Избегание инбридинга
Избегание инбридинга , или гипотеза избежания инбридинга , представляет собой концепцию в эволюционной биологии , которая относится к предотвращению пагубных последствий инбридинга . Животные лишь изредка избегают инбридинга. [1] Гипотеза предотвращения инбридинга утверждает, что определенные механизмы развиваются внутри вида или внутри данной популяции вида в результате ассортативного спаривания , естественного и полового отбора , чтобы предотвратить размножение между родственными особями. Хотя инбридинг может повлечь за собой определенные эволюционные издержки, избегание инбридинга, которое ограничивает количество потенциальных партнеров для данной особи, может привести к альтернативным издержкам. [2] Таким образом, существует баланс между инбридингом и предотвращением инбридинга. Этот баланс определяет, будут ли развиваться механизмы инбридинга и конкретный характер таких механизмов. [3]
Исследование 2007 года показало, что инбредные мыши значительно снизили выживаемость, когда их снова ввели в естественную среду обитания. [4]
Инбридинг может привести к инбридинговой депрессии , которая представляет собой снижение приспособленности данной популяции из-за инбридинга. Инбредная депрессия возникает вследствие появления невыгодных признаков вследствие спаривания вредных рецессивных аллелей спаривающейся пары в потомстве . [5] При спаривании двух родственных особей вероятность спаривания вредных рецессивных аллелей в полученном потомстве выше по сравнению с случаем, когда спариваются неродственные особи, из-за повышенной гомозиготности . Однако инбридинг также дает возможность генетической очистки вредных аллелей, которые в противном случае продолжали бы существовать в популяции и потенциально могли бы увеличиваться в частоте со временем . Другим возможным негативным эффектом инбридинга является ослабление иммунной системы из-за меньшего разнообразия аллелей иммунитета в результате аутбредной депрессии . [6]
Обзор генетики инбредной депрессии в популяциях диких животных и растений, а также у человека привел к выводу, что инбредная депрессия и ее противоположность — гетерозис (гибридная сила) преимущественно обусловлены наличием в популяциях рецессивных вредных аллелей. . [7] Инбридинг , включая самооплодотворение у растений и аутомиктический партеногенез ( телитоки ) у перепончатокрылых , имеет тенденцию приводить к вредному проявлению вредных рецессивных аллелей (инбредная депрессия). Перекрестное оплодотворение между неродственными особями обычно приводит к маскировке вредных рецессивных аллелей в потомстве. [8] [9]
Многие исследования показали, что гомозиготные особи часто находятся в невыгодном положении по сравнению с гетерозиготными. [10] Например, исследование, проведенное на популяции южноафриканских гепардов, показало, что отсутствие генетической изменчивости среди особей в популяции привело к негативным последствиям для особей, таким как более высокий уровень смертности молоди и аномалии сперматозоидов. [11] Когда гетерозиготы обладают преимуществом в приспособленности по сравнению с гомозиготой, популяция с большим количеством гомозигот будет иметь относительно низкую приспособленность, что приведет к инбредной депрессии. Благодаря этим описанным механизмам последствия инбредной депрессии часто бывают достаточно серьезными, чтобы вызвать эволюцию механизмов избегания инбридинга. [12]
Механизмы
[ редактировать ]Механизмы предотвращения инбридинга возникли в ответ на отбор против инбредного потомства. Избегание инбридинга происходит в природе по крайней мере с помощью четырех механизмов: распознавания родственников , расселения, совокуплений экстрапары/экстрагруппы и задержки созревания/подавления репродукции. [3] [12] Эти механизмы не являются взаимоисключающими, и в определенный момент времени в популяции может возникнуть более одного.
