Трихроматия

Трихроматия или трихроматизм наличие трех независимых каналов для передачи цветовой информации, получаемых от трех разных типов колбочек глаза — это . [1] Организмы, обладающие трихроматией, называются трихроматами.
организма Обычное объяснение трихромазии состоит в том, что сетчатка содержит три типа цветовых рецепторов (называемых колбочками у позвоночных ) с разными спектрами поглощения . В действительности число таких типов рецепторов может превышать три, поскольку разные типы могут быть активны при разной интенсивности света. У позвоночных с тремя типами колбочек при низкой интенсивности света палочки могут способствовать цветовому зрению .
Люди и другие животные, являющиеся трихроматами
[ редактировать ]Люди и некоторые другие млекопитающие развили трихроматию , частично основанную на пигментах, унаследованных от ранних позвоночных. Например, у рыб и птиц четыре пигмента для зрения используются . Эти дополнительные зрительные пигменты рецепторов колбочек улавливают энергию других длин волн , иногда включая ультрафиолет . В конце концов два из этих пигментов были потеряны (у плацентарных млекопитающих ), а другой появился, что привело к трихроматии у некоторых приматов . [2] Люди и близкородственные приматы обычно являются трихроматами, как и некоторые самки большинства видов обезьян Нового Света , а также самцы и самки ревунов . [3]
Недавние исследования показывают, что трихроматия может быть широко распространена среди сумчатых . [4] Исследование, проведенное в отношении трихроматии у австралийских сумчатых, предполагает, что чувствительность к средней длине волны (MWS), колбочки медового опоссума ( Tarsipes rostratus ) и толстохвостого даннарта ( Sminthopsis crassicaudata ) являются особенностями, возникающими в результате унаследованного рептилий строения сетчатки . Возможность трихроматии у сумчатых потенциально имеет иную эволюционную основу, чем у приматов . Дальнейшие биологические и поведенческие тесты могут подтвердить, является ли трихроматия общей характеристикой сумчатых. [2]
Большинство других млекопитающих в настоящее время считаются дихроматами , имеющими только два типа колбочек (хотя ограниченная трихроматия возможна при низких уровнях освещенности, когда активны и палочки, и колбочки). [5] Большинство исследований хищных животных, как и других млекопитающих, выявляют дихроматию ; примеры включают домашнюю собаку , хорька и пятнистую гиену . [6] [7] Некоторые виды насекомых (например, медоносные пчелы ) также являются трихроматами, чувствительными к ультрафиолету , синему и зеленому цвету, а не к синему, зеленому и красному. [3]
Исследования показывают, что трихроматия позволяет животным отличать ярко окрашенные фрукты и молодые листья от другой растительности, которая не способствует их выживанию. [8] Другая теория состоит в том, что обнаружение покраснения кожи и, следовательно, настроения могло повлиять на развитие трихроматного зрения у приматов. Красный цвет также оказывает и другие эффекты на поведение приматов и человека, о чем говорится в статье о психологии цвета . [9]
Типы шишек, встречающиеся исключительно у приматов
[ редактировать ]Приматы - единственные известные трихроматы плацентарных млекопитающих. [10] [ не удалось пройти проверку ] Их глаза состоят из трех разных типов колбочек, каждый из которых содержит различный фотопигмент ( опсин ). Их пиковая чувствительность приходится на синюю (коротковолновые колбочки S), зеленую (средневолновые колбочки M) и желто-зеленую (длинноволновые колбочки L) области цветового спектра. [11] Шишки S составляют 5–10% шишек и образуют правильную мозаику. Специальные биполярные и ганглиозные клетки передают эти сигналы от S-конусов, и есть свидетельства того, что они имеют отдельный сигнальный путь через таламус в зрительную кору также . С другой стороны, колбочки L и M трудно отличить по форме или другим анатомическим признакам — их опсины различаются только по 15 из 363 аминокислот, поэтому никому пока не удалось выработать к ним специфические антитела. Но Моллон и Боумейкер [12] обнаружил, что конусы L и конусы M распределены случайным образом и находятся в одинаковом количестве. [13]
Механизм трехцветного цветового зрения
[ редактировать ]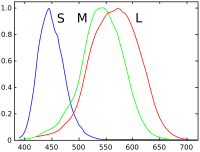

В столбце 1 шар освещен монохроматическим светом. Умножение спектра на кривые спектральной чувствительности колбочек дает ответ для каждого типа колбочек.
