Хемотаксис
Хемотаксис (от хемо- + таксис ) — это движение организма или сущности в ответ на химический раздражитель. [ 1 ] Соматические клетки , бактерии и другие одноклеточные или многоклеточные организмы направляют свои движения в соответствии с определенными химическими веществами в окружающей среде. Это важно для бактерий, чтобы найти пищу (например, глюкозу ), плывя к самой высокой концентрации пищевых молекул, или спасаясь от ядов (например, фенола ). У многоклеточных организмов хемотаксис имеет решающее значение для раннего развития (например, движения спермы к яйцеклетке во время оплодотворения ) и развития (например, миграции нейронов или лимфоцитов ), а также для нормального функционирования и здоровья (например, миграция лейкоцитов во время травмы или инфекция). [ 2 ] Кроме того, было признано, что механизмы, обеспечивающие хемотаксис у животных, могут быть нарушены во время рака метастазирования . [ 3 ] а аберрантное изменение общих свойств этих сетей, которые контролируют хемотаксис, может привести к канцерогенезу . [ 4 ] Аберрантный хемотаксис лейкоцитов и лимфоцитов также способствует воспалительным заболеваниям, таким как атеросклероз, астма и артрит. [ 5 ] [ 6 ] [ 7 ] [ 8 ] Субклеточные компоненты, такие как участок полярности, генерируемый спаривающимися дрожжами, также могут проявлять хемотаксическое поведение. [ 9 ]
Положительный хемотаксис возникает, если движение направлено к более высокой концентрации рассматриваемого химического вещества; отрицательный хемотаксис, если движение происходит в противоположном направлении. Химически вызванный кинезис (произвольно направленный или ненаправленный) можно назвать хемокинезом .
История исследований хемотаксиса
[ редактировать ]еще на заре развития микроскопии Хотя миграция клеток была обнаружена Левенгуком , в лекции по хемотаксису в Калифорнийском технологическом институте утверждается, что «эрудитическое описание хемотаксиса было впервые сделано Т. В. Энгельманном (1881) и В. Ф. Пфеффером (1884) у бактерий, и HS Jennings (1906) у инфузорий . [ 10 ] Лауреат Нобелевской премии И. Мечников также внес вклад в изучение этой области в 1882—1886 гг., исследуя процесс как начальный этап фагоцитоза . [ 11 ] Значение хемотаксиса в биологии и клинической патологии было широко признано в 1930-х годах, и к этому времени были разработаны наиболее фундаментальные определения, лежащие в основе этого явления. [ кем? ] Наиболее важные аспекты контроля качества исследований хемотаксиса были описаны Х. Харрисом в 1950-х годах. [ 12 ] В 1960-х и 1970-х годах революция в современной клеточной биологии и биохимии предоставила ряд новых методов, которые стали доступны для исследования мигрирующих клеток-ответчиков и субклеточных фракций, ответственных за хемотаксическую активность. [ 13 ] Доступность этой технологии привела к открытию C5a, основного хемотаксического фактора, участвующего в остром воспалении. Новаторские работы Дж. Адлера модернизировали капиллярный анализ Пфеффера и явились важным поворотным моментом в понимании всего процесса внутриклеточной передачи сигналов бактерий. [ 14 ] [ 15 ]
Бактериальный хемотаксис - общая характеристика
[ редактировать ]
Некоторые бактерии , такие как E. coli , имеют несколько жгутиков на клетку (обычно 4–10). Они могут вращаться двумя способами:
- Вращение против часовой стрелки объединяет жгутики в один вращающийся пучок, заставляя бактерию плавать по прямой линии; и
- Вращение по часовой стрелке разбивает пучок жгутиков на части, так что каждый жгутик направлен в разном направлении, в результате чего бактерия падает на месте. [ 16 ]
Направления вращения указаны для наблюдателя вне клетки, смотрящего вниз по жгутикам в сторону клетки. [ 17 ]
Поведение
[ редактировать ]Общее движение бактерии является результатом чередования фаз падения и плавания, называемых бегущим движением . [ 18 ] В результате траектория движения бактерии, плавающей в однородной среде, будет представлять собой случайное блуждание с относительно прямым плаванием, прерываемым случайными падениями, которые переориентируют бактерию. [ 19 ] Бактерии, такие как E. coli, не могут выбирать направление своего плавания и не могут плавать по прямой более нескольких секунд из-за вращательной диффузии ; другими словами, бактерии «забывают» направление, в котором они движутся. Постоянно оценивая свой курс и корректируя свое движение в неправильном направлении, бактерии могут направить свое случайное движение в сторону благоприятных мест. [ 20 ]
При наличии химического градиента бактерии будут хемотаксировать или направлять свое общее движение в зависимости от градиента. Если бактерия почувствует, что движется в правильном направлении (к аттрактанту/от репеллента), она будет продолжать плыть по прямой в течение более длительного времени, прежде чем упасть; однако, если он движется в неправильном направлении, он упадет раньше. Бактерии, такие как E. coli, используют временное зондирование, чтобы решить, улучшается ли их ситуация или нет, и таким образом находят место с самой высокой концентрацией аттрактанта, обнаруживая даже небольшие различия в концентрации. [ 21 ]
Это предвзятое случайное блуждание является результатом простого выбора между двумя методами случайного движения; а именно акробатика и прямое плавание. [ 22 ] Спиральная природа отдельной жгутиковой нити имеет решающее значение для возникновения этого движения. Белковая структура, составляющая жгутиковую нить, флагеллин , консервативна среди всех жгутиковых бактерий. [ 23 ] Позвоночные животные, похоже, воспользовались этим фактом, обладая иммунным рецептором ( TLR5 ), предназначенным для распознавания этого консервативного белка. [ 24 ]
Как и во многих случаях в биологии, существуют бактерии, которые не следуют этому правилу. Многие бактерии, такие как вибрионы , моножгутиковые и имеют один жгутик на одном полюсе клетки. Их метод хемотаксиса различен. У других есть единственный жгутик, который находится внутри клеточной стенки. Эти бактерии передвигаются, вращая всю клетку, имеющую форму штопора. [ 25 ] [ нужна страница ]
Преобразование сигнала
[ редактировать ]
Химические градиенты воспринимаются множеством трансмембранных рецепторов , называемых метил-принимающими белками хемотаксиса (MCP), которые различаются по молекулам, которые они обнаруживают. [ 26 ] Известно, что во всем бактериальном царстве кодируются тысячи рецепторов MCP. [ 27 ] Эти рецепторы могут связывать аттрактанты или репелленты прямо или косвенно посредством взаимодействия с белками периплазматического пространства . [ 28 ] Сигналы от этих рецепторов передаются через плазматическую мембрану в цитозоль , где белки Che . активируются [ 29 ] Белки Che изменяют частоту переворота и изменяют рецепторы. [ 29 ]
Регуляция жгутика
[ редактировать ]Белки CheW и CheA связываются с рецептором. Отсутствие активации рецептора приводит к CheA по аутофосфорилированию гистидинкиназы одному высококонсервативному остатку гистидина. [ 30 ] [ нужен лучший источник ] CheA, в свою очередь, переносит фосфорильные группы на консервативные остатки аспартата в регуляторах ответа CheB и CheY; CheA представляет собой гистидинкиназу и не переносит активно фосфорильную группу, скорее, регулятор ответа CheB забирает фосфорильную группу у CheA. [ нужна ссылка ] Этот механизм передачи сигнала называется двухкомпонентной системой и является распространенной формой передачи сигнала у бактерий. [ нужна ссылка ] CheY вызывает переворачивание путем взаимодействия с белком-переключателем жгутика FliM, вызывая изменение вращения жгутика с против часовой стрелки на вращение по часовой стрелке. Изменение состояния вращения одного жгутика может разрушить весь пучок жгутиков и вызвать его переворачивание. [ нужна ссылка ]
Регуляция рецепторов
[ редактировать ]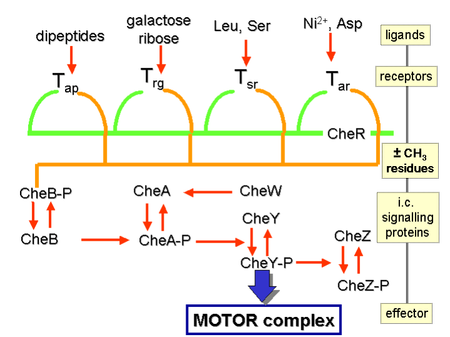
