Потенциал кардиостимулятора
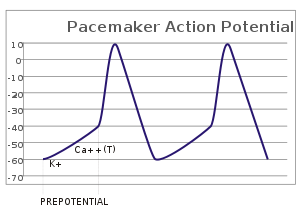
В пейсмекерных клетках сердца ( например, синоатриальном узле ) пейсмекерный потенциал (также называемый пейсмекерным током ) представляет собой медленное положительное увеличение напряжения на клеточной мембране ( мембранный потенциал ), которое происходит между концом один потенциал действия и начало следующего потенциала действия. Это увеличение мембранного потенциала является причиной того, что клеточная мембрана , которая обычно поддерживает мембранный потенциал покоя около -65 мВ, [1] достичь порогового потенциала и, следовательно, запустить следующий потенциал действия; таким образом, пейсмекерный потенциал – это то, что управляет самогенерируемым ритмическим возбуждением ( автоматизмом ) пейсмекерных клеток, а скорость изменения (т. е. наклон) пейсмекерного потенциала – это то, что определяет время следующего потенциала действия и, таким образом, внутренний потенциал действия. Скорость стрельбы ячейки. В здоровом синоатриальном узле (САН, сложная ткань в правом предсердии, содержащая пейсмекерные клетки, которые обычно определяют собственную частоту пульсации всего сердца) [2] [3] ), потенциал кардиостимулятора является основным фактором, определяющим частоту сердечных сокращений. Поскольку потенциал кардиостимулятора представляет собой время отсутствия сокращений между сердечными ударами ( диастолу ), его также называют диастолической деполяризацией .Величина чистого входящего тока, необходимого для перемещения потенциала клеточной мембраны во время пейсмекерной фазы, чрезвычайно мала, порядка нескольких пАм, но этот чистый поток возникает время от времени, изменяя вклад нескольких токов, которые протекают с разным напряжением и временной зависимостью. . Доказательства в поддержку активного присутствия K + , Как 2+ , уже + каналы и Na + /К + В литературе сообщалось по-разному во время кардиостимуляторной фазы, но некоторые указания указывают на то, что «забавный» (I f ) ток является одним из наиболее важных. [4] (см. смешной ток ). В настоящее время имеются существенные доказательства того, что саркоплазматический ретикулум (SR) Ca 2+ -транзиенты участвуют в генерации диастолической деполяризации посредством процесса с участием Na-Ca-обменника.
Ритмическая активность некоторых нейронов, таких как пре-Бетцингеровский комплекс , модулируется нейротрансмиттерами и нейропептидами, и такая модуляционная связь придает нейронам необходимую пластичность для генерации отличительных, зависящих от состояния ритмических паттернов, которые зависят от потенциалов пейсмейкера. [5]
Кардиостимуляторы
[ редактировать ]
Сердце имеет несколько кардиостимуляторов, каждый из которых срабатывает со своей собственной частотой:
- Узел SA: 60–100 ударов в минуту.
- Атриовентрикулярный узел (АВН): 40–60 ударов в минуту.
- Волокна Пуркинье: 20–40 ударов в минуту.
Потенциалы обычно перемещаются в порядке
СА-узел → Атриовентрикулярный узел → Волокна Пуркинье
Обычно все фокусы в конечном итоге срабатывают со скоростью узла SA, а не с их собственной частотой, что является явлением, известным как подавление перегрузки. Таким образом, в нормальном, здоровом сердце наблюдается только собственная частота узла SA.
Патология
[ редактировать ]Однако при патологических состояниях внутренняя скорость становится очевидной. Рассмотрим сердечный приступ, повреждающий область сердца между SA-узлом и AV-узлом.
Узел SA → |блок| АВ-узел → волокна Пуркинье
Другие фокусы не увидят срабатывание узла SA; однако они увидят предсердные очаги. Сердце теперь будет биться с собственной частотой АВ-узла.
Индукция
[ редактировать ]Запуск клеток-водителей ритма индуцируется электрически при достижении порогового потенциала клеточной мембраны. Пороговый потенциал — это потенциал, которого должна достичь возбудимая клеточная мембрана, такая как миоцит , чтобы вызвать потенциал действия. [6] Эта деполяризация вызвана очень небольшими чистыми внутренними токами ионов кальция через клеточную мембрану, что приводит к возникновению потенциала действия. [7] [8]
Биокидостимуляторы
[ редактировать ]Биокидостимуляторы являются результатом быстро развивающейся области исследований по замене электронного кардиостимулятора . Биописмекер превращает покоящиеся клетки миокарда (например, клетки предсердий) в клетки кардиостимулятора. Это достигается за счет экспрессии в клетках гена, создающего кардиостимуляторный ток. [9]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Берн, Роберт; Мэтью Леви; Брюс Кеппен; Брюс Стэнтон (2004). Физиология . Эльзевир Мосби. п. 276 . ISBN 978-0-8243-0348-8 .
- ^ Веркерк А.О., ван Борен М.М., Петерс Р.Дж., Брукхейс Э., Лам К., Коронель Р., де Баккер Дж.М., Тан Х.Р. (октябрь 2007 г.). «Ток кардиостимулятора (I)f)) в синоатриальном узле человека» . Эур Харт Дж . 28 (1): 2472–8. doi : 10.1093/eurheartj/ehm339 . ПМИД 17823213 .
- ^ Борон, Уолтер. Ф; Эмиль Бульпап (2003). Медицинская физиология . Эльзевир Сондерс. п. 489. ИСБН 978-0-7216-0076-5 .
- ^ ДиФранческо Д. (май 2006 г.). «Забавные каналы в контроле сердечного ритма и механизм действия селективных блокаторов». Фармакол. Рез . 53 (5): 399–406. дои : 10.1016/j.phrs.2006.03.006 . ПМИД 16638640 .
- ^ Моргадо-Валле, Консуэло; Бельтран-Паррасаль, Луис (2017). «Генерация дыхательного ритма: целое больше, чем сумма частей». Пластиковый мозг . Достижения экспериментальной медицины и биологии. Том. 1015. стр. 147–161. дои : 10.1007/978-3-319-62817-2_9 . ISBN 978-3-319-62815-8 . ISSN 0065-2598 . ПМИД 29080026 .
- ^ Кэмпбелл, Нил. А (1996). Биология . Бенджамин Каммингс. п. Г–21. ISBN 978-0-07-366175-9 .
- ^ Веркерк А.О., ван Гиннекен А.С., Вилдерс Р. (январь 2009 г.). «Пейсмекерная активность синоатриального узла человека: роль тока, активируемого гиперполяризацией, I (f)». Инт Джей Кардиол . 132 (3): 318–36. дои : 10.1016/j.ijcard.2008.12.196 . ПМИД 19181406 .
- ^ Борон, Уолтер. Ф; Эмиль Бульпап (2003). Медицинская физиология . Эльзевир Сондерс. п. 487. ИСБН 978-0-7216-0076-5 .
- ^ Веркерк А.О., Зегерс Дж.Г., Ван Гинекен А.С., Вилдерс Р. (2008). «Динамический зажим потенциала действия как мощный инструмент в разработке генного биокардиостимулятора». 2008 30-я ежегодная международная конференция Общества инженерии в медицине и биологии IEEE . Том. 1. С. 133–6. дои : 10.1109/IEMBS.2008.4649108 . ISBN 978-1-4244-1814-5 . ПМИД 19162611 . S2CID 6543917 .
{{cite book}}: CS1 maint: дата и год ( ссылка )