В столбце 2 метамерия используется для моделирования сцены с синими, зелеными и красными светодиодами, дающими аналогичный отклик.
Трихроматическое цветовое зрение — это способность людей и некоторых других животных видеть разные цвета , опосредованная взаимодействием между тремя типами цветочувствительных колбочек . Теория трихроматического цвета возникла в 18 веке, когда Томас Янг предположил, что цветовое зрение является результатом деятельности трех разных фоторецепторных клеток . С середины XIX века в своем «Трактате о физиологической оптике » [14] [15] Герман фон Гельмгольц позже расширил идеи Янга, используя эксперименты по сопоставлению цветов, которые показали, что людям с нормальным зрением необходимы три длины волны для создания нормального диапазона цветов. Физиологические доказательства трихроматической теории были позже даны Гуннаром Светичиным (1956). [16]
Каждый из трех типов колбочек сетчатки глаза содержит ретиналем разного типа светочувствительный пигмент , который состоит из трансмембранного белка, называемого опсином , и светочувствительной молекулы, называемой 11-цис- . Каждый отдельный пигмент особенно чувствителен к определенной длине волны ( света то есть пигмент с наибольшей вероятностью вызовет клеточную реакцию, когда на него попадет фотон с определенной длиной волны, к которой этот пигмент наиболее чувствителен). Три типа колбочек — L, M и S — содержат пигменты, которые лучше всего реагируют на свет длинных (особенно 560 нм), средних (530 нм) и коротких (420 нм) волн соответственно. [17] [18]
Поскольку вероятность реакции данного конуса зависит не только от длины волны падающего на него света, но и от его интенсивности , мозг не смог бы различать разные цвета, если бы он получал входные данные только от одного типа колбочек. Таким образом, для возникновения способности воспринимать цвет необходимо взаимодействие как минимум двух типов колбочек. Имея как минимум два типа колбочек, мозг может сравнивать сигналы каждого типа и определять как интенсивность, так и цвет света. Например, умеренная стимуляция средневолновой колбочки может означать, что она стимулируется очень ярким красным (длинноволновым) светом или не очень интенсивным желтовато-зеленым светом. Но очень яркий красный свет будет вызывать более сильную реакцию от колбочек L, чем от колбочек M, тогда как не очень интенсивный желтоватый свет будет вызывать более сильную реакцию от колбочек M, чем от других колбочек. Таким образом, трихроматическое цветовое зрение достигается за счет использования комбинаций клеточных реакций.
Подсчитано, что среднестатистический человек может различать до десяти миллионов различных цветов. [19]
См. также
[ редактировать ]- Визуальная система
- Монохромность
- Дихроматия
- Тетрахромация
- Пентахроматия
- Креветки-богомолы (додекахроматы)
- Эволюция цветового зрения у приматов
- Теория Янга – Гельмгольца
- Цветовое пространство LMS
Ссылки
[ редактировать ]- ^ «Цветовой словарь» . Архивировано из оригинала 4 октября 2015 года . Проверено 8 ноября 2006 г.
- ^ Перейти обратно: а б Аррезе, Кэтрин; Томас, Натан; Бизли, Лин; Шанд, Джулия (2002). «Трихроматия у австралийских сумчатых» . Современная биология . 12 (8): 657–660. Бибкод : 2002CBio...12..657A . дои : 10.1016/S0960-9822(02)00772-8 . ПМИД 11967153 . S2CID 14604695 .
- ^ Перейти обратно: а б Роу, Майкл Х (2002). «Трихроматическое цветовое зрение у приматов» . Новости физиологических наук . 17 (3): 93–98. дои : 10.1152/nips.01376.2001 . ПМИД 12021378 . S2CID 15241669 .
- ^ Арресе, Калифорния; Одди, AY; Ранхэм, ПБ; Харт, Н.С.; Шанд, Дж; Хант, DM (2005). «Топография конуса и спектральная чувствительность у двух потенциально трехцветных сумчатых: квокка ( Setonix brachyurus ) и квенда ( Isooodon obesulus )» . Труды Лондонского королевского общества Б. 272 (1595): 791–796. дои : 10.1098/рспб.2004.3009 . ПМК 1599861 . ПМИД 15888411 .
