, они продемонстрировали решающую роль натриевой проницаемости для потенциала действия. [бф] Они впервые зарегистрировали электрические изменения в мембране нейронов, которые опосредуют потенциал действия. [нижнегреческий 5] Кульминацией этого направления исследований стали пять статей 1952 года Ходжкина, Каца и Эндрю Хаксли , в которых они применили технику фиксации напряжения для определения зависимости проницаемости аксональной мембраны для ионов натрия и калия от напряжения и времени, на основании чего они смогли количественно реконструировать потенциал действия. [я] Ходжкин и Хаксли соотнесли свойства своей математической модели с дискретными ионными каналами , которые могли существовать в нескольких различных состояниях, включая «открытое», «закрытое» и «инактивированное». Их гипотезы были подтверждены в середине 1970-х и 1980-х годах Эрвином Неером и Бертом Сакманном , которые разработали технику фиксации патчей для изучения состояний проводимости отдельных ионных каналов. [бг] В 21 веке исследователи начинают понимать структурную основу этих состояний проводимости и селективности каналов для своих видов ионов. [бх] атомного разрешения через кристаллические структуры , [с] измерение расстояния флуоресценции [bj] и исследования криоэлектронной микроскопии . [бк]
Юлиус Бернштейн был также первым, кто ввел уравнение Нернста для потенциала покоя на мембране; это было обобщено Дэвидом Э. Голдманом на одноименное уравнение Гольдмана в 1943 году. [час] Натриево -калиевый насос был открыт в 1957 году. [бл] [нижнегреческий 6] и его свойства постепенно выяснены, [бм] [бн] [быть] кульминацией которого стало определение его структуры с атомным разрешением с помощью рентгеновской кристаллографии . [бп] Кристаллические структуры родственных ионных насосов также были решены, что дает более широкое представление о том, как работают эти молекулярные машины . [бк]
Количественные модели
[ редактировать ]
Математические и вычислительные модели необходимы для понимания потенциала действия и предлагают прогнозы, которые можно проверить на экспериментальных данных, обеспечивая строгую проверку теории. Наиболее важной и точной из ранних нейронных моделей является модель Ходжкина-Хаксли , которая описывает потенциал действия с помощью связанного набора четырех обыкновенных дифференциальных уравнений (ОДУ). [я] Хотя модель Ходжкина-Хаксли может быть упрощением с некоторыми ограничениями. [81] по сравнению с реалистичной нервной мембраной, существующей в природе, ее сложность вдохновила на создание нескольких еще более упрощенных моделей. [82] [бр] например, модель Морриса – Лекара [бс] и модель ФитцХью – Нагумо , [бт] оба из которых имеют только два связанных ОДУ. Свойства моделей Ходжкина-Хаксли и ФитцХью-Нагумо и их родственников, таких как модель Бонхёффера-Ван дер Поля, [этот] были хорошо изучены в области математики, [83] [бв] вычисление [84] и электроника. [ш] Однако простые модели генераторного потенциала и потенциала действия не могут точно воспроизвести частоту и форму нервных спайков вблизи порога, особенно для механорецепторов , таких как тельца Пачини . [85] Более современные исследования сосредоточены на более крупных и интегрированных системах; объединяя модели потенциала действия с моделями других частей нервной системы (например, дендритов и синапсов), исследователи могут изучать нейронные вычисления. [86] и простые рефлексы , такие как рефлексы бегства и другие, контролируемые центральными генераторами паттернов . [87] [бх]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ В целом, хотя это простое описание инициации потенциала действия является точным, оно не объясняет такие явления, как блок возбуждения (способность препятствовать активации нейронами потенциалов действия путем стимуляции их большими шагами тока) и способность вызывать потенциалы действия путем кратковременно гиперполяризуя мембрану. Однако , анализируя динамику системы натриевых и калиевых каналов в мембранном пластыре с помощью компьютерных моделей , эти явления легко объяснить. [нижнегреческий 1]
- ^ Эти волокна Пуркинье являются мышечными волокнами и не связаны с клетками Пуркинье , которые представляют собой нейроны, обнаруженные в мозжечке .
Ссылки
[ редактировать ]Сноски
[ редактировать ]- ^ Ходжкин А.Л., Хаксли А.Ф. (август 1952 г.). «Количественное описание мембранного тока и его применение к проводимости и возбуждению нерва» . Журнал физиологии . 117 (4): 500–44. doi : 10.1113/jphysicalol.1952.sp004764 . ПМК 1392413 . ПМИД 12991237 .
- ^ Уильямс Дж. А. (февраль 1981 г.). «Электрические корреляты секреции в эндокринных и экзокринных клетках». Протокол ФРС . 40 (2): 128–34. ПМИД 6257554 .
- ^ «Сокращение сердечной мышцы» . Проверено 28 мая 2021 г.
- ^ Пикард Б. (июнь 1973 г.). «Потенциалы действия высших растений» (PDF) . Ботаническое обозрение . 39 (2): 188. Бибкод : 1973BotRv..39..172P . дои : 10.1007/BF02859299 . S2CID 5026557 .
- ^ Летерье С (февраль 2018 г.). «Начальный сегмент аксона: обновленная точка зрения» . Журнал неврологии . 38 (9): 2135–2145. doi : 10.1523/JNEUROSCI.1922-17.2018 . ПМК 6596274 . ПМИД 29378864 .
- ^ Первс Д., Августин Г.Дж., Фитцпатрик Д. и др., ред. (2001). «Ионные каналы, управляемые напряжением» . Нейронаука (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. Архивировано из оригинала 5 июня 2018 года . Проверено 29 августа 2017 г.
- ^ Перейти обратно: а б с д и ж г час Баллок, Орканд и Гриннелл, 1977 , стр. 150–151.
- ^ Перейти обратно: а б с д и Янг, 1981 , стр. 89–90.
- ^ Перейти обратно: а б Шмидт-Нильсен 1997 , стр. 484.
- ^ Перейти обратно: а б с Пурвес и др. 2008 , стр. 48–49; Баллок, Орканд и Гриннелл, 1977 , с. 141; Шмидт-Нильсен 1997 , с. 483; Юнге 1981 , с. 89.
- ^ Стивенс 1966 , с. 127.
- ^ Шмидт-Нильсен , стр. 484.
- ^ Тамагава Х., Фунатани М., Икеда К. (январь 2016 г.). «Теория адсорбции Линга как механизм генерации мембранного потенциала, наблюдаемый как в живых, так и в неживых системах» . Мембраны . 6 (1): 11. doi : 10.3390/membranes6010011 . ПМЦ 4812417 . ПМИД 26821050 .
- ^ Перейти обратно: а б с д и Санес Д.Х., Рех Т.А. (1 января 2012 г.). Развитие нервной системы (Третье изд.). Эльзевир Академик Пресс. стр. 211–214. ISBN 9780080923208 . OCLC 762720374 .
- ^ Куропатка Д. (1991). Кальциевые каналы: их свойства, функции, регуляция и клиническое значение . ЦРК Пресс. стр. 138–142. ISBN 9780849388071 .
- ^ Черный I (1984). Клеточная и молекулярная биология развития нейронов | Ира Блэк | Спрингер . Спрингер. п. 103. ИСБН 978-1-4613-2717-2 . Архивировано из оригинала 17 июля 2017 года.
- ^ Педерсен Р. (1998). Актуальные темы биологии развития, том 39 . Эльзевир Академик Пресс. ISBN 9780080584621 .
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 11.
- ^ Сильверторн 2010 , с. 253.