Родственное признание
[ редактировать ]

Распознавание родственников — это механизм, с помощью которого люди идентифицируют и избегают спаривания с близкородственными сородичами . Было множество задокументированных примеров того, как люди находили близкородственных особей непривлекательными. В одной серии исследований исследователи сформировали искусственные пары родственников и неродственников (искусственное означает, что они предпочтительно объединяли особей в пары для спаривания в целях экспериментов) и сравнили репродуктивные результаты двух групп. В этих исследованиях парные родственники продемонстрировали снижение воспроизводства и более высокое нежелание спариваться по сравнению с неродственниками. [12] [13] [14] [15] Например, в исследовании Симмонса, посвященном полевым сверчкам, самки сверчков демонстрировали большую задержку спаривания для парных братьев и сестер, сводных братьев и сестер, чем для не-братьев и сестер. [13] В другой серии исследований исследователи позволили людям выбирать себе партнеров из числа сородичей, которые лежат в спектре родства. В этом наборе люди с большей вероятностью выбирали неродственных особей, а не родственных. [12] [14] [16] Например, в исследовании Кракова и др. Самцов диких домашних мышей помещали на арену с четырьмя отдельными отверстиями, ведущими к клеткам с подстилкой из сородичей. Представители сородичей продемонстрировали ряд родственных связей с испытуемыми, а самцы значительно предпочитали постель неродственных братьев и сестер постели родственных самок. [14]
Исследования показали, что распознавание родственников более развито у видов, у которых модели расселения способствуют частым встречам взрослых родственников. [12]
Существует значительное количество вариаций в механизмах, используемых для распознавания родственников. сигналах человека Эти механизмы включают распознавание, основанное на ассоциации или знакомстве, собственных фенотипических , химических сигналах и генах MHC . В рамках механизмов ассоциации/знакомства люди изучают фенотипические профили своих родственников и используют этот шаблон для распознавания родственников. [12] Многие виды достигают этого, «знакомясь» со своими братьями и сестрами, однопометниками или товарищами по гнезду. Эти виды полагаются на то, что потомство выращивается в непосредственной близости, чтобы добиться признания родственников. Это называется эффектом Вестермарка . [17] Например, Холмс и Шерман провели сравнительное исследование арктических сусликов и сусликов Белдинга. Они манипулировали выращенными группами, включив в них как братьев и сестер, так и соседей по гнезду, и обнаружили, что у обоих видов особи были одинаково агрессивны по отношению к своим товарищам по гнезду, независимо от родства. [18] У некоторых видов, где социальные группы очень стабильны, родство и связь между младенцами и другими особями обычно сильно коррелируют. [12] [19] Следовательно, степень ассоциации можно использовать как меру распознавания родственников.
Люди также могут использовать свои собственные характеристики или фенотип в качестве шаблона для распознавания родственников. Например, в одном исследовании Матео и Джонстон вырастили золотистых хомяков только с неродственными особями, а затем заставили их различать запахи родственных и неродственных особей без каких-либо постнатальных встреч с родственниками. Хомяки смогли различать запахи, продемонстрировав использование собственного фенотипа для распознавания родственников. [20] Это исследование также представляет собой пример вида, использующего химические сигналы для распознавания родственников.
Гены главного комплекса гистосовместимости , или гены MHC, участвуют в распознавании родственников. [21] Одна из идей заключается в том, что гены MHC кодируют определенный профиль феромонов для каждого человека, который используется для различения родственных и неродственных сородичей. Несколько исследований продемонстрировали участие генов MHC в распознавании родственников. Например, Мэннинг и др. провели исследование на домашних мышах, в ходе которого изучалось поведение этого вида при совместном гнездовании или выхаживании собственных детенышей, а также детенышей других особей. Как Мэннинг и др. , Согласно теории родственного отбора домовые мыши будут выборочно вскармливать детенышей своих сородичей, чтобы максимизировать инклюзивную приспособленность. Мэннинг и др. продемонстрировали, что домашние мыши используют гены MHC в процессе различения родственников, отдавая предпочтение особям, которые имеют одну и ту же аллельную форму генов MHC. [22]
Признание человеческого рода
[ редактировать ]Возможное использование механизмов, основанных на обонянии, для распознавания родственников человека и предотвращения инбридинга было изучено в трех различных типах исследований. [23] Результаты показали, что обоняние может способствовать развитию в детстве избегания инцеста ( эффект Вестермарка ).