CheB при активации CheA действует как метилэстераза , удаляя метильные группы из остатков глутамата на цитозольной стороне рецептора; он действует антагонистически с CheR, метилтрансферазой , которая добавляет метильные остатки к тем же остаткам глутамата. [ 31 ] Если уровень аттрактанта останется высоким, то уровень фосфорилирования CheA (а, следовательно, CheY и CheB) останется низким, клетка будет плавно плавать, а уровень метилирования MCP увеличится (поскольку CheB-P отсутствует для деметилирования). [ 31 ] MCP больше не реагируют на аттрактант, когда они полностью метилированы; следовательно, даже если уровень аттрактанта может оставаться высоким, уровень CheA-P (и CheB-P) увеличивается, и клетка начинает разрушаться. [ 31 ] MCP могут быть деметилированы CheB-P, и когда это произойдет, рецепторы снова смогут реагировать на аттрактанты. [ 31 ] В отношении репеллентов ситуация противоположная: полностью метилированные МЦП лучше всего реагируют на репелленты, а наименее метилированные МЦП хуже всего реагируют на репелленты. [ нужна ссылка ] Эта регуляция позволяет бактерии «запоминать» химические концентрации из недавнего прошлого, на несколько секунд, и сравнивать их с теми, которые она испытывает в настоящее время, таким образом «знать», движется ли она вверх или вниз по градиенту. [ 32 ] Что бактерии имеют химические градиенты, то в повышении абсолютной величины чувствительности на том или ином фоне участвуют другие механизмы. Хорошо известными примерами являются сверхчувствительная реакция мотора на сигнал CheY-P и кластеризация хеморецепторов. [ 33 ] [ 34 ]
Хемоаттрактанты и хемопелленты
[ редактировать ]Хемоаттрактанты и хеморепелленты — неорганические или органические вещества, обладающие индуктором хемотаксиса в подвижных клетках. Эти хемотаксические лиганды создают градиенты химических концентраций, к которым организмы, прокариотические и эукариотические, движутся навстречу или от них соответственно. [ 35 ]
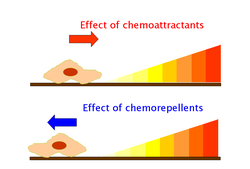
Эффекты хемоаттрактантов вызываются через хеморецепторы, такие как метил-принимающие белки хемотаксиса (MCP). [ 36 ] MCP в E.coli включают Tar, Tsr, Trg и Tap. [ 37 ] Хемоаттрактанты Trg включают рибозу и галактозу с фенолом в качестве хемопеллента. Tap и Tsr распознают дипептиды и серин как хемоаттрактанты соответственно. [ 37 ]
Хемоаттрактанты или хемопелленты связывают MCP во внеклеточном домене; внутриклеточный сигнальный домен передает изменения концентрации этих хемотаксических лигандов нижестоящим белкам, таким как CheA, который затем передает этот сигнал жгутиковым моторам через фосфорилированный CheY (CheY-P). [ 36 ] Затем CheY-P может контролировать вращение жгутиков, влияя на направление подвижности клеток. [ 36 ]
Для E.coli , S. meliloti и R. spheroides связывание хемоаттрактантов с MCP ингибирует активность CheA и, следовательно, CheY-P, что приводит к плавному прохождению, но для B. substilis активность CheA увеличивается. [ 36 ] События метилирования в E.coli приводят к тому, что MCP имеют более низкое сродство к хемоаттрактантам, что вызывает повышение активности CheA и CheY-P, что приводит к падению активности. [ 36 ] Таким образом, клетки способны адаптироваться к немедленной концентрации хемоаттрактанта и обнаруживать дальнейшие изменения для модуляции подвижности клеток. [ 36 ]
Хемоаттрактанты эукариот хорошо характеризуют иммунные клетки. Формильные пептиды , такие как fMLF , привлекают лейкоциты , такие как нейтрофилы и макрофаги , вызывая движение к местам инфекции. [ 38 ] Неацилированные метионинильные пептиды не действуют как хемоаттрактанты на нейтрофилы и макрофаги. [ 38 ] Лейкоциты также движутся к хемоаттрактантам C5a, компоненту комплемента и патоген -специфичным лигандам на бактериях. [ 38 ]
Механизмы действия хемопеллентов менее известны, чем механизмы хемоаттрактантов. Хотя хемопелленты вызывают у организмов реакцию избегания, Tetrahymena thermophila адаптируется к хеморепелленту, пептиду Нетрин-1 , в течение 10 минут после воздействия; однако воздействие хемопеллентов, таких как GTP , PACAP-38 и ноцицептин , не приводит к такой адаптации. [ 39 ] ГТФ и АТФ являются хеморепеллентами в микромолярных концентрациях как для Tetrahymena , так и для Paramecium . Эти организмы избегают этих молекул, вызывая реакции избегания, чтобы переориентироваться от градиента. [ 40 ]
Эукариотический хемотаксис
[ редактировать ]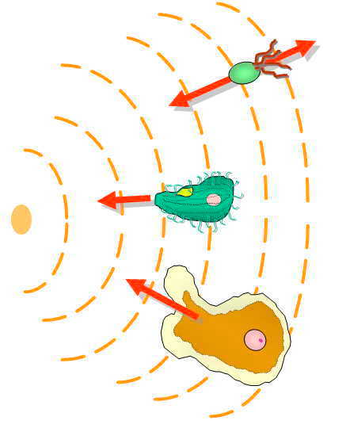
Механизм хемотаксиса эукариотических клеток совершенно отличается от механизма хемотаксиса бактерий E. coli ; однако обнаружение химических градиентов по-прежнему остается важным шагом в этом процессе. [ 41 ] [ нужен лучший источник ] Из-за своего небольшого размера и других биофизических ограничений E. coli не может напрямую обнаруживать градиент концентрации. [ 42 ] Вместо этого они используют измерение временного градиента, при котором они перемещаются на большие расстояния, в несколько раз превышающие их собственную ширину, и измеряют скорость изменения воспринимаемой химической концентрации. [ 43 ] [ 44 ]
Эукариотические клетки намного крупнее прокариот и имеют рецепторы, равномерно расположенные по всей клеточной мембране . [ 43 ] Эукариотический хемотаксис включает в себя обнаружение пространственного градиента концентрации путем сравнения асимметричной активации этих рецепторов на разных концах клетки. [ 43 ] Активация этих рецепторов приводит к миграции к хемоаттрактантам или от хеморепеллентов. [ 43 ] У спаривающихся дрожжей, которые неподвижны, участки полярных белков на клеточной коре могут перемещаться хемотаксическим образом вверх по градиентам феромонов. [ 45 ] [ 9 ]
Также было показано, что как прокариотические, так и эукариотические клетки способны к хемотаксической памяти. [ 44 ] [ 46 ] У прокариот этот механизм включает метилирование рецепторов, называемых метил-принимающими белками хемотаксиса (MCP). [ 44 ] Это приводит к их десенсибилизации и позволяет прокариотам «помнить» и адаптироваться к химическому градиенту. [ 44 ] Напротив, хемотаксическая память у эукариот может быть объяснена моделью локального возбуждения и глобального торможения (LEGI). [ 46 ] [ 47 ] LEGI включает баланс между быстрым возбуждением и отсроченным торможением, который контролирует нижестоящую передачу сигналов, такую как активация Ras и выработка PIP3 . [ 48 ]
Уровни рецепторов, внутриклеточные сигнальные пути и эффекторные механизмы представляют собой разнообразные компоненты эукариотического типа. В эукариотических одноклеточных клетках основными эффекторами являются амебоидное движение и ресничка или эукариотический жгутик (например, Amoeba или Tetrahymena ). [ 49 ] [ 50 ] Некоторые эукариотические клетки высшего происхождения позвоночных , например иммунные клетки, также перемещаются туда, где им необходимо быть. Помимо иммунокомпетентных клеток ( гранулоцитов , моноцитов , лимфоцитов ), большая группа клеток, ранее считавшихся фиксированными в тканях, также подвижна в особых физиологических (например, тучные клетки , фибробласты , эндотелиальные клетки ) или патологических состояниях (например, метастазы ). . [ 51 ] Хемотаксис имеет большое значение на ранних стадиях эмбриогенеза , поскольку развитие зародышевых листков направляется градиентами сигнальных молекул. [ 52 ] [ 53 ]
Подвижность
[ редактировать ]В отличие от подвижности при бактериальном хемотаксисе, механизм физического перемещения эукариотических клеток неясен. По-видимому, существуют механизмы, с помощью которых внешний хемотаксический градиент воспринимается и превращается во внутриклеточные градиенты Ras и PIP3 к градиенту и активации сигнального пути, кульминацией которого является полимеризация актиновых , что приводит филаментов. Растущий дистальный конец актиновых филаментов развивает связи с внутренней поверхностью плазматической мембраны через различные наборы пептидов и приводит к образованию передних псевдоподий и задних уроподий . [ 54 ] [ 55 ] Реснички эукариотических клеток также могут вызывать хемотаксис; в данном случае это в основном Ca 2+ -зависимая индукция системы микротрубочек базального тельца и ритма 9+2 микротрубочек внутри ресничек. Организованное биение сотен ресничек синхронизируется подмембранной системой, построенной между базальными тельцами. Детали сигнальных путей до сих пор не совсем ясны.