- ^ Перейти обратно: а б с Пурвес и др. 2008 , стр. 49–50; Баллок, Орканд и Гриннелл, 1977 , стр. 140–141; Шмидт-Нильсен 1997 , стр. 480–481.
- ^ Перейти обратно: а б с д Шмидт-Нильсен 1997 , стр. 483–484.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 177–240; Шмидт-Нильсен 1997 , стр. 490–499; Стивенс 1966 , с. 47–68.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 178–180; Шмидт-Нильсен 1997 , стр. 490–491.
- ^ Первес и др. 2001 .
- ^ Первес и др. 2008 , стр. 26–28.
- ^ «Миелинизация увеличивает пространственный масштаб аналоговой модуляции синаптической передачи: исследование моделирования» . Границы клеточной нейронауки .
- ^ Збили, М.; Дебанн, Д. (2019). «Прошлое и будущее аналого-цифровой модуляции синаптической передачи» . Границы клеточной нейронауки . 13 :160. дои : 10.3389/fncel.2019.00160 . ПМК 6492051 . ПМИД 31105529 .
- ^ Кларк, Беверли; Хойссер, Михаэль (8 августа 2006 г.). «Нейронное кодирование: аналоговая передача сигналов в аксонах» . Современная биология . 16 (15): 585–588 р. дои : 10.1016/j.cub.2006.07.007 . ПМИД 16890514 . S2CID 8295969 .
- ^ Лю, Венке; Лю, Цин; Крозье, Роберт А.; Дэвис, Робин Л. (2021). «Аналоговая передача тонкой структуры потенциала действия в аксонах спиральных ганглиев» . Журнал нейрофизиологии . 126 (3): 888–905. дои : 10.1152/jn.00237.2021 . ПМЦ 8461829 . ПМИД 34346782 .
- ^ Шмидт-Нильсен 1997 , стр. 535–580; Баллок, Орканд и Гриннелл, 1977 , стр. 49–56, 76–93, 247–255; Стивенс 1966 , стр. 69–79.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 53; Баллок, Орканд и Гриннелл, 1977 , стр. 122–124.
- ^ Янг 1981 , стр. 115–132.
- ^ Перейти обратно: а б Баллок, Орканд и Гриннелл, 1977 , стр. 152–153.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 444–445.
- ^ Первес и др. 2008 , с. 38.
- ^ Стивенс 1966 , стр. 127–128.
- ^ Первес и др. 2008 , стр. 61–65.
- ^ Первес и др. 2008 , стр. 64–74; Баллок, Орканд и Гриннелл, 1977 , стр. 149–150; Юнге 1981 , стр. 84–85; Стивенс 1966 , стр. 152–158.
- ^ Перейти обратно: а б с Пурвес и др. 2008 , с. 47; Пурвес и др. 2008 , с. 65; Баллок, Орканд и Гриннелл, 1977 , стр. 147–148; Стивенс 1966 , с. 128.
- ^ Голдин, А.Л. в Waxman 2007 , Нейрональные каналы и рецепторы , стр. 43–58.
- ^ Стивенс 1966 , с. 49.
- ^ Первес и др. 2008 , с. 34; Баллок, Орканд и Гриннелл, 1977 , с. 134; Шмидт-Нильсен 1997 , стр. 478–480.
- ^ Перейти обратно: а б с д Пурвес и др. 2008 , с. 49.
- ^ Перейти обратно: а б с д Стивенс 1966 , стр. 19–20.
- ^ Перейти обратно: а б с Баллок, Орканд и Гриннелл, 1977 , с. 151; Юнге 1981 , стр. 4–5.
- ^ Перейти обратно: а б Баллок, Орканд и Гриннелл, 1977 , с. 152.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 147–149; Стивенс 1966 , стр. 126–127.
- ^ Первес и др. 2008 , с. 37.
- ^ Перейти обратно: а б Пурвес и др. 2008 , с. 56.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 160–164.
- ^ Стивенс 1966 , стр. 21–23.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 161–164.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 509.
- ^ Тасаки, Я в поле 1959 , стр. 75–121.
- ^ Шмидт-Нильсен 1997 , рисунок 12.13.
- ^ Баллок, Орканд и Гриннелл 1977 , стр. 163.
- ^ Ваксман, С.Г. в Waxman 2007 , Рассеянный склероз как нейродегенеративное заболевание , стр. 333–346.
- ^ Перейти обратно: а б Ралл, В. Кох и Сегев, 1989 , Теория кабеля для дендритных нейронов , стр. 9–62.
- ^ Сегев I, Флешман Дж.В., Берк Р.Э. (1989). «Компартментные модели сложных нейронов». В Кох С. , Сегев I (ред.). Методы нейронного моделирования: от синапсов к сетям . Кембридж, Массачусетс: MIT Press. стр. 63–96. ISBN 978-0-262-11133-1 . LCCN 88008279 . OCLC 18384545 .
- ^ Первес и др. 2008 , стр. 52–53.
- ^ Ганонг 1991 , стр. 59–60.
- ^ Тандон, Пракаш Н. (1 июля 2019 г.). «Джагдиш Чандра Бос и нейробиология растений: Часть I» (PDF) . Индийский журнал истории науки . 54 (2). дои : 10.16943/ijhs/2019/v54i2/49660 . ISSN 0019-5235 .
- ^ Фелле Х.Х., Циммерманн М.Р. (июнь 2007 г.). «Системная передача сигналов в ячмене через потенциалы действия». Планта . 226 (1): 203–14. Бибкод : 2007Plant.226..203F . дои : 10.1007/s00425-006-0458-y . ПМИД 17226028 . S2CID 5059716 .
- ^ Люкен Д.О. (декабрь 2005 г.). «Места обитания Dionaea muscipula (Ловушка Венеры), Droseraceae, связанных с заливами Каролины». Юго-восточный натуралист . 4 (4): 573–584. doi : 10.1656/1528-7092(2005)004[0573:HODMVF]2.0.CO;2 . ISSN 1528-7092 . S2CID 9246114 .
- ^ Перейти обратно: а б Бём Дж., Шерцер С., Крол Э., Кройцер И., фон Мейер К., Лори С. и др. (февраль 2016 г.). «Венерина мухоловка Dionaea muscipula подсчитывает потенциал действия, вызванный добычей, для индуцирования поглощения натрия» . Современная биология . 26 (3): 286–95. Бибкод : 2016CBio...26..286B . дои : 10.1016/j.cub.2015.11.057 . ПМЦ 4751343 . ПМИД 26804557 .
- ^ Перейти обратно: а б с д и ж г Хедрих Р., Неер Э. (март 2018 г.). «Венерина мухоловка: как работает возбудимое хищное растение». Тенденции в науке о растениях . 23 (3): 220–234. Бибкод : 2018TPS....23..220H . doi : 10.1016/j.tplants.2017.12.004 . ПМИД 29336976 .
- ^ Первес Д., Августин Г.Дж., Фитцпатрик Д. и др., редакторы. Нейронаука. 2-е издание. Сандерленд (Массачусетс): Sinauer Associates; 2001. Электрические потенциалы мембран нервных клеток. Доступно: [1]
- ^ Волков А.Г., Адесина Т., Йованов Е. (май 2007 г.). «Закрытие венерины мухоловки путем электростимуляции двигательных клеток» . Сигнализация и поведение растений . 2 (3): 139–45. Бибкод : 2007PlSiB...2..139В . дои : 10.4161/psb.2.3.4217 . ПМК 2634039 . ПМИД 19516982 .
- ^ Градманн, Д; Маммерт, Х. в Спансвике, Лукасе и Дейнти, 1980 , Потенциалы действия растений , стр. 333–344.
