Количественные модели потенциала действия
В нейрофизиологии несколько математических моделей потенциала действия разработано , которые делятся на два основных типа. Первый тип направлен на количественное моделирование экспериментальных данных, т.е. на точное воспроизведение измерений тока и напряжения. Известная Ходжкина-Хаксли модель аксона кальмара Лолиго является примером таких моделей. [1] Хотя модель HH качественно правильна, она не описывает точно каждый тип возбудимой мембраны, поскольку она рассматривает только два иона (натрия и калия), каждый из которых имеет только один тип потенциал-чувствительного канала. Однако другие ионы, такие как кальций, могут быть важны, и для всех ионов существует большое разнообразие каналов. [2] Например, сердечный потенциал действия иллюстрирует, как потенциалы действия различной формы могут генерироваться на мембранах с потенциал-чувствительными кальциевыми каналами и различными типами натриевых/калиевых каналов. Второй тип математической модели является упрощением первого типа; цель — не воспроизвести экспериментальные данные, а качественно понять роль потенциалов действия в нейронных цепях. Для этой цели подробные физиологические модели могут оказаться излишне сложными и затмить «лес за деревьями». Модель ФитцХью-Нагумо типична для этого класса, который часто изучается на предмет его поведения вовлекания . [3] Увлечение обычно наблюдается в природе, например, при синхронном освещении светлячков , которое координируется вспышкой потенциалов действия; [4] унос также можно наблюдать в отдельных нейронах. [5] Оба типа моделей могут использоваться для понимания поведения небольших биологических нейронных сетей , таких как центральные генераторы шаблонов, ответственные за некоторые автоматические рефлекторные действия. [6] Такие сети могут генерировать сложную временную структуру потенциалов действия, которая используется для координации мышечных сокращений, например, тех, которые участвуют в дыхании или быстром плавании, чтобы спастись от хищника. [7]
Модель Ходжкина – Хаксли
[ редактировать ]
В 1952 году Алан Ллойд Ходжкин и Эндрю Хаксли разработали набор уравнений, соответствующих их экспериментальным данным о фиксации напряжения на аксональной мембране. [1] [8] Модель предполагает, что емкость мембраны C постоянна; таким образом, трансмембранное напряжение V изменяется с общим трансмембранным током I tot согласно уравнению

где I Na , I K и I L — токи, проходящие через локальные натриевые каналы, калиевые каналы и каналы «утечки» (объемные каналы) соответственно. Начальный член I ext представляет собой ток, поступающий от внешних источников, таких как возбуждающие постсинаптические потенциалы от дендритов или электрода ученого.
Модель далее предполагает, что данный ионный канал либо полностью открыт, либо закрыт; если он закрыт, его проводимость равна нулю, тогда как если открыт, его проводимость равна некоторому постоянному значению g . Следовательно, чистый ток через ионный канал зависит от двух переменных: вероятности p открытия канала и разницы напряжения от равновесного напряжения этого иона, V - V eq . Например, ток через калиевый канал можно записать как

что эквивалентно закону Ома . По определению, чистый ток не течет ( I K = 0), когда трансмембранное напряжение равно равновесному напряжению этого иона (когда V = E K ).
Чтобы точно соответствовать своим данным, Ходжкин и Хаксли предположили, что каждый тип ионного канала имеет несколько «ворот», так что канал открыт, только если все ворота открыты, и закрыты в противном случае. Они также предположили, что вероятность открытия ворот не зависит от открытия других ворот; позже это предположение было подтверждено для ворот инактивации. [9] Ходжкин и Хаксли смоделировали потенциал-чувствительный калиевый канал как имеющий четыре ворота; если p n обозначает вероятность открытия одного такого вентиля, то вероятность открытия всего канала является произведением четырех таких вероятностей, т. е. p открыто, K = n 4 . Аналогичным образом, вероятность потенциал-чувствительного натриевого канала была смоделирована так, чтобы иметь три одинаковых вентиля с вероятностью m и четвертый вентиль, связанный с инактивацией, с вероятностью h ; таким образом, p open, Na = m 3 ч . Предполагается, что вероятности для каждых ворот подчиняются кинетике первого порядка.