Миграционные реакции, связанные с хемотаксисом
[ редактировать ]
Хемотаксис относится к направленной миграции клеток в ответ на химические градиенты; Существует несколько вариантов химической миграции, перечисленных ниже.
- Хемокинез относится к увеличению подвижности клеток в ответ на химические вещества в окружающей среде. В отличие от хемотаксиса, миграция, стимулируемая хемокинезом, лишена направленности и вместо этого усиливает поведение сканирования окружающей среды. [ 56 ]
- При гаптотаксисе градиент . хемоаттрактанта выражен или связан на поверхности, в отличие от классической модели хемотаксиса, в которой градиент развивается в растворимой жидкости [ 57 ] Наиболее распространенной биологически активной гаптотактической поверхностью является внеклеточный матрикс (ECM); наличие связанных лигандов отвечает за индукцию трансэндотелиальной миграции и ангиогенеза .
- Некротаксис представляет собой особый тип хемотаксиса, когда молекулы хемоаттрактанта высвобождаются из некротических или апоптотических клеток. В зависимости от химического характера выделяемых веществ некротаксис может накапливать или отталкивать клетки, что подчеркивает патофизиологическую значимость этого явления.
Рецепторы
[ редактировать ]В общем, эукариотические клетки ощущают присутствие хемотаксических стимулов посредством использования 7-трансмембранных (или серпентиновых) гетеротримерных рецепторов, связанных с G-белком , - класса, представляющего значительную часть генома . [ 58 ] Некоторые члены этого суперсемейства генов используются в зрении ( родопсины ), а также в обонянии (обонянии). [ 59 ] [ 60 ] Основные классы рецепторов хемотаксиса запускаются:
- Формильные пептиды – формилпептидные рецепторы (FPR),
- Хемокины - хемокиновые рецепторы (CCR или CXCR) и
- Лейкотриены – лейкотриеновые рецепторы (БЛТ). [ 61 ]
Однако индукция широкого набора мембранных рецепторов (например, циклических нуклеотидов , аминокислот , инсулина , вазоактивных пептидов) также вызывает миграцию клетки. [ 62 ]
Хемотаксический отбор
[ редактировать ]
В то время как некоторые рецепторы хемотаксиса экспрессируются на поверхности мембраны с долговременными характеристиками, поскольку они детерминированы генетически, другие имеют краткосрочную динамику, поскольку они собираются ad hoc в присутствии лиганда. [ 63 ] Разнообразные особенности рецепторов и лигандов хемотаксиса позволяют отбирать клетки-ответчики на хемотаксис с помощью простого анализа хемотаксиса. С помощью хемотаксического отбора мы можем определить, действует ли еще не охарактеризованная молекула через долгосрочный или кратковременный рецепторный путь. [ 64 ] Термин «хемотаксический отбор» также используется для обозначения метода, который разделяет эукариотические или прокариотические клетки в соответствии с их хемотаксической чувствительностью к селекторным лигандам. [ 65 ] [ нужен неосновной источник ] [ нужен неосновной источник ]
Хемотаксические лиганды
[ редактировать ]

Число молекул, способных вызывать хемотаксические ответы, относительно велико, и можно выделить первичные и вторичные хемотаксические молекулы. [ нужна ссылка ] Основные группы первичных лигандов следующие:
- Формильные пептиды представляют собой ди-, три-, тетрапептиды бактериального происхождения, формилированные на N-конце пептида. [ нужна ссылка ] [ 66 ] Они выделяются из бактерий in vivo или после разложения клетки. Типичным представителем этой группы является N-формилметиониллейцилфенилаланин (сокращенно fMLF или fMLP). [ нужна ссылка ] Бактериальный fMLF является ключевым компонентом воспаления и оказывает характерное хемоаттрактантное действие на нейтрофильные гранулоциты и моноциты. [ нужна ссылка ] Лиганды и рецепторы хемотаксических факторов, связанные с формильными пептидами, обобщены в соответствующей статье « Рецепторы формилпептидов» .
- Комплемент 3а ( С3а ) и комплемент 5а ( С5а ) являются промежуточными продуктами каскада комплемента. [ нужна ссылка ] Их синтез сопряжен с тремя альтернативными путями (классическим, лектинзависимым и альтернативным) активации комплемента ферментом конвертазой. [ нужна ссылка ] Основными клетками-мишенями этих производных являются также нейтрофильные гранулоциты и моноциты. [ нужна ссылка ]
- Хемокины принадлежат к особому классу цитокинов ; их группы (хемокины C, CC, CXC, CX 3 C) не только представляют собой структурно родственные молекулы с особым расположением дисульфидных мостиков, но и их специфичность к клеткам-мишеням разнообразна. [ нужна ссылка ] Хемокины CC действуют на моноциты (например, RANTES ), а хемокины CXC специфичны для нейтрофильных гранулоцитов (например, IL-8 ). [ нужна ссылка ] Исследования трехмерной структуры хемокинов показали, что характерный состав бета-листов и альфа-спирали обеспечивает экспрессию последовательностей, необходимых для взаимодействия с хемокиновыми рецепторами. [ нужна ссылка ] Образование димеров и их повышенная биологическая активность были продемонстрированы кристаллографией нескольких хемокинов, например IL-8. [ нужна ссылка ]
- Метаболиты полиненасыщенных жирных кислот
- Лейкотриены представляют собой эйкозаноидные липидные медиаторы, образующиеся в результате метаболизма арахидоновой кислоты с помощью ALOX5 (также называемого 5-липоксигеназой). Их наиболее известным членом с активностью хемотаксического фактора является лейкотриен B4 , который вызывает адгезию, хемотаксис и агрегацию лейкоцитов. Хемоаттрактантное действие LTB4 индуцируется посредством любого из двух рецепторов, связанных с G-белком , BLT1 и BLT2 , которые высоко экспрессируются в клетках, участвующих в воспалении и аллергии . [ 67 ]
- Семейство эйкозаноидов 5-гидроксикозатетраеновой кислоты представляет собой метаболиты арахидоновой кислоты, также образуемые ALOX5. Три члена семейства образуются естественным путем и обладают заметной хемотаксической активностью. Они перечислены в порядке убывания эффективности: 5-оксо-эйкозатетраеновая кислота , 5-оксо-15-гидрокси-эйкозатетраеновая кислота и 5-гидроксиэйкозатетраеновая кислота . Это семейство агонистов стимулирует хемотаксические реакции в эозинофилах , нейтрофилах и моноцитах человека путем связывания с оксоэйкозаноидным рецептором 1 , который, как и рецепторы лейкотриена B4, является рецептором, связанным с G-белком. [ 67 ] Помимо кожи, нейтрофилы являются первой линией защиты организма от бактериальных инфекций. Покинув близлежащие кровеносные сосуды, эти клетки распознают химические вещества, вырабатываемые бактериями в порезе или царапине, и мигрируют «в сторону запаха».