- ^ Опритов В.А. и др. «Прямая связь генерации потенциала действия в клетках высшего растения (Cucurbita Pepo) с работой электрогенного насоса». Российский журнал физиологии растений . 49, нет. 1, 2002, стр. 142–147.
- ^ Баллок и Хорридж 1965 .
- ^ Кристан ВБ (октябрь 2016 г.). «Ранняя эволюция нейронов» . Современная биология . 26 (20): Р949–Р954. Бибкод : 2016CBio...26.R949K . дои : 10.1016/j.cub.2016.05.030 . ПМИД 27780067 .
- ^ Хеллиер Дж.Л. (2014). Мозг, нервная система и их болезни . АВС-Клио. п. 532. ИСБН 9781610693387 .
- ^ Янг 1981 , стр. 63–82.
- ^ Кеттенманн и Грантин 1992 .
- ^ Снелл, FM в Лавалле, Шанне и Эберте, 1969 , Некоторые электрические свойства микроэлектродов-пипеток с тонкими наконечниками .
- ^ Мангал 1961 ; МакГенри и Гаррисон, 1969 ; Уорден, Свази и Адельман, 1975 .
- ^ Финкельштейн Г.В. (2013). Эмиль дю Буа-Реймон: нейробиология, личность и общество в Германии девятнадцатого века . Кембридж, Массачусетс. ISBN 9781461950325 . OCLC 864592470 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Олеско, Кэтрин М. и Фредерик Л. Холмс. «Эксперимент, количественная оценка и открытия: ранние физиологические исследования Гельмгольца, 1843-50». В книге «Герман фон Гельмгольц и основы науки девятнадцатого века» под ред. Дэвид Кэхан, 50–108. Беркли; Лос-Анджелес; Лондон: Калифорнийский университет, 1994.
- ^ Бернштейн 1912 .
- ^ Баранаускас Г., Мартина М. (январь 2006 г.). «Токи натрия активируются без задержки типа Ходжкина и Хаксли в центральных нейронах млекопитающих» . Журнал неврологии . 26 (2): 671–84. doi : 10.1523/jneurosci.2283-05.2006 . ПМК 6674426 . ПМИД 16407565 .
- ^ Хоппенстедт 1986 .
- ^ Сато, С; Фукай, Х; Номура, Т; Дой, С. в Reeke et al. 2005 , Бифуркационный анализ уравнений Ходжкина-Хаксли , стр. 459–478.
* ФитцХью, Р. в Шванне, 1969 , Математические модели возбуждения и распространения в нерве , стр. 12–16.
* Гукенхаймер и Холмс, 1986 , стр. 12–16. - ^ Нельсон, Мэн; Ринзель, Дж., Bower & Beeman, 1995 , Модель Ходжкина-Хаксли , стр. 29–49.
* Ринцель, Дж. и Эрментраут, Великобритания; в Кох и Сегев, 1989 , Анализ нервной возбудимости и колебаний , стр. 135–169. - ^ Бисвас А., Маниваннан М., Сринивасан М.А. (2015). «Порог вибротактильной чувствительности: нелинейная стохастическая модель механотрансдукции тельца Пачини» . Транзакции IEEE на тактильных ощущениях . 8 (1): 102–13. дои : 10.1109/ТОХ.2014.2369422 . ПМИД 25398183 . S2CID 15326972 .
- ^ МакКаллок 1988 , стр. 19–39, 46–66, 72–141; Андерсон и Розенфельд 1988 , стр. 15–41.
- ^ Геттинг, П.А. в книге Кох и Сегев, 1989 , Реконструкция малых нейронных сетей , стр. 171–194.
Журнальные статьи
[ редактировать ]- ^ Макдональд П.Е., Рорсман П. (февраль 2006 г.). «Колебания, межклеточное взаимодействие и секреция инсулина в бета-клетках поджелудочной железы» . ПЛОС Биология . 4 (2): е49. дои : 10.1371/journal.pbio.0040049 . ПМЦ 1363709 . ПМИД 16464129 .

- ^ Перейти обратно: а б Барнетт М.В., Ларкман, премьер-министр (июнь 2007 г.). «Потенциал действия» . Практическая неврология . 7 (3): 192–7. ПМИД 17515599 . Архивировано из оригинала 8 июля 2011 года.
- ^ Голдинг Н.Л., Кэт В.Л., Спрастон Н. (декабрь 2001 г.). «Дихотомия обратного распространения потенциала действия в дендритах пирамидальных нейронов CA1». Журнал нейрофизиологии . 86 (6): 2998–3010. дои : 10.1152/jn.2001.86.6.2998 . ПМИД 11731556 . S2CID 2915815 .
- ^ Сасаки, Т., Мацуки, Н., Икегая, Ю. 2011 Модуляция потенциала действия во время аксональной проводимости Science 331 (6017), стр. 599–601
- ^ Аур Д., Коннолли С.И., Джог М.С. (ноябрь 2005 г.). «Вычисление направленности импульсов с помощью тетродов». Журнал методов нейробиологии . 149 (1): 57–63. doi : 10.1016/j.jneumeth.2005.05.006 . ПМИД 15978667 . S2CID 34131910 .
- ^ Аур Д., Джог, MS., 2010 Нейроэлектродинамика: понимание языка мозга, IOS Press, 2010. два : 10.3233/978-1-60750-473-3-i
- ^ Ноубл Д. (ноябрь 1960 г.). «Сердечная деятельность и потенциалы кардиостимулятора на основе уравнений Ходжкина-Хаксли». Природа . 188 (4749): 495–7. Бибкод : 1960Natur.188..495N . дои : 10.1038/188495b0 . ПМИД 13729365 . S2CID 4147174 .
- ^ Перейти обратно: а б Гольдман Д.Е. (сентябрь 1943 г.). «Потенциал, импеданс и выпрямление в мембранах» . Журнал общей физиологии . 27 (1): 37–60. дои : 10.1085/jgp.27.1.37 . ПМК 2142582 . ПМИД 19873371 .
- ^ Перейти обратно: а б с д и Ходжкин А.Л. , Хаксли А.Ф., Кац Б. (апрель 1952 г.). «Измерение вольт-амперных отношений в мембране гигантского аксона Лолиго» . Журнал физиологии . 116 (4): 424–48. дои : 10.1113/jphysicalol.1952.sp004716 . ПМЦ 1392219 . ПМИД 14946712 .
* Ходжкин А.Л. , Хаксли А.Ф. (апрель 1952 г.). «Токи, переносимые ионами натрия и калия через мембрану гигантского аксона Лолиго» . Журнал физиологии . 116 (4): 449–72. дои : 10.1113/jphysicalol.1952.sp004717 . ПМЦ 1392213 . ПМИД 14946713 .
* Ходжкин А.Л. , Хаксли А.Ф. (апрель 1952 г.). «Компоненты мембранной проводимости гигантского аксона Лолиго» . Журнал физиологии . 116 (4): 473–96. doi : 10.1113/jphysicalol.1952.sp004718 . ПМК 1392209 . ПМИД 14946714 .
* Ходжкин А.Л. , Хаксли А.Ф. (апрель 1952 г.). «Двойное влияние мембранного потенциала на натриевую проводимость в гигантском аксоне Лолиго» . Журнал физиологии . 116 (4): 497–506. дои : 10.1113/jphysicalol.1952.sp004719 . ПМЦ 1392212 . ПМИД 14946715 .