- ^ Джейкобс, Джеральд Х.; Натанс, Джереми (2009). «Эволюция цветового зрения приматов» . Научный американец . 300 (4): 56–63. Бибкод : 2009SciAm.300d..56J . doi : 10.1038/scientificamerican0409-56 (неактивен 2 апреля 2024 г.). ISSN 0036-8733 . JSTOR 26001303 . ПМИД 19363921 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Кальдероне, Дж.Б.; Джейкобс, GH (2003). «Спектральные свойства и распределение хорьковых колбочек в сетчатке» (PDF) . Визуальная нейронаука . 20 (1): 11–17. дои : 10.1017/s0952523803201024 . ПМИД 12699079 . S2CID 10415194 .
- ^ Кальдероне, Дж.Б.; Риз, Бельгия; Джейкобс, GH (2003). «Топография фоторецепторов и ганглиозных клеток сетчатки пятнистой гиены ( Crocuta crocuta )». Мозг, поведение и эволюция . 62 (4): 182–192. дои : 10.1159/000073270 . ПМИД 14573992 . S2CID 9167855 .
- ^ Шарп Л.Т., де Лука Э., Хансен Т., Ягле Х., Гегенфуртнер К.Р. (2006). «Преимущества и недостатки человеческой дихромазии» . Журнал видения . 6 (3): 213–223. дои : 10.1167/6.3.3 . ПМИД 16643091 .
- ^ Дайана Видерманн, Роберт А. Бартон и Рассел А. Хилл. Эволюционные взгляды на спорт и соревнования. В Робертс, Южная Каролина (2011). Робертс, С. Крейг (ред.). Прикладная эволюционная психология . Издательство Оксфордского университета. doi : 10.1093/acprof:oso/9780199586073.001.0001 . ISBN 9780199586073 .
- ^ Рональд Г. Бут (2002). Восприятие визуальной среды . Спрингер. п. 219. ИСБН 978-0-387-98790-3 .
- ^ Шнапф, Дж.Л.; Крафт, ТВ; Бэйлор, Д.А. (январь 1987 г.). «Спектральная чувствительность фоторецепторов колбочек человека». Природа . 325 (6103): 439–441. Бибкод : 1987Natur.325..439S . дои : 10.1038/325439a0 . ПМИД 3808045 . S2CID 11399054 .
- ^ Моллон, доктор медицинских наук; Боумейкер, Дж. К. (декабрь 1992 г.). «Пространственное расположение колбочек в ямке приматов». Природа . 360 (6405): 677–679. Бибкод : 1992Natur.360..677M . дои : 10.1038/360677a0 . ПМИД 1465131 . S2CID 4234999 .
- ^ Вассле, Хайнц (11 февраля 1999 г.). «Цветовое зрение: лоскутное одеяло из колбочек». Природа . 397 (6719): 473–475. Бибкод : 1999Natur.397..473W . дои : 10.1038/17216 . ПМИД 10028963 . S2CID 4431471 .
- ^ Гельмгольц, Герман (1909). Справочник по физиологической оптике (3-е изд.). Гамбург; Лейпциг: Леопольд Восс . Проверено 18 февраля 2020 г.
- ^ фон Гельмгольц, Герман (2013). Трактат по физиологической оптике . Курьерская корпорация. ISBN 978-0486174709 . Проверено 18 февраля 2020 г. .
- ^ Светичин, Г (1956). «Кривые спектрального отклика одиночных конусов». Acta Physiologica Scandinavica . 39 (134): 17–46. ПМИД 13444020 .
- ^ Свеча ER , Шварц Дж.Х., Джесселл Т.М. (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 100-1 182–185 . ISBN 978-0-8385-7701-1 .
- ^ Джейкобс Г.Х., Натанс Дж. (март 2009 г.). «Цветовое зрение: как наши глаза отражают эволюцию приматов» . Научный американец.
- ^ Леонг, Дженнифер. «Количество цветов, различимых человеческим глазом» . гиперучебник . Проверено 21 февраля 2013 г.
Внешние ссылки
[ редактировать ]- The Straight Dope : «Действительно ли кошки и собаки дальтоники? Откуда они это знают?»
- Светечин, Гуннар; МакНикол, Эдвард Ф. (ноябрь 1958 г.). «Ретинальные механизмы хроматического и ахроматического зрения». Анналы Нью-Йоркской академии наук . 74 (2): 385–404. Бибкод : 1959NYASA..74..385S . дои : 10.1111/j.1749-6632.1958.tb39560.x . ПМИД 13627867 . S2CID 27130943 .