- 5-гидроксиэйкозатриеновая кислота и 5-оксоэйкозатриеновая кислота являются метаболитами Мидовой кислоты (5 Z ,8 Z ,11 Z -эйкозатриреноидная кислота); они стимулируют хемотаксис лейкоцитов через оксоэйкозаноидный рецептор 1 [ 68 ] при этом 5-оксоэйкозатриеновая кислота столь же эффективна, как и ее аналог, производный арахидоновой кислоты, 5-оксо-эйкозатетраеновая кислота, в стимуляции хемотаксиса эозинофилов и нейтрофилов крови человека . [ 67 ]
- 12-Гидроксиейкозатетраеновая кислота представляет собой эйкозаноидный метаболит арахидоновой кислоты, образуемый ALOX12 , который стимулирует хемотаксис лейкоцитов через лейкотриеновый рецептор B4, BLT2. [ 67 ]
- Простагландин D2 представляет собой эйкозаноидный метаболит арахидононовой кислоты, вырабатываемый циклооксигеназой 1 или циклооксигеназой 2 , который стимулирует хемотаксис через рецептор простагландина DP2 . Он вызывает хемотаксические реакции в эозинофилах, базофилах и Т-хелперных клетках подтипа Th2. [ 69 ] [ нужен неосновной источник ] [ нужен неосновной источник ]
- 12-Гидроксигептадекатриеновая кислота представляет собой неэйкозаноидный метаболит арахидоновой кислоты, образуемый циклооксигеназой 1 или циклооксигеназой 2, который стимулирует хемотаксис лейкоцитов через лейкотриеновый B4-рецептор, BLT2. [ 70 ] [ нужен неосновной источник ] [ нужен неосновной источник ]
- 15-оксо-эйкозатетраеновая кислота представляет собой эйкозаноидный метаболит арахидоновой кислоты, полученный моим ALOX15 ; он обладает слабой хемотаксической активностью в отношении моноцитов человека (см. 15-Гидроксикозатетраеновая кислота#15-оксо-ЭТЕ ). [ 71 ] [ нужен неосновной источник ] [ нужен неосновной источник ] Рецептор или другой механизм, с помощью которого этот метаболит стимулирует хемотаксис, не выяснен.
Хемотаксический диапазон фитинга
[ редактировать ]
Хемотаксические реакции, вызываемые взаимодействиями лиганд - рецептор, варьируются в зависимости от концентрации лиганда. Исследования семейств лигандов (например, аминокислот или олигопептидов ) показывают, что хемоаттрактантная активность проявляется в широком диапазоне, тогда как хемопеллентная активность имеет узкие диапазоны. [ 72 ]
Клиническое значение
[ редактировать ]Измененный миграционный потенциал клеток имеет сравнительно большое значение в развитии ряда клинических симптомов и синдромов. Изменение хемотаксической активности внеклеточных (например, Escherichia coli ) или внутриклеточных (например, Listeria monocytogenes ) патогенов само по себе представляет собой важную клиническую мишень. Модификация эндогенной хемотаксической способности этих микроорганизмов фармацевтическими агентами может снизить или ингибировать частоту инфекций или распространение инфекционных заболеваний. Помимо инфекций, существуют и другие заболевания, при которых нарушение хемотаксиса является основным этиологическим фактором, например синдром Чедиака-Хигаси , при котором гигантские внутриклеточные везикулы ингибируют нормальную миграцию клеток.
| Тип заболевания | Хемотаксис увеличен | Хемотаксис снизился |
|---|---|---|
| Инфекции | Воспаления | СПИД , Бруцеллез |
| Хемотаксис приводит к заболеванию | — | Синдром Чедиака-Хигаси , синдром Картагенера |
| Нарушается хемотаксис | Атеросклероз , артрит , пародонтит , псориаз , реперфузионное повреждение , метастатические опухоли | Рассеянный склероз , болезнь Ходжкина , мужское бесплодие |
| Отравление | Асбест , бензпирен | Hg и Cr Соли , озон |
Математические модели
[ редактировать ]Разработано несколько математических моделей хемотаксиса в зависимости от типа
- Миграция (например, основные различия бактериального плавания, перемещение одноклеточных эукариот с ресничками / жгутиками и амебоидная миграция)
- Физико-химические характеристики химических веществ (например, диффузия ), действующих как лиганды.
- Биологические характеристики лигандов (аттрактантные, нейтральные и репеллентные молекулы)
- Системы анализа, применяемые для оценки хемотаксиса (см. время инкубации, развитие и стабильность градиентов концентрации)
- Другие воздействия окружающей среды, оказывающие прямое или косвенное влияние на миграцию (освещение, температура, магнитные поля и т.п.)
Хотя взаимодействие перечисленных выше факторов делает поведение решений математических моделей хемотаксиса достаточно сложным, основной феномен движения, обусловленного хемотаксисом, можно описать прямолинейно. Действительно, обозначим через пространственно неоднородная концентрация хемоаттрактанта и как его градиент. Затем хемотаксический клеточный поток (также называемый током) генерируемый хемотаксисом, связан с вышеуказанным градиентом по закону: [ 73 ]




где - пространственная плотность клеток и это так называемый «коэффициент хемотаксиса» - часто является не постоянной, а убывающей функцией хемоаттрактанта. Для некоторого количества который подвержен полному потоку и срок генерации/уничтожения , можно сформулировать уравнение неразрывности :





где это расхождение . Это общее уравнение применимо как к плотности клеток, так и к хемоаттрактанту. Следовательно, если включить диффузионный поток в общий поток, взаимодействия между этими величинами будут определяться набором связанных производных реакция-диффузия, дифференциальных уравнений в частных описывающих изменение и : [ 73 ]

![{\displaystyle {\begin{aligned}{\partial C \over {\partial t}}&=f(C)+\nabla \cdot \left[D_{C}\nabla CC\chi (\varphi)\nabla \color \right]\\{\partial \color \over {\partial t}}&=g(\color ,C)+\color \cdot (D_ {\color }\color \color )\end{aligned} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/924b3d46d7dc2a27228a7d98cf43d035a4622c41)
где описывает рост плотности клеток, представляет собой термин кинетики/источника для хемоаттрактанта, а коэффициенты диффузии для плотности клеток и хемоаттрактанта соответственно равны и .




Пространственная экология почвенных микроорганизмов является функцией их хемотаксической чувствительности к субстрату и другим организмам. [ 74 ] [ нужен неосновной источник ] [ нужен неосновной источник ] Было доказано, что хемотаксическое поведение бактерий приводит к нетривиальным структурам популяций даже в отсутствие гетерогенности окружающей среды. Наличие структурных неоднородностей порового масштаба оказывает дополнительное влияние на возникающие бактериальные структуры.
Измерение хемотаксиса
[ редактировать ]Доступен широкий спектр методов для оценки хемотаксической активности клеток или хемоаттрактантного и хеморепеллентного характера лигандов. Основные требования к измерениям следующие:
- Градиенты концентрации могут развиваться относительно быстро и длительно сохраняться в системе.
- Различают хемотаксическую и хемокинетическую активность.
- Миграция клеток свободна вперед и назад по оси градиента концентрации.
- Обнаруженные ответы являются результатом активной миграции клеток.