* Ходжкин А.Л. , Хаксли А.Ф. (август 1952 г.). «Количественное описание мембранного тока и его применение к проводимости и возбуждению нерва» . Журнал физиологии . 117 (4): 500–44. doi : 10.1113/jphysicalol.1952.sp004764 . ПМК 1392413 . ПМИД 12991237 . - ^ Наундорф Б., Вольф Ф., Волгушев М. (апрель 2006 г.). «Уникальные особенности инициации потенциала действия в корковых нейронах» (PDF) . Природа . 440 (7087): 1060–3. Бибкод : 2006Natur.440.1060N . дои : 10.1038/nature04610 . ПМИД 16625198 . S2CID 1328840 . Архивировано из оригинала (PDF) 20 декабря 2018 года . Проверено 24 сентября 2019 г.
- ^ Ходжкин А.Л. (июль 1937 г.). «Доказательства передачи электрического тока в нервах: Часть I» . Журнал физиологии . 90 (2): 183–210. дои : 10.1113/jphysicalol.1937.sp003507 . ПМК 1395060 . ПМИД 16994885 .
* Ходжкин А.Л. (июль 1937 г.). «Доказательства передачи электрического тока в нервах: Часть II» . Журнал физиологии . 90 (2): 211–32. doi : 10.1113/jphysicalol.1937.sp003508 . ПМК 1395062 . ПМИД 16994886 . - ^ Зальц Б (2006). «Приобретение миелина: история успеха». Пуринергическая передача сигналов во взаимодействиях нейронов и глии . Симпозиумы Фонда Новартис. Том. 276. стр. 15–21, обсуждение 21–5, 54–7, 275–81. дои : 10.1002/9780470032244.ch3 . ISBN 978-0-470-03224-4 . ПМИД 16805421 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Поляк С., Пелеш Е (декабрь 2003 г.). «Локальная дифференциация миелинизированных аксонов в узлах Ранвье». Обзоры природы. Нейронаука . 4 (12): 968–80. дои : 10.1038/nrn1253 . ПМИД 14682359 . S2CID 14720760 .
- ^ Саймонс М., Троттер Дж. (октябрь 2007 г.). «Подведем итоги: клеточная биология миелинизации». Современное мнение в нейробиологии . 17 (5): 533–40. дои : 10.1016/j.conb.2007.08.003 . ПМИД 17923405 . S2CID 45470194 .
- ^ Сюй К., Теракава С. (август 1999 г.). «Фенестрационные узлы и широкое субмиелиновое пространство составляют основу необычайно быстрого проведения импульсов миелинизированными аксонами креветок» . Журнал экспериментальной биологии . 202 (Часть 15): 1979–89. дои : 10.1242/jeb.202.15.1979 . ПМИД 10395528 .
- ^ Перейти обратно: а б Хурш Дж. Б. (1939). «Скорость проводимости и диаметр нервных волокон». Американский журнал физиологии . 127 : 131–39. дои : 10.1152/ajplegacy.1939.127.1.131 .
- ^ Лилли Р.С. (март 1925 г.). «Факторы, влияющие на передачу и восстановление в модели пассивного железного нерва» . Журнал общей физиологии . 7 (4): 473–507. дои : 10.1085/jgp.7.4.473 . ПМК 2140733 . ПМИД 19872151 . См. также Кейнс и Эйдли, 1991 , с. 78
- ^ Тасаки I (1939). «Электросальтаторная передача нервного импульса и воздействие наркоза на нервные волокна». Являюсь. Дж. Физиол . 127 : 211–27. дои : 10.1152/ajplegacy.1939.127.2.211 .
- ^ Тасаки I, Такеучи Т (1941). «Ток действия, возникающий в узле Ранвье, и его значение для проведения возбуждения». Архив Пфлюгера по всей физиологии . 244 (6): 696–711. дои : 10.1007/BF01755414 . S2CID 8628858 .
* Тасаки I, Такеучи Т (1942). «Дальнейшие исследования тока действия медулированного нервного волокна и электросальтационной передачи нервного импульса». Архив Пфлюгера по всей физиологии . 245 (5): 764–82. дои : 10.1007/BF01755237 . S2CID 44315437 . - ^ Хаксли А.Ф. , Штемпфли Р. (май 1949 г.). «Доказательства скачкообразной проводимости в периферических миелинизированных нервных волокнах» . Журнал физиологии . 108 (3): 315–39. дои : 10.1113/jphysicalol.1949.sp004335 . ПМЦ 1392492 . ПМИД 16991863 .
* Хаксли А.Ф. , Стампфли Р. (февраль 1951 г.). «Прямое определение мембранного потенциала покоя и потенциала действия в одиночных миелинизированных нервных волокнах» . Журнал физиологии . 112 (3–4): 476–95. doi : 10.1113/jphysicalol.1951.sp004545 . ПМК 1393015 . ПМИД 14825228 . - ^ Раштон, Вашингтон (сентябрь 1951 г.). «Теория влияния размера волокон на мозговой нерв» . Журнал физиологии . 115 (1): 101–22. doi : 10.1113/jphysicalol.1951.sp004655 . ПМК 1392008 . ПМИД 14889433 .
- ^ Перейти обратно: а б Хартлайн Д.К., Колман Д.Р. (январь 2007 г.). «Быстрая проводимость и эволюция гигантских аксонов и миелинизированных волокон» . Современная биология . 17 (1): Р29-35. Бибкод : 2007CBio...17R..29H . дои : 10.1016/j.cub.2006.11.042 . ПМИД 17208176 . S2CID 10033356 .
- ^ Миллер Р.Х., Ми С. (ноябрь 2007 г.). «Рассекающая демиелинизация». Природная неврология . 10 (11): 1351–4. дои : 10.1038/nn1995 . ПМИД 17965654 . S2CID 12441377 .
- ^ Кельвин В.Т. (1855 г.). «К теории электрического телеграфа». Труды Королевского общества . 7 : 382–99. дои : 10.1098/rspl.1854.0093 . S2CID 178547827 .
- ^ Ходжкин А.Л. , Раштон В.А. (декабрь 1946 г.). «Электрические константы нервного волокна ракообразных» . Труды Королевского медицинского общества . 134 (873): 444–79. Бибкод : 1946РСПСБ.133..444Н . дои : 10.1098/rspb.1946.0024 . ПМИД 20281590 .
- ^ Зюудхоф ТЦ (2008). «Высвобождение нейротрансмиттера». Фармакология высвобождения нейромедиаторов . Справочник по экспериментальной фармакологии. Том. 184. стр. 1–21. дои : 10.1007/978-3-540-74805-2_1 . ISBN 978-3-540-74804-5 . ПМИД 18064409 .
- ^ Русаков Д.А. (август 2006 г.). «Са2+-зависимые механизмы пресинаптического контроля в центральных синапсах» . Нейробиолог . 12 (4): 317–26. дои : 10.1177/1073858405284672 . ПМЦ 2684670 . ПМИД 16840708 .
- ^ Юмо Ю., Дуссау Ф., Грант Нью-Джерси, Пулен Б. (май 2000 г.). «Как ботулинические и столбнячные нейротоксины блокируют высвобождение нейромедиаторов». Биохимия . 82 (5): 427–46. дои : 10.1016/S0300-9084(00)00216-9 . ПМИД 10865130 .
- ^ Зойдл Г., Дермицель Р. (ноябрь 2002 г.). «В поисках электрического синапса: взгляд в будущее». Исследования клеток и тканей . 310 (2): 137–42. дои : 10.1007/s00441-002-0632-x . ПМИД 12397368 . S2CID 22414506 .
- ^ Бринк П.Р., Кронин К., Раманан С.В. (август 1996 г.). «Щелевые контакты в возбудимых клетках». Журнал биоэнергетики и биомембран . 28 (4): 351–8. дои : 10.1007/BF02110111 . ПМИД 8844332 . S2CID 46371790 .