где как равновесное значение m eq, так и постоянная времени релаксации τ m зависят от мгновенного напряжения V на мембране. Если V изменяется во временном масштабе медленнее, чем τ m , вероятность m всегда будет примерно равна равновесному значению m eq ; однако если V изменяется быстрее, то m будет отставать от m eq . Сопоставив данные фиксации напряжения, Ходжкин и Хаксли смогли смоделировать, как эти равновесные значения и константы времени меняются в зависимости от температуры и трансмембранного напряжения. [1] Формулы сложны и экспоненциально зависят от напряжения и температуры. Например, постоянная времени вероятности активации натриевых каналов h изменяется как 3 (θ−6,3)/10 с температурой Цельсия θ и напряжением V как

Таким образом, уравнения Ходжкина-Хаксли представляют собой сложные нелинейные обыкновенные дифференциальные уравнения с четырьмя независимыми переменными : трансмембранным напряжением V и вероятностями m , h и n . [10] Общего решения этих уравнений не обнаружено. Менее амбициозный, но общеприменимый метод исследования таких нелинейных динамических систем — рассмотрение их поведения вблизи фиксированной точки . [11] Этот анализ показывает, что система Ходжкина–Хаксли претерпевает переход от устойчивого покоя к пачкам возбуждающего тока I ext колебаний при постепенном увеличении ; Примечательно, что аксон снова становится стабильно неподвижным по мере дальнейшего увеличения стимулирующего тока. [12] Также было проведено более общее исследование типов качественного поведения аксонов, предсказываемых уравнениями Ходжкина–Хаксли. [10]
Модель ФитцХью – Нагумо
[ редактировать ]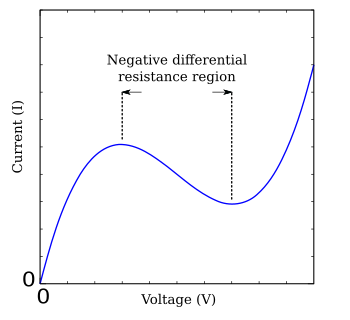
Из-за сложности уравнений Ходжкина – Хаксли были разработаны различные упрощения, которые демонстрируют качественно подобное поведение. [3] [13] Модель ФитцХью-Нагумо является типичным примером такой упрощенной системы. [14] [15] Модель FHN, основанная на туннельном диоде , имеет только две независимые переменные, но демонстрирует поведение стабильности, аналогичное полным уравнениям Ходжкина – Хаксли. [16] Уравнения


где g(V) — функция напряжения V , имеющая область отрицательного наклона в середине, окруженную одним максимумом и одним минимумом (рис. FHN). Хорошо изученным простым случаем модели ФитцХью–Нагумо является модель нерва Бонхёффера–ван дер Поля, которая описывается уравнениями [17]


где коэффициент ε предполагается малым. Эти уравнения можно объединить в дифференциальное уравнение второго порядка.

Это уравнение Ван дер Поля стимулировало множество исследований в области математики нелинейных динамических систем . Схемы операционных усилителей , реализующие модели потенциала действия ФХН и Ван дер Поля, были разработаны Кинером. [18]
Гибрид моделей Ходжкина-Хаксли и ФитцХью-Нагумо был разработан Моррисом и Лекаром в 1981 году и применен к мышечным волокнам ракообразных . [19] Верная физиологии ракушек, модель Морриса-Лекара заменяет потенциалзависимый натриевый ток модели Ходжкина-Хаксли зависимым от напряжения кальциевым током. Инактивации нет (нет переменной h ), и кальциевый ток мгновенно уравновешивается, так что опять же, есть только две зависящие от времени переменные: трансмембранное напряжение V и вероятность калиевых ворот n . Взрыв, унос и другие математические свойства этой модели были подробно изучены. [20]
Простейшими моделями потенциала действия являются модели «смыва и заполнения» (также называемые моделями «интеграции и запуска»), в которых входной сигнал суммируется (фаза «заполнения») до тех пор, пока не достигнет порогового значения, запуская импульс и обнуление суммирования (фаза «промывки»). [3] [21] [22] Все эти модели способны проявлять увлечение , которое обычно наблюдается в нервных системах. [3]
Внеклеточные потенциалы и токи
[ редактировать ]В то время как приведенные выше модели моделируют трансмембранное напряжение и ток на одном участке мембраны, другие математические модели относятся к напряжениям и токам в ионном растворе, окружающем нейрон. [23] Такие модели полезны при интерпретации данных с внеклеточных электродов, которые были распространены до изобретения стеклянных электродов-пипеток, которые позволяли осуществлять внутриклеточную запись. [24] Внеклеточная среда может быть смоделирована как нормальный изотропный ионный раствор ; в таких растворах ток следует линиям электрического поля в соответствии с континуальной формой закона Ома.