Несмотря на то, что идеальный анализ хемотаксиса до сих пор не доступен, существует несколько протоколов и оборудования, которые хорошо соответствуют условиям, описанным выше. Наиболее часто используемые приведены в таблице ниже:
| Тип анализа | Анализы на агаровых пластинках | Двухкамерные анализы | Другие |
|---|---|---|---|
| Примеры |
|
|
|
Искусственные хемотаксические системы
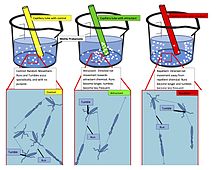
[ редактировать ]химические роботы , которые используют искусственный хемотаксис для автономной навигации. Были разработаны [ 75 ] [ 76 ] Приложения включают адресную доставку лекарств в организм. [ 77 ] Совсем недавно молекулы ферментов также продемонстрировали положительное хемотаксическое поведение в градиенте своих субстратов. [ 78 ] Термодинамически выгодное связывание ферментов со специфическими субстратами считается причиной ферментативного хемотаксиса. [ 79 ] Кроме того, ферменты в каскадах также демонстрируют хемотаксическую агрегацию, управляемую субстратом. [ 80 ]
Помимо активных ферментов, нереагирующие молекулы также проявляют хемотаксическое поведение. Это было продемонстрировано с помощью молекул красителя, которые движутся направленно в градиентах раствора полимера за счет благоприятных гидрофобных взаимодействий. [ 81 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Чисхолм Х , изд. (1911). . Британская энциклопедия . Том. 6 (11-е изд.). Издательство Кембриджского университета. п. 77.
- ^ де Оливейра С., Розовский Э.Э., Хуттенлохер А. (май 2016 г.). «Миграция нейтрофилов при инфекции и заживлении ран: движение вперед в обратном направлении» . Обзоры природы. Иммунология . 16 (6): 378–91. дои : 10.1038/nri.2016.49 . ПМК 5367630 . ПМИД 27231052 .
- ^ Стелтен CH, Parent CA, Montell DJ (май 2018 г.). «Подвижность клеток при инвазии и метастазировании рака: данные простых модельных организмов» . Обзоры природы. Рак . 18 (5): 296–312. дои : 10.1038/nrc.2018.15 . ПМК 6790333 . ПМИД 29546880 .
- ^ Чжан Х., Бхаттачарья С., Цай Х., Иглесиас П.А., Хуан Ч., Девреотес П.Н. (сентябрь 2020 г.). «Возбудимая сигнальная сеть Ras/PI3K/ERK контролирует миграцию и онкогенную трансформацию в эпителиальных клетках» . Развивающая клетка . 54 (5): 608–623. дои : 10.1016/j.devcel.2020.08.001 . ПМК 7505206 . ПМИД 32877650 .
- ^ Ли Дж., Лей К. (январь 2015 г.). «Миграция лимфоцитов в атеросклеротическую бляшку» . Атеросклероз, тромбоз и сосудистая биология . 35 (1): 40–9. дои : 10.1161/ATVBAHA.114.303227 . ПМЦ 4429868 . ПМИД 25301842 .
- ^ Гельфанд РЭБ (октябрь 2017). «Важность путей лейкотриенов B4-BLT1 и LTB4-BLT2 при астме» . Семинары по иммунологии . 33 : 44–51. дои : 10.1016/j.smim.2017.08.005 . ПМЦ 5679233 . ПМИД 29042028 .
- ^ Планагума А., Доменек Т., Понт М., Калама Э., Гарсиа-Гонсалес В., Лопес Р. и др. (октябрь 2015 г.). «Комбинированная терапия анти-CXC-рецепторами 1 и 2 является многообещающим противовоспалительным лечением респираторных заболеваний за счет уменьшения миграции и активации нейтрофилов». Легочная фармакология и терапия . 34 : 37–45. дои : 10.1016/j.pupt.2015.08.002 . ПМИД 26271598 .
- ^ Рана АК, Ли Й, Данг К, Ян Ф (декабрь 2018 г.). «Моноциты при ревматоидном артрите: циркулирующие предшественники макрофагов и остеокластов и их роль гетерогенности и пластичности в патогенезе РА». Международная иммунофармакология . 65 : 348–359. дои : 10.1016/j.intimp.2018.10.016 . ПМИД 30366278 . S2CID 53116963 .
- ^ Jump up to: а б Гоуз Д., Джейкобс К., Рамирес С., Элстон Т., Лью Д. (июнь 2021 г.). «Хемотаксическое перемещение участка полярности позволяет дрожжевым клеткам находить себе пару» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (22): e2025445118. Бибкод : 2021PNAS..11825445G . дои : 10.1073/pnas.2025445118 . ПМК 8179161 . ПМИД 34050026 .
- ^ Филлипс Р. (2007). «Как клетки решают, куда идти: случай бактериального хемотаксиса» (PDF) . Лекция по хемотаксису . Архивировано из оригинала (PDF) 19 июня 2010 года . Проверено 15 апреля 2017 г.
- ^ «Эли Мечников» . энциклопедия Британская Британская энциклопедия, Inc. 12 мая 2024 г.
- ^ Робертс Б., Чунг Э., Ю Ш., Ли С.З. и др. (Математический метод биоинженерной группы, презентация) (2012). «Модели Келлера-Сегеля для хемотаксиса» (PDF) . Интегрированные системы нейроинженерии . Калифорнийский университет – Сан-Диего. Архивировано из оригинала (PDF) 29 августа 2017 года . Проверено 1 апреля 2017 г.
- ^ Снайдерман Р., Гевурц Х., Мергенхаген С.Е. (август 1968 г.). «Взаимодействие системы комплемента с эндотоксичными липополисахаридами. Генерация фактора хемотаксиса полиморфноядерных лейкоцитов» . Журнал экспериментальной медицины . 128 (2): 259–75. дои : 10.1084/jem.128.2.259 . ПМК 2138524 . ПМИД 4873021 .
- ^ Адлер Дж., Цо В.В. (июнь 1974 г.). « Принятие решений у бактерий: хемотаксический ответ Escherichia coli на противоречивые стимулы». Наука . 184 (4143): 1292–4. Бибкод : 1974Sci...184.1292A . дои : 10.1126/science.184.4143.1292 . ПМИД 4598187 . S2CID 7221477 .
- ^ Берг Х (2004). Берг Х.К. (ред.). Кишечная палочка в движении . Биологическая и медицинская физика, Биомедицинская инженерия. Спрингер. п. 15 , 19–29. дои : 10.1007/b97370 . ISBN 0-387-00888-8 . S2CID 35733036 .
- ^ Юань Дж., Фарнер К.А., Тернер Л., Берг Х.К. (июль 2010 г.). «Асимметрия вращения бактериального жгутикового мотора по и против часовой стрелки» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (29): 12846–9. Бибкод : 2010PNAS..10712846Y . дои : 10.1073/pnas.1007333107 . ПМЦ 2919929 . ПМИД 20615986 .
- ^ «Бактериальный хемотаксис» (PDF) . Архивировано (PDF) из оригинала 6 мая 2017 года.
- ^ Берг Х.К., Браун Д.А. (октябрь 1972 г.). «Хемотаксис Escherichia coli, анализируемый с помощью трехмерного отслеживания». Природа . 239 (5374): 500–504. Бибкод : 1972Natur.239..500B . дои : 10.1038/239500a0 . ПМИД 4563019 . S2CID 1909173 .
- ^ Сурджик В., Вингрин Н.С. (апрель 2012 г.). «Реакция на химические градиенты: бактериальный хемотаксис» . Современное мнение в области клеточной биологии . 24 (2): 262–268. дои : 10.1016/j.ceb.2011.11.008 . ПМК 3320702 . ПМИД 22169400 .
- ^ Берг ХК (1993). Случайные блуждания в биологии (Расширенная, переработанная ред.). Принстон, Нью-Джерси: Princeton Univ. Нажимать. стр. 83–94. ISBN 978-0-691-00064-0 .
- ^ Сурджик В., Вингрин Н. (апрель 2012 г.). «Реакция на химические градиенты: бактериальный хемотаксис» . Современное мнение в области клеточной биологии . 24 (2): 262–8. дои : 10.1016/j.ceb.2011.11.008 . ПМК 3320702 . ПМИД 22169400 .