- ^ Хирш Н.П. (июль 2007 г.). «Нервно-мышечный узел в норме и болезни» . Британский журнал анестезии . 99 (1): 132–8. дои : 10.1093/bja/aem144 . ПМИД 17573397 .
- ^ Хьюз Б.В., Куснер Л.Л., Камински Х.Дж. (апрель 2006 г.). «Молекулярная архитектура нервно-мышечного соединения». Мышцы и нервы . 33 (4): 445–61. дои : 10.1002/mus.20440 . ПМИД 16228970 . S2CID 1888352 .
- ^ Перейти обратно: а б Ньюмарк Дж. (январь 2007 г.). «Нервно-паралитические агенты». Невролог . 13 (1): 20–32. дои : 10.1097/01.nrl.0000252923.04894.53 . ПМИД 17215724 . S2CID 211234081 .
- ^ Коста LG (апрель 2006 г.). «Актуальные проблемы фосфорорганической токсикологии». Клиника Химика Акта; Международный журнал клинической химии . 366 (1–2): 1–13. дои : 10.1016/j.cca.2005.10.008 . ПМИД 16337171 .
- ^ Перейти обратно: а б с Клебер А.Г., Руди Ю. (апрель 2004 г.). «Основные механизмы распространения сердечного импульса и связанные с ними аритмии». Физиологические обзоры . 84 (2): 431–88. doi : 10.1152/physrev.00025.2003 . ПМИД 15044680 . S2CID 21823003 .
- ^ Тамарго Дж., Кабальеро Р., Дельпон Э. (январь 2004 г.). «Фармакологические подходы в лечении мерцательной аритмии». Современная медицинская химия . 11 (1): 13–28. дои : 10.2174/0929867043456241 . ПМИД 14754423 .
- ^ Слейман CL, Лонг WS, Градманн Д. (апрель 1976 г.). « Потенциалы действия» у Neurospora crassa, мицелиального гриба». Biochimica et Biophysical Acta (BBA) – Биомембраны . 426 (4): 732–44. дои : 10.1016/0005-2736(76)90138-3 . ПМИД 130926 .
- ^ Муммерт Х., Градманн Д. (декабрь 1991 г.). «Потенциалы действия в вертлужной впадине: измерение и моделирование потенциалзависимых потоков». Журнал мембранной биологии . 124 (3): 265–73. дои : 10.1007/BF01994359 . ПМИД 1664861 . S2CID 22063907 .
- ^ Градманн Д. (2001). «Модели колебаний растений». Ауст. Дж. Физиол растений . 28 (7): 577–590. дои : 10.1071/pp01017 .
- ^ Бейлби М.Дж. (2007). Потенциал действия у харофитов . Международный обзор цитологии. Том. 257. стр. 43–82. дои : 10.1016/S0074-7696(07)57002-6 . ISBN 978-0-12-373701-4 . ПМИД 17280895 .
- ^ Градманн Д., Хоффштадт Дж. (ноябрь 1998 г.). «Электросвязь переносчиков ионов в растениях: взаимодействие с внутренними концентрациями ионов». Журнал мембранной биологии . 166 (1): 51–9. дои : 10.1007/s002329900446 . ПМИД 9784585 . S2CID 24190001 .
- ^ Фромм Дж., Лотнер С. (март 2007 г.). «Электрические сигналы и их физиологическое значение у растений» . Растение, клетка и окружающая среда . 30 (3): 249–257. дои : 10.1111/j.1365-3040.2006.01614.x . ПМИД 17263772 .
- ^ Лейс С.П., Маки ГО, Мич Р.В. (май 1999 г.). «Импульсная проводимость в губке» . Журнал экспериментальной биологии . 202 (Часть 9) (9): 1139–50. дои : 10.1242/jeb.202.9.1139 . ПМИД 10101111 .
- ^ Кейнс РД (1989). «Роль гигантских аксонов в изучении нервного импульса». Биоэссе . 10 (2–3): 90–3. дои : 10.1002/bies.950100213 . ПМИД 2541698 .
- ^ Менье К., Сегев I (ноябрь 2002 г.). «Игра в адвоката дьявола: полезна ли модель Ходжкина-Хаксли?». Тенденции в нейронауках . 25 (11): 558–63. дои : 10.1016/S0166-2236(02)02278-6 . ПМИД 12392930 . S2CID 1355280 .
- ^ Коул КС (1949). «Динамические электрические характеристики мембраны аксона кальмара». Арх. наук. Физиол . 3 : 253–8.
- ^ Линг Дж., Джерард Р.В. (декабрь 1949 г.). «Нормальный мембранный потенциал портняжных волокон лягушки». Журнал клеточной и сравнительной физиологии . 34 (3): 383–96. дои : 10.1002/jcp.1030340304 . ПМИД 15410483 .
- ^ Настук В.Л., Ходжкин А (1950). «Электрическая активность отдельных мышечных волокон». Журнал клеточной и сравнительной физиологии . 35 : 39–73. дои : 10.1002/jcp.1030350105 .
- ^ Брок Л.Г., Кумбс Дж.С., Экклс Дж.К. (август 1952 г.). «Запись потенциалов мотонейронов внутриклеточным электродом» . Журнал физиологии . 117 (4): 431–60. doi : 10.1113/jphysicalol.1952.sp004759 . ПМЦ 1392415 . ПМИД 12991232 .
- ^ Росс В.Н., Зальцберг Б.М., Коэн Л.Б., Давила Х.В. (декабрь 1974 г.). «Большое изменение поглощения красителя во время потенциала действия» . Биофизический журнал . 14 (12): 983–6. Бибкод : 1974BpJ....14..983R . дои : 10.1016/S0006-3495(74)85963-1 . ПМЦ 1334592 . ПМИД 4429774 .
* Гринкевич Г., Поение М., Цянь Р.Ю. (март 1985 г.). «Новое поколение индикаторов Ca2+ со значительно улучшенными флуоресцентными свойствами» . Журнал биологической химии . 260 (6): 3440–50. дои : 10.1016/S0021-9258(19)83641-4 . ПМИД 3838314 . - ^ Бу Дж., Адамс Х., Бербари Э.Дж., Рубарт М. (март 2009 г.). «Равномерная реполяризация потенциала действия внутри сарколеммы желудочковых кардиомиоцитов in situ» . Биофизический журнал . 96 (6): 2532–46. Бибкод : 2009BpJ....96.2532B . дои : 10.1016/j.bpj.2008.12.3896 . ПМЦ 2907679 . ПМИД 19289075 .
- ^ Миллиган СП, Эдвардс С. (июль 1965 г.). «Некоторые факторы, влияющие на время восстановления контрактурной способности после калиевой контрактуры в поперечно-полосатой мышце лягушки» . Журнал общей физиологии . 48 (6): 975–83. дои : 10.1085/jgp.48.6.975 . ПМК 2195447 . ПМИД 5855511 .
* Ричи Дж. М., Рогарт Р.Б. (1977). «Связывание сакситоксина и тетродотоксина с возбудимыми тканями». Обзоры физиологии, биохимии и фармакологии, том 79 . Том. 79. стр. 1–50. дои : 10.1007/BFb0037088 . ISBN 0-387-08326-Х . ПМИД 335473 .
* Кейнс Р.Д., Ричи Дж.М. (август 1984 г.). «О связывании меченого сакситоксина с гигантским аксоном кальмара». Труды Лондонского королевского общества. Серия Б, Биологические науки . 222 (1227): 147–53. Бибкод : 1984РСПСБ.222..147К . дои : 10.1098/rspb.1984.0055 . ПМИД 6148754 . S2CID 11465181 . - ^ Пикколино М (октябрь 1997 г.). «Луиджи Гальвани и животное электричество: два столетия после основания электрофизиологии». Тенденции в нейронауках . 20 (10): 443–8. дои : 10.1016/S0166-2236(97)01101-6 . ПМИД 9347609 . S2CID 23394494 .