где j и E — векторы, представляющие плотность тока и электрическое поле соответственно, и где σ — проводимость . Таким образом, j можно найти из E , которое, в свою очередь, можно найти с помощью уравнений Максвелла . Уравнения Максвелла можно свести к относительно простой задаче электростатики , поскольку концентрации ионов изменяются слишком медленно (по сравнению со скоростью света ), чтобы магнитные эффекты были важны. Электрический потенциал φ( x ) в любой внеклеточной точке x можно решить, используя тождества Грина [23]
![{\displaystyle \phi (\mathbf {x}) = {\frac {1}{4\pi \sigma _ {\mathrm {снаружи} }}}\oint _ {\mathrm {мембрана} }{\frac {\ частичный }{\partial n}}{\frac {1}{\left|\mathbf {x} -{\boldsymbol {\xi }}\right|}}\left[\sigma _{\mathrm {outside} } \phi _{\mathrm {снаружи} }({\boldsymbol {\xi }})-\sigma _{\mathrm {внутри} }\phi _{\mathrm {внутри} }({\boldsymbol {\xi }} )\вправо]dS}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2c0404ac20c90ec72259f14abc2b5590a76bfb85)
где интеграция происходит по всей поверхности мембраны; — положение на мембране, σ внутри и φ внутри — проводимость и потенциал непосредственно внутри мембраны, а σ снаружи и φ снаружи — соответствующие значения сразу за пределами мембраны. Таким образом, учитывая эти значения σ и φ на мембране, внеклеточный потенциал φ( x ) можно рассчитать для любого положения x ; в свою очередь, электрическое поле E и плотность тока j можно рассчитать из этого потенциального поля. [25]