- ^ Макнаб Р.М., Кошланд, Д.Э. (сентябрь 1972 г.). «Механизм восприятия градиента в бактериальном хемотаксисе» . Труды Национальной академии наук Соединенных Штатов Америки . 69 (9): 2509–2512. Бибкод : 1972PNAS...69.2509M . дои : 10.1073/pnas.69.9.2509 . ПМК 426976 . ПМИД 4560688 .
- ^ Неделькович М., Састре Д.Е., Сундберг Э.Ю. (июль 2021 г.). «Бактериальная жгутиковая нить: супрамолекулярная многофункциональная наноструктура» . Международный журнал молекулярных наук . 22 (14): 7521. doi : 10.3390/ijms22147521 . ПМК 8306008 . ПМИД 34299141 .
- ^ Чжун М., Ян Х., Ли Ю (октябрь 2017 г.). «Флагеллин: уникальная молекулярная структура, связанная с микробами, и многогранный иммуномодулятор» . Клеточная и молекулярная иммунология . 14 (10): 862–864. дои : 10.1038/cmi.2017.78 . ПМК 5649114 . ПМИД 28845044 .
- ^ Берг ХК (2003). Кишечная палочка в движении . Нью-Йорк, штат Нью-Йорк: Спрингер. ISBN 978-0-387-00888-2 . [ нужна страница ]
- ^ Уодхамс Г.Х., Армитидж Дж.П. (декабрь 2004 г.). «Понимание всего этого: бактериальный хемотаксис». Обзоры природы. Молекулярно-клеточная биология . 5 (12): 1024–1037. дои : 10.1038/nrm1524 . ПМИД 15573139 . S2CID 205493118 .
- ^ Гальперин М.Ю. (июнь 2005 г.). «Перепись мембраносвязанных и внутриклеточных белков сигнальной трансдукции у бактерий: бактериальный IQ, экстраверты и интроверты» . БМК Микробиология . 5:35 . дои : 10.1186/1471-2180-5-35 . ПМЦ 1183210 . ПМИД 15955239 .
- ^ Аулетта Дж (2011). Когнитивная биология: работа с информацией от бактерий до разума . США: Издательство Оксфордского университета. п. 266. ИСБН 978-0-19-960848-5 .
- ^ Jump up to: а б Фальке Дж.Дж., Басс Р.Б., Батлер С.Л., Червиц С.А., Дэниэлсон М.А. (1997). «Двухкомпонентный сигнальный путь бактериального хемотаксиса: молекулярный взгляд на передачу сигнала рецепторами, киназами и адаптационными ферментами» . Ежегодный обзор клеточной биологии и биологии развития . 13 : 457–512. дои : 10.1146/annurev.cellbio.13.1.457 . ПМК 2899694 . ПМИД 9442881 .
- ^ ToxCafe (2 июня 2011 г.). «Хемотаксис» . Архивировано из оригинала 11 июля 2015 года . Проверено 23 марта 2017 г. - через YouTube.
- ^ Jump up to: а б с д Уодхамс Г.Х., Армитидж Дж.П. (декабрь 2004 г.). «Понимание всего этого: бактериальный хемотаксис». Обзоры природы. Молекулярно-клеточная биология . 5 (12): 1024–1037. дои : 10.1038/nrm1524 . ПМИД 15573139 . S2CID 205493118 .
- ^ Шу С., Чен ПК, Фунг Ю.К. (2008). Вводный текст в биоинженерию (Продвинутая серия по биомеханике - Том 4) . Сингапур: World Scientific Publishing Co. Pte. ООО с. 418. ИСБН 9789812707932 .
- ^ Клюзель П., Сюретт М., Лейблер С. (март 2000 г.). «Сверхчувствительный бактериальный мотор, выявленный путем мониторинга сигнальных белков в отдельных клетках». Наука . 287 (5458): 1652–5. Бибкод : 2000Sci...287.1652C . дои : 10.1126/science.287.5458.1652 . ПМИД 10698740 . S2CID 5334523 .
- ^ Сурджик В. (декабрь 2004 г.). «Кластеризация рецепторов и обработка сигналов в хемотаксисе E. coli». Тенденции в микробиологии . 12 (12): 569–76. CiteSeerX 10.1.1.318.4824 . дои : 10.1016/j.tim.2004.10.003 . ПМИД 15539117 .
- ^ Сюй Ф, Бирман Р, Хили Ф, Нгуен Х (2016). «Многомасштабная модель хемотаксиса Escherichia coli от внутриклеточного сигнального пути до подвижности и поглощения питательных веществ в градиенте питательных веществ и изотропной жидкой среде» . Компьютеры и математика с приложениями . 71 (11): 2466–2478. дои : 10.1016/j.camwa.2015.12.019 .
- ^ Jump up to: а б с д и ж Сзурмант Х., Ордал Г.В. (июнь 2004 г.). «Разнообразие механизмов хемотаксиса бактерий и архей» . Обзоры микробиологии и молекулярной биологии . 68 (2): 301–19. дои : 10.1128/MMBR.68.2.301-319.2004 . ПМК 419924 . ПМИД 15187186 .
- ^ Jump up to: а б Ямамото К., Макнаб Р.М., Имаэ Ю. (январь 1990 г.). «Функции репеллентного ответа хеморецепторов Trg и Tap Escherichia coli» . Журнал бактериологии . 172 (1): 383–8. дои : 10.1128/jb.172.1.383-388.1990 . ПМК 208443 . ПМИД 2403544 .
- ^ Jump up to: а б с Шиффманн Э., Коркоран Б.А., Валь С.М. (март 1975 г.). «N-формилметионилпептиды как хемоаттрактанты лейкоцитов» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (3): 1059–62. Бибкод : 1975PNAS...72.1059S . дои : 10.1073/pnas.72.3.1059 . ПМК 432465 . ПМИД 1093163 .
- ^ Курувилла Х., Шмидт Б., Сонг С., Бхаджан М., Мерикал М., Элли С. и др. (2016). «Пептид нетрин-1 является хеморепеллентом у Tetrahymena thermophila» . Международный журнал пептидов . 2016 : 7142868. doi : 10.1155/2016/7142868 . ПМЦ 4830718 . ПМИД 27123011 .
- ^ Хеннесси ТМ (июнь 2005 г.). «Реакция инфузорий Tetrahymena и Paramecium на внешние АТФ и ГТФ» . Пуринергическая сигнализация . 1 (2): 101–10. дои : 10.1007/s11302-005-6213-1 . ПМК 2096533 . ПМИД 18404496 .
- ^ Кохидай Л (2016). «Хемотаксис как выражение общения тетрахимен». В Вицани Г., Новацки М. (ред.). Биокоммуникация инфузорий . стр. 65–82. дои : 10.1007/978-3-319-32211-7_5 . ISBN 978-3-319-32211-7 .
- ^ Берг Х.К., Перселл Э.М. (ноябрь 1977 г.). «Физика хеморецепции» . Биофизический журнал . 20 (2): 193–219. Бибкод : 1977BpJ....20..193B . дои : 10.1016/s0006-3495(77)85544-6 . ПМЦ 1473391 . ПМИД 911982 .
- ^ Jump up to: а б с д Левин Х., Раппель В.Дж. (февраль 2013 г.). «Физика хемотаксиса эукариот» . Физика сегодня . 66 (2): 24–30. Бибкод : 2013ФТ....66б..24Л . дои : 10.1063/PT.3.1884 . ПМЦ 3867297 . ПМИД 24363460 .
- ^ Jump up to: а б с д Владимиров Н., Сурджик В. (ноябрь 2009 г.). «Хемотаксис: как бактерии используют память». Биологическая химия . 390 (11): 1097–1104. дои : 10.1515/BC.2009.130 . ПМИД 19747082 . S2CID 207440927 .