- ^ Пикколино М. (апрель 2000 г.). «Двухсотлетие вольтовой батареи (1800-2000): искусственный электрический орган». Тенденции в нейронауках . 23 (4): 147–51. дои : 10.1016/S0166-2236(99)01544-1 . ПМИД 10717671 . S2CID 393323 .
- ^ Бернштейн Дж. (1902). «Исследования по термодинамике биоэлектрических токов» . Архив Пфлюгера по всей физиологии . 92 (10–12): 521–562. дои : 10.1007/BF01790181 . S2CID 33229139 .
- ^ Коул К.С. , Кертис Х.Дж. (май 1939 г.). «Электрическое сопротивление гигантского аксона кальмара во время активности» . Журнал общей физиологии . 22 (5): 649–70. дои : 10.1085/jgp.22.5.649 . ПМК 2142006 . ПМИД 19873125 .
- ^ Лапик Л. (1907). «Количественные исследования электрического возбуждения нервов, трактуемого как поляризация». Дж. Физиол. Патол. Генерал 9 : 620–635.
- ^ Ходжкин А.Л. , Кац Б. (март 1949 г.). «Влияние ионов натрия на электрическую активность гигантского аксона кальмара» . Журнал физиологии . 108 (1): 37–77. дои : 10.1113/jphysicalol.1949.sp004310 . ПМЦ 1392331 . ПМИД 18128147 .
- ^ Неер Э. , Сакманн Б. (апрель 1976 г.). «Одноканальные токи, записанные с мембраны денервированных мышечных волокон лягушки». Природа . 260 (5554): 799–802. Бибкод : 1976Natur.260..799N . дои : 10.1038/260799a0 . ПМИД 1083489 . S2CID 4204985 .
* Хэмилл О.П., Марти А., Неер Э., Сакманн Б., Сигворт Ф.Дж. (август 1981 г.). «Улучшенные методы патч-зажима для записи тока с высоким разрешением из клеток и патчей бесклеточных мембран». Архив Пфлюгерса . 391 (2): 85–100. дои : 10.1007/BF00656997 . ПМИД 6270629 . S2CID 12014433 .
* Неер Э. , Сакманн Б. (март 1992 г.). «Техника патч-клампа». Научный американец . 266 (3): 44–51. Бибкод : 1992SciAm.266c..44N . doi : 10.1038/scientificamerican0392-44 . ПМИД 1374932 . - ^ Йеллен Дж. (сентябрь 2002 г.). «Потенциал-управляемые калиевые каналы и их родственники». Природа . 419 (6902): 35–42. Бибкод : 2002Natur.419...35Y . дои : 10.1038/nature00978 . ПМИД 12214225 . S2CID 4420877 .
- ^ Дойл Д.А., Мораис Кабрал Дж., Пфюцнер Р.А., Куо А., Гулбис Дж.М., Коэн С.Л. и др. (апрель 1998 г.). «Строение калиевого канала: молекулярные основы К+-проводимости и селективности». Наука . 280 (5360): 69–77. Бибкод : 1998Sci...280...69D . дои : 10.1126/science.280.5360.69 . ПМИД 9525859 .
* Чжоу Ю, Мораис-Кабрал Дж. Х., Кауфман А., Маккиннон Р. (ноябрь 2001 г.). «Химия координации и гидратации ионов, выявленная комплексом K + канал-Fab при разрешении 2,0 А». Природа . 414 (6859): 43–8. Бибкод : 2001Natur.414...43Z . дои : 10.1038/35102009 . ПМИД 11689936 . S2CID 205022645 .
* Цзян Ю, Ли А, Чен Дж, Рута В, Каден М, Чайт БТ, Маккиннон Р (май 2003 г.). «Рентгеновская структура потенциалзависимого K+-канала». Природа . 423 (6935): 33–41. Бибкод : 2003Natur.423...33J . дои : 10.1038/nature01580 . ПМИД 12721618 . S2CID 4347957 . - ^ Ча А., Снайдер Г.Е., Селвин П.Р., Безанилья Ф. (декабрь 1999 г.). «Перемещение в атомном масштабе области измерения напряжения в калиевом канале, измеренное с помощью спектроскопии» . Природа . 402 (6763): 809–13. Бибкод : 1999Natur.402..809C . дои : 10.1038/45552 . ПМИД 10617201 . S2CID 4353978 .
* Глаунер К.С., Маннуццу Л.М., Ганди К.С., Исакофф Э.Ю. (декабрь 1999 г.). «Спектроскопическое картирование движения датчика напряжения в калиевом канале шейкера». Природа . 402 (6763): 813–7. Бибкод : 1999Natur.402..813G . дои : 10.1038/45561 . ПМИД 10617202 . S2CID 4417476 .
* Безанилья Ф (апрель 2000 г.). «Датчик напряжения в потенциалзависимых ионных каналах». Физиологические обзоры . 80 (2): 555–92. дои : 10.1152/physrev.2000.80.2.555 . ПМИД 10747201 . S2CID 18629998 . - ^ Каттералл, Вашингтон (февраль 2001 г.). «Трехмерное изображение натриевых каналов» . Природа . 409 (6823): 988–9, 991. Бибкод : 2001Natur.409..988C . дои : 10.1038/35059188 . ПМИД 11234048 . S2CID 4371677 .
* Сато С., Уэно Ю., Асаи К., Такахаси К., Сато М., Энгель А., Фудзиёси Ю. (февраль 2001 г.). «Потенциал-чувствительный натриевый канал представляет собой колоколообразную молекулу с несколькими полостями». Природа . 409 (6823): 1047–51. Бибкод : 2001Natur.409.1047S . дои : 10.1038/35059098 . ПМИД 11234014 . S2CID 4430165 . - ^ Скоу Дж. К. (февраль 1957 г.). «Влияние некоторых катионов на аденозинтрифосфатазу периферических нервов». Биохимика и биофизика Acta . 23 (2): 394–401. дои : 10.1016/0006-3002(57)90343-8 . ПМИД 13412736 . S2CID 32516710 .
- ^ Ходжкин А.Л. , Кейнс Р.Д. (апрель 1955 г.). «Активный транспорт катионов в гигантских аксонах Sepia и Loligo» . Журнал физиологии . 128 (1): 28–60. doi : 10.1113/jphysicalol.1955.sp005290 . ПМЦ 1365754 . PMID 14368574 .
- ^ Колдуэлл П.С., Ходжкин А.Л., Кейнс Р.Д., Шоу Т.Л. (июль 1960 г.). «Влияние инъекции «богатых энергией» фосфатных соединений на активный транспорт ионов в гигантских аксонах Лолиго» . Журнал физиологии . 152 (3): 561–90. дои : 10.1113/jphysicalol.1960.sp006509 . ПМЦ 1363339 . ПМИД 13806926 .
- ^ Колдуэлл ПК, Кейнс РД (июнь 1957 г.). «Использование энергии фосфатных связей для экструзии натрия из гигантских аксонов». Журнал физиологии . 137 (1): 12–3П. doi : 10.1113/jphysicalol.1957.sp005830 . ПМИД 13439598 . S2CID 222188054 .