См. также
[ редактировать ]- Модели биологических нейронов
- Уравнение тока GHK
- Модели нейронных вычислений
- Сальтаторная проводимость
- Биоэлектроника
- Теория кабеля
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Ходжкин А.Л. , Хаксли А.Ф. , Кац Б. (1952). «Токи, переносимые ионами натрия и калия через мембрану гигантского аксона Лолиго» . Журнал физиологии . 116 (4): 424–448. дои : 10.1113/jphysicalol.1952.sp004717 . ПМЦ 1392213 . ПМИД 14946713 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
Ходжкин А.Л. , Хаксли А.Ф. (1952). «Токи, переносимые ионами натрия и калия через мембрану гигантского аксона Лолиго» . Журнал физиологии . 116 (4): 449–472. дои : 10.1113/jphysicalol.1952.sp004717 . ПМЦ 1392213 . ПМИД 14946713 .
Ходжкин А.Л. , Хаксли А.Ф. (1952). «Компоненты мембранной проводимости гигантского аксона Лолиго» . Дж Физиол . 116 (4): 473–496. doi : 10.1113/jphysicalol.1952.sp004718 . ПМК 1392209 . ПМИД 14946714 .
Ходжкин А.Л. , Хаксли А.Ф. (1952). «Двойное влияние мембранного потенциала на натриевую проводимость в гигантском аксоне Лолиго» . Дж Физиол . 116 (4): 497–506. doi : 10.1113/jphysicalol.1952.sp004719 . ПМЦ 1392212 . ПМИД 14946715 .
Ходжкин А.Л. , Хаксли А.Ф. (1952). «Количественное описание мембранного тока и его применение к проводимости и возбуждению нерва» . Дж Физиол . 117 (4): 500–544. doi : 10.1113/jphysicalol.1952.sp004764 . ПМК 1392413 . ПМИД 12991237 . - ^ Ямада В.М., Кох С. , Адамс П.Р. (1989). «Множественные каналы и динамика кальция». В К. Кох , И Сегев (ред.). Методы нейронного моделирования: от синапсов к сетям . Кембридж, Массачусетс: Брэдфордская книга, MIT Press. стр. 97–133. ISBN 978-0-262-11133-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б с д Хоппенстедт (1986). Введение в математику нейронов . Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-31574-6 .
- ^ Хэнсон, FE; Кейс, Дж. Ф.; Бак, Э.; Бак, Дж. (1971). «Синхронность и мгновенное вовлечение у светлячка Новой Гвинеи». Наука . 174 (4005): 161–164. Бибкод : 1971Sci...174..161H . дои : 10.1126/science.174.4005.161 . ПМИД 17742039 . S2CID 41760422 .
- ^ Гуттман Р., Фельдман Л., Якобссон Э. (1980). «Частота уноса аксона кальмара». Дж. Член. Биол . 56 (1): 9–18. дои : 10.1007/BF01869347 . ПМИД 7441721 . S2CID 10775478 .
- ^ Получение ПА (1989). «Реконструкция малых нейронных сетей». В C Кох и я Сегев (ред.). Методы нейронного моделирования: от синапсов к сетям . Кембридж, Массачусетс: Брэдфордская книга, MIT Press. стр. 171–194. ISBN 978-0-262-11133-1 .
- ^ Хупер, Скотт Л. «Генератор центральных шаблонов». Эмбриональный ELS (1999) http://www.els.net/elsonline/figpage/I0000206.html [ постоянная мертвая ссылка ] (2 из 2) [06.02.2001 11:42:28] Онлайн: по состоянию на 27 ноября 2007 г. [1] .
- ^ Нельсон М.Е., Ринзель Дж. (1994). «Модель Ходжкина – Хаксли» (PDF) . В Бауэр Дж., Биман Д. (ред.). Книга GENESIS: исследование реалистичных нейронных моделей с помощью системы GENERAL NEural SIMulation . Нью-Йорк: Springer Verlag. стр. 29–49.
- ^ Армстронг CM, Безанилья Ф, Рохас Э (1973). «Разрушение инактивации натриевой проводимости в аксонах кальмаров, перфузированных проназой» . Дж. Генерал Физиол . 62 (4): 375–391. дои : 10.1085/jgp.62.4.375 . ПМК 2226121 . ПМИД 4755846 .
Рохас Э, Руди Б (1976). «Разрушение инактивации натриевой проводимости специфической протеазой в перфузируемых нервных волокнах Лолиго » . Дж. Физиол . 262 (2): 501–531. дои : 10.1113/jphysicalol.1976.sp011608 . ПМК 1307656 . ПМИД 994046 . - ^ Перейти обратно: а б Сато С., Фукаи Х., Номура Т., Дой С. (2005). «Бифуркационный анализ уравнений Ходжкина – Хаксли». Рике Г.Н., Познански Р.Р., Линдси К.А., Розенберг Дж.Р., Спорнс О (ред.). Моделирование в нейронауках: от биологических систем к нейромиметической робототехнике (2-е изд.). Бока-Ратон: CRC Press. стр. 459–478. ISBN 978-0-415-32868-5 .
- ^ Гукенхаймер Дж. , Холмс П. (1986). Нелинейные колебания, динамические системы и бифуркации векторных полей (2-е издание, переработанное и исправленное изд.). Нью-Йорк: Springer Verlag. стр. 12–16. ISBN 978-0-387-90819-9 .
- ^ Сабах Н.Х., Спенглер Р.А. (1970). «Повторяющийся ответ модели Ходжкина-Хаксли для гигантского аксона кальмара». Журнал теоретической биологии . 29 (2): 155–171. Бибкод : 1970JThBi..29..155S . дои : 10.1016/0022-5193(70)90017-2 . ПМИД 5500466 .