- ^ Гоуз Д., Лью Д. (май 2020 г.). «Механистический взгляд на движение участков полярности, управляемое актином, у дрожжей» . Молекулярная биология клетки . 31 (10): 1085–1102. дои : 10.1091/mbc.e20-01-0040 . ПМЦ 7346724 . ПМИД 32186970 .
- ^ Jump up to: а б Скоге М., Юэ Х., Эрикстад М., Бэ А., Левин Х., Гройсман А. и др. (октябрь 2014 г.). «Клеточная память в хемотаксисе эукариот» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (40): 14448–53. Бибкод : 2014PNAS..11114448S . дои : 10.1073/pnas.1412197111 . ПМК 4210025 . ПМИД 25249632 .
- ^ Кучер Б., Девреотес П., Иглесиас П.А. (февраль 2004 г.). «Локальное возбуждение, глобальный механизм торможения для определения градиента: интерактивный апплет». СТКЭ науки . 2004 (219): табл.3. дои : 10.1126/stke.2192004pl3 . ПМИД 14872096 . S2CID 4660870 .
- ^ Сюн Ю, Хуан Ч., Иглесиас П.А., Девреотес П.Н. (октябрь 2010 г.). «Клетки перемещаются с помощью возбудимой сети с локальным возбуждением и глобальным торможением» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (40): 17079–86. Бибкод : 2010PNAS..10717079X . дои : 10.1073/pnas.1011271107 . ПМЦ 2951443 . ПМИД 20864631 .
- ^ Багорда А., родитель, Калифорния (август 2008 г.). «Краткий обзор хемотаксиса эукариот» . Журнал клеточной науки . 121 (Часть 16): 2621–4. CiteSeerX 10.1.1.515.32 . дои : 10.1242/jcs.018077 . ПМЦ 7213762 . ПМИД 18685153 .
- ^ Кохидай Л (1999). «Хемотаксис: правильный физиологический ответ для оценки филогении сигнальных молекул». Акта Биологика Хунгарика . 50 (4): 375–94. дои : 10.1007/BF03543060 . ПМИД 10735174 . S2CID 248703226 .
- ^ Кедрин Д., ван Ринен Дж., Эрнандес Л., Кондилис Дж., Сегал Дж.Э. (сентябрь 2007 г.). «Подвижность клеток и регуляция цитоскелета при инвазии и метастазировании». Журнал биологии молочной железы и неоплазии . 12 (2–3): 143–52. дои : 10.1007/s10911-007-9046-4 . ПМИД 17557195 . S2CID 31704677 .
- ^ Солница-Крезель Л., Сепич Д.С. (2012). «Гаструляция: образование и формирование зародышевых листков». Ежегодный обзор клеточной биологии и биологии развития . 28 : 687–717. doi : 10.1146/annurev-cellbio-092910-154043 . ПМИД 22804578 . S2CID 11331182 .
- ^ Шеллард А., мэр Р. (июль 2016 г.). «Хемотаксис во время миграции нервного гребня». Семинары по клеточной биологии и биологии развития . 55 : 111–8. дои : 10.1016/j.semcdb.2016.01.031 . ПМИД 26820523 .
- ^ Пал Д.С., Банерджи Т., Лин Ю., де Трогофф Ф., Борлейс Дж., Иглесиас П.А. и др. (июль 2023 г.). «Активация отдельных нижестоящих узлов в сети факторов роста управляет миграцией иммунных клеток» . Развивающая клетка . 58 (13): 1170–1188.e7. дои : 10.1016/j.devcel.2023.04.019 . ПМЦ 10524337 . ПМИД 37220748 .
- ^ Линь Ю, Пал Д.С., Банерджи П., Банерджи Т., Цинь Г., Дэн Ю. и др. (июль 2024 г.). «Подавление Ras усиливает поляризацию и миграцию клеток, обусловленную сократимостью заднего актомиозина». Природная клеточная биология : 1–15. дои : 10.1038/s41556-024-01453-4 . ПМИД 38951708 .
- ^ Беккер Э.Л. (октябрь 1977 г.). «Стимулированное передвижение нейтрофилов: хемокинез и хемотаксис». Архивы патологии и лабораторной медицины . 101 (10): 509–13. ПМИД 199132 .
- ^ Картер С.Б. (январь 1967 г.). «Гаптотаксис и механизм подвижности клеток». Природа . 213 (5073): 256–60. Бибкод : 1967Natur.213..256C . дои : 10.1038/213256a0 . ПМИД 6030602 . S2CID 4212997 .
- ^ Ким Дж. Я., Хаастерт П. В., Девреотес П. Н. (апрель 1996 г.). «Социальные чувства: сигнальные пути рецепторов, связанных с G-белком, у Dictyostelium discoideum» . Химия и биология . 3 (4): 239–243. дои : 10.1016/s1074-5521(96)90103-9 . ПМИД 8807851 .
- ^ Монтелл С. (ноябрь 1999 г.). «Визуальная трансдукция у дрозофилы». Ежегодный обзор клеточной биологии и биологии развития . 15 (1): 231–268. дои : 10.1146/annurev.cellbio.15.1.231 . ПМИД 10611962 . S2CID 14193715 .
- ^ Антунес Дж., Симоэш де Соуза FM (2016). «Передача сигналов обонятельных рецепторов». G-белковые рецепторы – передача сигналов, транспортировка и регуляция . Методы клеточной биологии. Том. 132. стр. 127–45. дои : 10.1016/bs.mcb.2015.11.003 . ISBN 9780128035955 . ПМИД 26928542 .
- ^ Томас М.А., Кляйст А.Б., Фолькман Б.Ф. (август 2018 г.). «Расшифровка хемотаксического сигнала» . Журнал биологии лейкоцитов . 104 (2): 359–374. дои : 10.1002/JLB.1MR0218-044 . ПМК 6099250 . ПМИД 29873835 .
- ^ ван Хаастерт П.Дж., Де Вит Р.Дж., Конейн Т.М. (август 1982 г.). «Антагонисты хемоаттрактантов выявляют отдельные рецепторы цАМФ, фолиевой кислоты и птерина у Dictyostelium» (PDF) . Экспериментальные исследования клеток . 140 (2): 453–6. дои : 10.1016/0014-4827(82)90139-2 . ПМИД 7117406 . S2CID 27784085 .
- ^ Вицани Г., Новацкий М. (2016). Биокоммуникация инфузорий . Спрингер. ISBN 978-3-319-32211-7 . [ нужна страница ]
- ^ Кохидай Л (2016). «Хемотаксис как выражение коммуникации тетрахимен» . В Вицани Г., Новацки М. (ред.). Биокоммуникация инфузорий . Спрингер. стр. 65–82. дои : 10.1007/978-3-319-32211-7_5 . ISBN 978-3-319-32211-7 .
- ^ Кохидай Л., Чаба Г. (июль 1998 г.). «Хемотаксис и хемотаксический отбор, индуцированный цитокинами (IL-8, RANTES и TNF-альфа) у одноклеточных Tetrahymenapyriformis». Цитокин . 10 (7): 481–6. дои : 10.1006/cyto.1997.0328 . ПМИД 9702410 . S2CID 33755476 .
- ^ Зигмонд С.Х. (ноябрь 1977 г.). «Способность полиморфно-ядерных лейкоцитов ориентироваться в градиентах хемотаксических факторов» . Журнал клеточной биологии . 75 (2, часть 1): 606–16. дои : 10.1083/jcb.75.2.606 . ПМК 2109936 . ПМИД 264125 .
- ^ Jump up to: а б с д Пауэлл В.С., Рокач Дж. (апрель 2015 г.). «Биосинтез, биологические эффекты и рецепторы гидроксиэйкозатетраеновых кислот (HETE) и оксоэйкозатетраеновых кислот (оксо-ETE), полученных из арахидоновой кислоты» . Biochimica et Biophysical Acta (BBA) - Молекулярная и клеточная биология липидов . 1851 (4): 340–55. дои : 10.1016/j.bbalip.2014.10.008 . ПМК 5710736 . ПМИД 25449650 .