- ^ Морт Дж.П., Педерсен Б.П., Тоуструп-Йенсен М.С., Соренсен Т.Л., Петерсен Дж., Андерсен Дж.П. и др. (декабрь 2007 г.). «Кристаллическая структура натриево-калиевого насоса». Природа . 450 (7172): 1043–9. Бибкод : 2007Nature.450.1043M . дои : 10.1038/nature06419 . ПМИД 18075585 . S2CID 4344526 .
- ^ Ли А.Г., Ист-Дж.М. (июнь 2001 г.). «Что структура кальциевого насоса говорит нам о его механизме» . Биохимический журнал . 356 (Часть 3): 665–83. дои : 10.1042/0264-6021:3560665 . ПМЦ 1221895 . ПМИД 11389676 .
- ^ * Фицхью Р. (май 1960 г.). «Пороги и плато в нервных уравнениях Ходжкина-Хаксли» . Журнал общей физиологии . 43 (5): 867–96. дои : 10.1085/jgp.43.5.867 . ПМК 2195039 . ПМИД 13823315 .
* Кеплер Т.Б., Эбботт Л.Ф., Мардер Э. (1992). «Редукция моделей нейронов, основанных на проводимости». Биологическая кибернетика . 66 (5): 381–7. дои : 10.1007/BF00197717 . ПМИД 1562643 . S2CID 6789007 . - ^ Моррис С., Лекар Х. (июль 1981 г.). «Колебания напряжения в гигантском мышечном волокне ракушки» . Биофизический журнал . 35 (1): 193–213. Бибкод : 1981BpJ....35..193M . дои : 10.1016/S0006-3495(81)84782-0 . ПМЦ 1327511 . ПМИД 7260316 .
- ^ Фитцхью Р. (июль 1961 г.). «Импульсы и физиологические состояния в теоретических моделях нервной мембраны» . Биофизический журнал . 1 (6): 445–66. Бибкод : 1961BpJ.....1..445F . дои : 10.1016/S0006-3495(61)86902-6 . ПМЦ 1366333 . ПМИД 19431309 .
* Нагумо Дж., Аримото С., Ёсидзава С. (1962). «Активная линия передачи импульсов, имитирующая нервный аксон». Труды ИРЭ . 50 (10): 2061–2070. дои : 10.1109/JRPROC.1962.288235 . S2CID 51648050 . - ^ Бонхёффер К.Ф. (сентябрь 1948 г.). «Активация пассивного железа как модель возбуждения нерва» . Журнал общей физиологии . 32 (1): 69–91. дои : 10.1085/jgp.32.1.69 . ПМК 2213747 . ПМИД 18885679 .
* Бонхёффер К.Ф. (1953). «Модели нервного возбуждения». естественные науки . 40 (11): 301–311. Бибкод : 1953NW.....40..301B . дои : 10.1007/BF00632438 . S2CID 19149460 .
* Ван дер Поль Б (1926). «О релаксациях-колебаниях». Философский журнал . 2 : 977–992.
* Ван дер Пол Б. , Ван дер Марк Дж (1928). «Сердцебиение рассматривается как релаксационные колебания и электрическая модель сердца». Философский журнал . 6 : 763–775. дои : 10.1080/14786441108564652 .
* Ван дер Пол Б. , ван дер Марк Дж (1929). «Сердцебиение рассматривается как релаксационные колебания и электрическая модель сердца». Арх. Неерл. Физиол . 14 : 418–443. - ^ Эванс Дж.В. (1972). «Уравнения нервных аксонов. I. Линейные приближения» . Университет Индианы. Математика. Дж . 21 (9): 877–885. дои : 10.1512/iumj.1972.21.21071 .
* Эванс Дж.В., Феро Дж. (1977). «Теория локальной устойчивости нервного импульса». Математика. Биосци . 37 : 23–50. дои : 10.1016/0025-5564(77)90076-1 . - ^ Кинер Дж. П. (1983). «Аналоговая схема для уравнений Ван дер Поля и ФитцХью-Нагумо». Транзакции IEEE по системам, человеку и кибернетике . 13 (5): 1010–1014. дои : 10.1109/TSMC.1983.6313098 . S2CID 20077648 .
- ^ Хупер С.Л. (март 2000 г.). «Генератор центральных шаблонов». Современная биология . 10 (5): 176–179 Р. Бибкод : 2000CBio...10.R176H . CiteSeerX 10.1.1.133.3378 . дои : 10.1016/S0960-9822(00)00367-5 . ПМИД 10713861 . S2CID 11388348 .
Книги
[ редактировать ]- Андерсон Дж.А., Розенфельд Э., ред. (1988). Нейрокомпьютеры: основы исследований . Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-01097-9 . LCCN 87003022 . OCLC 15860311 .
- Бернштейн Дж. (1912). Электробиология, наука об электрических процессах в организме, представленная на . современной основе Брауншвейг: Видег и сын. LCCN12027986 . OCLC 11358569 .
- Бауэр Дж. М., Биман Д. (1995). Книга GENESIS: исследование реалистичных нейронных моделей с помощью системы GENERAL NEural SIMulation . Санта-Клара, Калифорния: ТЕЛОС. ISBN 978-0-387-94019-9 . LCCN 94017624 . OCLC 30518469 .
- Мангал М.А. (1961). История электрической активности мозга . Лондон: Питман. LCCN 62001407 . OCLC 556863 .
- Буллок Т.Х. , Хорридж Г.А. (1965). Структура и функции нервной системы беспозвоночных . Серия книг по биологии. Сан-Франциско: WH Freeman. LCCN 65007965 . OCLC 558128 .
- Буллок Т.Х. , Орканд Р., Гриннелл А. (1977). Введение в нервную систему . Серия книг по биологии. Сан-Франциско: WH Freeman. ISBN 978-0-7167-0030-2 . LCCN 76003735 . ОСЛК 2048177 .
- Филд Дж, изд. (1959). Справочник по физиологии: критическое, всестороннее изложение физиологических знаний и концепций: Раздел 1: Нейрофизиология . Том. 1. Вашингтон, округ Колумбия: Американское физиологическое общество. LCCN 60004587 . OCLC 830755894 .
- Ганонг, ВФ (1991). Обзор медицинской физиологии (15-е изд.). Норуолк, Коннектикут: Эпплтон и Ланге. ISBN 978-0-8385-8418-7 . ISSN 0892-1253 . LCCN 87642343 . ОСЛК 23761261 .
{{cite book}}:|journal=игнорируется ( помогите ) - Гукенхаймер Дж., Холмс П., ред. (1986). Нелинейные колебания, динамические системы и бифуркации векторных полей . Прикладные математические науки. Том. 42 (2-е изд.). Нью-Йорк: Springer Verlag. ISBN 978-0-387-90819-9 . OCLC 751129941 .
- Хоппенстедт (1986). Введение в математику нейронов . Кембриджские исследования по математической биологии. Том. 6. Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-31574-6 . LCCN 85011013 . OCLC 12052275 .
- Юнге Д. (1981). Нервное и мышечное возбуждение (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-410-2 . LCCN 80018158 . OCLC 6486925 .
- Кеттенманн Х., Грантин Р., ред. (1992). Практические электрофизиологические методы: руководство для исследований in vitro в области нейробиологии позвоночных . Нью-Йорк: Уайли. ISBN 978-0-471-56200-9 . LCCN 92000179 . ОСЛК 25204689 .
- Кейнс Р.Д. , Эйдли DJ (1991). Нерв и мышца (2-е изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-41042-7 . LCCN 90015167 . ОСЛК 25204483 .
- Кох С. , Сегев И., ред. (1989). Методы нейронного моделирования: от синапсов к сетям . Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-11133-1 . LCCN 88008279 . OCLC 18384545 .