Эванс Дж.В. (1972). «Уравнения нервных аксонов. I. Линейные приближения» . Университет Индианы. Математика. Дж . 21 (9): 877–885. дои : 10.1512/iumj.1972.21.21071 .
Эванс Дж.В., Феро Дж. (1977). «Теория локальной устойчивости нервного импульса». Математика. Биосци . 37 (1–2): 23–50. дои : 10.1016/0025-5564(77)90076-1 . - ^ ФитцХью Р. (1960). «Пороги и плато в уравнениях нервов Ходжкина-Хаксли» . Дж. Генерал Физиол . 43 (5): 867–896. дои : 10.1085/jgp.43.5.867 . ПМК 2195039 . ПМИД 13823315 .
Кеплер Т.Б., Эбботт Л.Ф., Мардер Э. (1992). «Редукция моделей нейронов, основанных на проводимости». Биологическая кибернетика . 66 (5): 381–387. дои : 10.1007/BF00197717 . ПМИД 1562643 . S2CID 6789007 . - ^ ФитцХью Р. (1961). «Импульсы и физиологические состояния в теоретических моделях нервной мембраны» . Биофизический журнал . 1 (6): 445–466. Бибкод : 1961BpJ.....1..445F . дои : 10.1016/S0006-3495(61)86902-6 . ПМЦ 1366333 . ПМИД 19431309 .
- ^ Нагумо Дж., Аримото С., Ёсидзава С. (1962). «Активная линия передачи импульсов, имитирующая нервный аксон». Труды ИРЭ . 50 (10): 2061–2070. дои : 10.1109/JRPROC.1962.288235 . S2CID 51648050 .
- ^ ФитцХью Р. (1969). «Математические модели возбуждения и распространения в нерве». В HP Шванн (ред.). Биологическая инженерия . Нью-Йорк: МакГроу-Хилл. стр. 1–85.
- ^ Бонхёффер К.Ф. (1948). «АКТИВАЦИЯ ПАССИВНОГО ЖЕЛЕЗА КАК МОДЕЛЬ ВОЗБУЖДЕНИЯ НЕРВА» . Дж. Генерал Физиол . 32 (1): 69–91. дои : 10.1085/jgp.32.1.69 . ПМК 2213747 . ПМИД 18885679 .
Бонхёффер К.Ф. (1953). «Модели нервного возбуждения». естественные науки . 40 (11): 301–311. Бибкод : 1953NW.....40..301B . дои : 10.1007/BF00632438 . S2CID 19149460 .
ван дер Поль Б (1926). «О релаксациях-колебаниях». Философский журнал . 2 : 978–992.
ван дер Поль Б , ван дер Марк Дж (1928). «Сердцебиение рассматривается как релаксационные колебания и электрическая модель сердца». Философский журнал . 6 : 763–775.
ван дер Пол Б. , ван дер Марк Дж (1929). «Сердцебиение рассматривается как релаксационные колебания и электрическая модель сердца». Арх. Неерл. Физиол . 14 : 418–443. - ^ Кинер Дж. П. (1983). «Аналоговая схема для уравнений Ван дер Поля и ФитцХью-Нагумо». Транзакции IEEE по системам, человеку и кибернетике . 13 (5): 1010–1014. дои : 10.1109/TSMC.1983.6313098 . S2CID 20077648 .
- ^ Моррис С., Лекар Х. (1981). «Колебания напряжения в гигантском мышечном волокне ракушки» . Биофизический журнал . 35 (1): 193–213. Бибкод : 1981BpJ....35..193M . дои : 10.1016/S0006-3495(81)84782-0 . ПМЦ 1327511 . ПМИД 7260316 .
- ^ Ринцель Дж., Эрментраут ГБ (1989). «Анализ нервной возбудимости и колебаний». В К. Кох , И Сегев (ред.). Методы нейронного моделирования: от синапсов к сетям . Кембридж, Массачусетс: Брэдфордская книга, MIT Press. стр. 135–169. ISBN 978-0-262-11133-1 .
- ^ Кинер Дж. П., Хоппенстедт, Ринзель Дж. (1981). «Интегрированные модели реакции нервной мембраны на колебательный сигнал». SIAM Journal по прикладной математике . 41 (3): 503–517. дои : 10.1137/0141042 .
- ^ Цейнар, Павел; Вышата, Олдржих; Кукал, Яромир; Беранек, Мартин; Валиш, Мартин; Прохазка, Алеш (2020). «Простая модель возбуждающего и тормозного нейрона с конденсаторным переключателем, все части которой биологически объяснены, позволяет вводить хаотические колебания, зависящие от схемы огня» . Научные отчеты . 10 (1): 7353. Бибкод : 2020NatSR..10.7353C . дои : 10.1038/s41598-020-63834-7 . ПМК 7192907 . ПМИД 32355185 .
- ^ Перейти обратно: а б Стивенс, Чарльз Ф. (1966). Нейрофизиология: Учебник . Нью-Йорк: Джон Уайли и сыновья. стр. 161–173 . LCCN 66015872 .
- ^ Линг Дж., Джерард Р.В. (1949). «Нормальный мембранный потенциал портняжных волокон лягушки». Дж. Селл. Комп. Физиол . 34 (3): 383–396. дои : 10.1002/jcp.1030340304 . ПМИД 15410483 .
- ^ Лоренте де Но Р (1947). «Исследование физиологии нервов». Стад. Рокфеллеровский институт. Мед. Исследовать . 132 : Глава. 16.
Мауро А (1960). «Свойства тонких генераторов, связанные с электрофизиологическими потенциалами в объемных проводниках». Дж. Нейрофизиология . 23 (2): 132–143. дои : 10.1152/jn.1960.23.2.132 .
Вудбери Дж.В. (1965). «Глава 3: Потенциалы в объемном проводнике». В ТЦ Рух; HD Паттон (ред.). Физиология и биофизика . Филадельфия: WB Saunders Co.
Дальнейшее чтение
[ редактировать ]- Гласс Л., Макки MC (1988). От часов к хаосу: ритмы жизни . Принстон, Нью-Джерси: Принстонский университет. ISBN 978-0-691-08496-1 .