- ^ Пауэлл В.С., Рокач Дж. (октябрь 2013 г.). «Хемоаттрактант эозинофилов 5-оксо-ETE и рецептор OXE» . Прогресс в исследованиях липидов . 52 (4): 651–65. дои : 10.1016/j.plipres.2013.09.001 . ПМК 5710732 . ПМИД 24056189 .
- ^ Мацуока Т., Нарумия С. (сентябрь 2007 г.). «Передача сигналов рецептором простагландина при заболевании» . Научный мировой журнал . 7 : 1329–47. дои : 10.1100/tsw.2007.182 . ПМК 5901339 . ПМИД 17767353 .
- ^ Ёкомизо Т. (февраль 2015 г.). «Два различных лейкотриеновых B4-рецептора, BLT1 и BLT2». Журнал биохимии . 157 (2): 65–71. дои : 10.1093/jb/mvu078 . ПМИД 25480980 .
- ^ Соццани С., Чжоу Д., Локати М., Бернаскони С., Луини В., Мантовани А. и др. (ноябрь 1996 г.). «Стимулирующие свойства 5-оксо-эйкозаноидов для моноцитов человека: синергизм с моноцитарными хемотаксическими белками-1 и -3» . Журнал иммунологии . 157 (10): 4664–71. дои : 10.4049/jimmunol.157.10.4664 . ПМИД 8906847 . S2CID 23499393 .
- ^ Кохидай Л., Ланг О и Чаба Г. (2003). «Соответствие хемотаксического диапазона аминокислот и его корреляция с физико-химическими параметрами у Tetrahymenapyriformis - Эволюционные последствия». Клеточная и молекулярная биология . 49 : ОЛ487–95. ПМИД 14995080 .
- ^ Jump up to: а б Мюррей Джей Ди (2002). Математическая биология I: Введение (PDF) . Междисциплинарная прикладная математика. Том. 17 (3-е изд.). Нью-Йорк: Спрингер. стр. 395–417. дои : 10.1007/b98868 . ISBN 978-0-387-95223-9 . Архивировано (PDF) из оригинала 6 мая 2022 года.
- ^ Гарасу М., Центлер Ф., Фетцер И., Таллнер М. (2014). «Как хемотаксические характеристики бактерий могут определять структуру их популяций». Биология и биохимия почвы . 69 : 346–358. Бибкод : 2014SBiBi..69..346G . doi : 10.1016/j.soilbio.2013.11.019 .
- ^ Маккензи Д. (6 марта 2023 г.). «Как животные следуют за своим носом» . Знающий журнал . Ежегодные обзоры. doi : 10.1146/knowable-030623-4 . S2CID 257388244 . Проверено 13 марта 2023 г.
- ^ Редди Дж., Мурти В.Н., Вергассола М. (10 марта 2022 г.). «Обонятельное зондирование и навигация в турбулентной среде» . Ежегодный обзор физики конденсированного состояния . 13 (1): 191–213. Бибкод : 2022ARCMP..13..191R . doi : 10.1146/annurev-conmatphys-031720-032754 . ISSN 1947-5454 . S2CID 243966350 .
- ^ Лагзи I (2013). «Химическая робототехника — хемотаксические носители лекарств» . Центральноевропейский медицинский журнал . 8 (4): 377–382. дои : 10.2478/s11536-012-0130-9 . S2CID 84150518 .
- ^ Сенгупта С., Дей К.К., Муддана Х.С., Табуйо Т., Ибеле М.Е., Батлер П.Дж. и др. (январь 2013 г.). «Молекулы ферментов как наномоторы». Журнал Американского химического общества . 135 (4): 1406–1414. дои : 10.1021/ja3091615 . ПМИД 23308365 .
- ^ Мохаджерани Ф, Чжао Х, Сомасундар А, Велегол Д, Сен А (октябрь 2018 г.). «Теория ферментативного хемотаксиса: от экспериментов к моделированию». Биохимия . 57 (43): 6256–6263. arXiv : 1809.02530 . doi : 10.1021/acs.biochem.8b00801 . ПМИД 30251529 . S2CID 52816076 .
- ^ Чжао X, Палаччи Х., Ядав В., Спиринг М.М., Гилсон М.К., Батлер П.Дж. и др. (март 2018 г.). «Хемотаксическая сборка, управляемая субстратом, в ферментном каскаде». Природная химия . 10 (3): 311–317. Бибкод : 2018НатЧ..10..311З . дои : 10.1038/nchem.2905 . ПМИД 29461522 .
- ^ Гуха Р., Мохаджерани Ф., Коллинз М., Гош С., Сен А., Велегол Д. (ноябрь 2017 г.). «Хемотаксис молекулярных красителей в полимерных градиентах в растворе». Журнал Американского химического общества . 139 (44): 15588–15591. дои : 10.1021/jacs.7b08783 . ПМИД 29064685 .
Дальнейшее чтение
[ редактировать ]- Альбертс Б., Джонсон А., Льюис Дж., Уолтер П., Рафф MC (2002). «Бактериальный хемотаксис зависит от двухкомпонентного сигнального пути, активируемого гистидин-киназными рецепторами» . Молекулярная биология клетки . Группа Тейлор и Фрэнсис. ISBN 978-0-8153-4069-0 . Проверено 18 сентября 2017 г.
- Багорда А., родитель, Калифорния (август 2008 г.). «Краткий обзор хемотаксиса эукариот» . Журнал клеточной науки . 121 (Часть 16): 2621–4. CiteSeerX 10.1.1.515.32 . дои : 10.1242/jcs.018077 . ПМЦ 7213762 . ПМИД 18685153 .
- Берг ХК (1993). Случайные блуждания в биологии (Расширенная, переработанная ред.). Принстон, Нью-Джерси: Princeton Univ. Нажимать. ISBN 978-0-691-00064-0 .
- Берг ХК (2003). «Э. coli в движении». Физика сегодня . 58 (2). Нью-Йорк: Спрингер: 64–65. Бибкод : 2005ФТ....58б..64Б . дои : 10.1063/1.1897527 . ISBN 978-0-387-00888-2 .
- Дюзенбери Д.Б. (2009). Жизнь на микроуровне: неожиданная физика маленького размера . Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-03116-6 .
- Айзенбах М. (2004). Ленгелер Дж.В. (ред.). Хемотаксис . Лондон: Издательство Имперского колледжа. ISBN 978-1-86094-413-0 .
- Айзенбах М. (декабрь 2011 г.). «Бактериальный хемотаксис». Энциклопедия наук о жизни . дои : 10.1002/9780470015902.a0001251.pub3 . ISBN 978-0470016176 .
- Hazelbauer GL (13 октября 2012 г.). «Бактериальный хемотаксис: первые годы молекулярных исследований» . Ежегодный обзор микробиологии . 66 (1): 285–303. doi : 10.1146/annurev-micro-092611-150120 . ПМЦ 3989901 . ПМИД 22994495 .
- Джин Т., Херельд Д. (2016). Хемотаксис: методы и протоколы . Хумана Пресс. ISBN 978-1-4939-3480-5 .
- Миллер Л.Д., Рассел М.Х., Александр Г. (2009). «Разнообразие бактериальных хемотаксических реакций и адаптация ниши». Достижения прикладной микробиологии . Том. 66. стр. 53–75. дои : 10.1016/S0065-2164(08)00803-4 . ISBN 9780123747884 . ПМИД 19203648 .
- Рао К.В., Кирби-младший, Аркин А.П. (февраль 2004 г.). «Дизайн и разнообразие бактериального хемотаксиса: сравнительное исследование Escherichia coli и Bacillus subtilis» . ПЛОС Биология . 2 (2): Е49. дои : 10.1371/journal.pbio.0020049 . ПМК 340952 . ПМИД 14966542 .
- Уильямс AH (20 декабря 2010 г.). «Хемотаксис в движении – средство активного обучения» . Журнал микробиологии и биологического образования . 11 (2): 177–8. дои : 10.1128/jmbe.v11i2.216 . ПМЦ 3577161 . ПМИД 23653726 .