- Лавалле М., Шанн О.Ф., Эбер Н.К., ред. (1969). Стеклянные микроэлектроды . Нью-Йорк: Уайли. ISBN 978-0-471-51885-3 . LCCN 68009252 . ОСЛК 686 .
- Маккалок WS (1988). Воплощения разума . Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-63114-3 . LCCN 88002987 . ОСЛК 237280 .
- МакГенри LC, Гаррисон FH (1969). История неврологии Гаррисона . Спрингфилд, Иллинойс: Чарльз К. Томас. OCLC 429733931 .
- Сильверторн ДУ (2010). Физиология человека: комплексный подход (5-е изд.). Сан-Франциско: Пирсон. ISBN 978-0-321-55980-7 . LCCN 2008050369 . OCLC 268788623 .
- Спэнсвик Р.М., Лукас У.Дж., Дэйнти Дж., ред. (1980). Мембранный транспорт растений: актуальные концептуальные проблемы . Развитие биологии растений. Том. 4. Амстердам: Elsevier Biomedical Press. ISBN 978-0-444-80192-0 . LCCN 79025719 . OCLC 5799924 .
- Первес Д., Августин Г.Дж., Фицпатрик Д., Холл В.К., Ламантия А.С., Макнамара Дж.О., Уильямс С.М. (2001). «Высвобождение передатчиков из синаптических везикул» . Нейронаука (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-742-4 . LCCN 00059496 . OCLC 806472664 .
- Первс Д., Августин Г.Дж., Фицпатрик Д., Холл В.К., Ламантия А.С., Макнамара Дж.О., Уайт Л.Е. (2008). Нейронаука (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-697-7 . LCCN 2007024950 . OCLC 144771764 .
- Рик Г.Н., Познански Р.Р., Спорнс О., Розенберг Дж.Р., Линдси К.А., ред. (2005). Моделирование в нейронауках: от биологических систем к нейромиметической робототехнике . Бока-Ратон, Флорида: Тейлор и Фрэнсис. ISBN 978-0-415-32868-5 . LCCN 2005298022 . ОСЛК 489024131 .
- Шмидт-Нильсен К. (1997). Физиология животных: адаптация и окружающая среда (5-е изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-57098-5 . LCCN 96039295 . OCLC 35744403 .
- Шванн HP, изд. (1969). Биологическая инженерия . Межвузовская серия по электронике. Том. 9. Нью-Йорк: МакГроу-Хилл. ISBN 978-0-07-055734-5 . LCCN 68027513 . ОСЛК 51993 .
- Стивенс CF (1966). Нейрофизиология: Учебник . Нью-Йорк: Джон Уайли и сыновья. ISBN 9780471824367 . LCCN 66015872 . ОСЛК 1175605 .
- Ваксман С.Г., изд. (2007). Молекулярная неврология . Берлингтон, Массачусетс: Elsevier Academic Press. ISBN 978-0-12-369509-3 . LCCN 2008357317 . OCLC 154760295 .
- Уорден Ф.Г., Свази Дж.П., Адельман Дж., ред. (1975). Нейронауки, пути открытий . Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-23072-8 . LCCN 75016379 . ОСЛК 1500233 .
Веб-страницы
[ редактировать ]- ^ Фитцхью Р., Ижикевич Э. (2006). «Модель ФитцХью-Нагумо» . Схоларпедия . 1 (9): 1349. Бибкод : 2006SchpJ...1.1349I . дои : 10.4249/scholarpedia.1349 .
- ^ «Нобелевская премия по физиологии и медицине 1963 года» (пресс-релиз). Шведская королевская академия наук. 1963. Архивировано из оригинала 16 июля 2007 года . Проверено 21 февраля 2010 г.
- ^ «Нобелевская премия по физиологии и медицине 1991 г.» (пресс-релиз). Шведская королевская академия наук. 1991. Архивировано из оригинала 24 марта 2010 года . Проверено 21 февраля 2010 г.
- ^ «Нобелевская премия по физиологии и медицине 1906 года» (пресс-релиз). Шведская королевская академия наук. 1906. Архивировано из оригинала 4 декабря 2008 года . Проверено 21 февраля 2010 г.
- ^ Варлоу С. (июнь 2007 г.). «Недавняя эволюция симбиотического ионного канала в семействе бобовых изменила ионную проводимость и улучшила функциональность передачи сигналов кальция» . Практическая неврология . 7 (3). Издательская группа BMJ: 192–197. Архивировано из оригинала 14 марта 2012 года . Проверено 23 марта 2013 г.
- ^ «Нобелевская премия по химии 1997 г.» (Пресс-релиз). Шведская королевская академия наук. 1997. Архивировано из оригинала 23 октября 2009 года . Проверено 21 февраля 2010 г.
Дальнейшее чтение
[ редактировать ]- Эйдли DJ, Стэнфилд PR (1996). Ионные каналы: молекулы в действии . Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-49882-1 .
- Медведь М.Ф., Коннорс Б.В., Парадизо М.А. (2001). Нейронаука: исследование мозга . Балтимор: Липпинкотт. ISBN 0-7817-3944-6 .
- Клэй-младший (май 2005 г.). «Возвращение к аксональной возбудимости» . Прогресс биофизики и молекулярной биологии . 88 (1): 59–90. doi : 10.1016/j.pbiomolbio.2003.12.004 . ПМИД 15561301 .
- Дойч С., Микели-Цанаку Э (1987). Нейроэлектрические системы . Нью-Йорк: Издательство Нью-Йоркского университета. ISBN 0-8147-1782-9 .
- Хилле Б (2001). Ионные каналы возбудимых мембран (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-321-1 .
- Джонстон Д., Ву С.М. (1995). Основы клеточной нейрофизиологии . Кембридж, Массачусетс: Брэдфордская книга, MIT Press. ISBN 0-262-10053-3 .
- Кандель Э.Р. , Шварц Дж.Х., Джесселл Т.М. (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 0-8385-7701-6 .
- Миллер С. (1987). «Как работают белки ионных каналов». В Качмарек Л.К., Левитан И.Б. (ред.). Нейромодуляция: биохимический контроль возбудимости нейронов . Нью-Йорк: Издательство Оксфордского университета. стр. 39–63. ISBN 978-0-19-504097-5 .
- Нельсон Д.Л., Кокс М.М. (2008). Ленингерские принципы биохимии (5-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-7108-1 .
Внешние ссылки
[ редактировать ]- Ионный поток в потенциалах действия в Blackwell Publishing
- Распространение потенциала действия в миелинизированных и немиелиновых аксонах в Blackwell Publishing.
- Генерация AP в сердечных клетках и генерация AP в нейронных клетках
- Мембранный потенциал покоя из книги «Жизнь: наука о биологии» , У.К. Первс, Д. Садава, Г.Х. Орианс и Х.К. Хеллер, 8-е издание, Нью-Йорк: У.Х. Фриман, ISBN 978-0-7167-7671-0 .
- Ионное движение и напряжение Гольдмана для произвольных концентраций ионов в Университете Аризоны.
- Мультфильм, иллюстрирующий потенциал действия
- Распространение потенциала действия
- Создание потенциала действия: моделирование ограничения напряжения и тока [ постоянная мертвая ссылка ]
- Программное обеспечение с открытым исходным кодом для моделирования потенциалов действия нейронов и сердца на SourceForge.net
- Введение в потенциал действия , Neuroscience Online (электронный учебник по нейробиологии Медицинской школы UT в Хьюстоне)
- Академия Хана: электротоник и потенциал действия. Архивировано 2 июля 2014 г. в Wayback Machine.
| Базы данных органов управления : Национальные |
|---|