Избегание посткопулятивного инбридинга у мышей
[ редактировать ]Эксперименты с использованием экстракорпорального оплодотворения на мышах предоставили доказательства отбора сперматозоидов на гаметном уровне. [24] Когда сперму самцов- братьев и сестер смешивали, наблюдалось смещение оплодотворения в сторону спермы самцов, не являющихся братьями и сестрами. Результаты были интерпретированы как отбор сперматозоидов, управляемый яйцеклетками, против родственных сперматозоидов.
Предотвращение инбридинга у растений
[ редактировать ]Эксперименты были проведены с двудомным растением Silene latifolia, чтобы проверить, благоприятствует ли после опыления отбор менее родственным донорам пыльцы и снижает ли инбридинг . [25] Результаты показали, что у S. latifolia и, предположительно, в других растительных системах с инбридинговой депрессией отбор пыльцы или эмбрионов после опыления несколькими донорами может снизить инбридинг.
Рассредоточение
[ редактировать ]
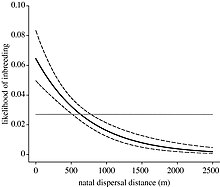
Некоторые виды используют расселение как способ разделения близких родственников и предотвращения инбридинга. [12] Первоначальный путь расселения, по которому могут пойти виды, известен как натальное расселение, при котором особи удаляются от места рождения. Впоследствии виды могут прибегнуть к размножению, при котором особи перемещаются из одной ненатальной группы в другую. Нельсон-Флауэр и др. (2012) провели исследование южных пестрых болтунов и обнаружили, что особи могут путешествовать на большие расстояния от натальных групп, чем от ненатальных групп. [26] Это можно объяснить возможностью встречи сородичей в пределах местных ареалов при расселении. Степень расселения особей конкретного вида зависит от того, смогут ли выгоды от расселения перевесить как издержки инбридинга, так и издержки расселения. Передвижения на дальние расстояния могут нести риски смертности и энергетических затрат. [27]
Рассредоточение по признаку пола
[ редактировать ]Во многих случаях расселения представители одного пола проявляют большую тенденцию покидать свою натальную область, чем представители противоположного пола. [28] Степень предвзятости в отношении определенного пола зависит от множества факторов, которые включают, помимо прочего: систему спаривания, социальную организацию, затраты на инбридинг и расселение, а также физиологические факторы. [27] [28] [29] [30] Когда издержки и выгоды от расселения симметричны как для самцов, так и для самок, тогда не ожидается никакого расселения по признаку пола у видов. [27]
Женское расселение
[ редактировать ]Птицы склонны применять моногамные системы спаривания, при которых самцы остаются в своих натальных группах, чтобы защищать знакомые территории с высоким качеством ресурсов. [28] Самки обычно тратят много энергии при производстве потомства, поэтому инбридинг обходится самкам дорого с точки зрения выживания потомства и репродуктивного успеха. Тогда самки получат больше выгоды, рассредоточившись и выбрав среди этих территориальных самцов. Кроме того, согласно гипотезе Эдипа , дочери самок птиц могут обманывать своих матерей посредством выводкового паразитизма , поэтому самки будут выселять самок из гнезда, заставляя дочерей разбредаться. Расселение самок наблюдается не только у птиц; самцы могут оставаться филопатрическими у млекопитающих, когда средняя продолжительность пребывания взрослых самцов в размножающейся группе превышает средний возраст созревания и зачатия самок. [30] Например, в сообществе шимпанзе в национальном парке Гомбе самцы, как правило, остаются в своем родном сообществе на протяжении всей своей жизни, тогда как самки обычно переезжают в другие сообщества, как только достигают зрелости. [31]
Мужское расселение
[ редактировать ]Расселение самцов чаще встречается у млекопитающих с кооперативным размножением и полигинной системой. Молодые самцы австралийских сумчатых имеют большую тенденцию покидать свои натальные группы, в то время как самки остаются филопатрическими. [32] У Antechinus это связано с тем, что самцы погибают сразу после спаривания; поэтому, когда они расходятся для спаривания, они часто встречаются с женскими натальными группами, в которых отсутствуют самцы. Более того, гипотеза Эдипа также утверждает, что отцы в полигинных системах будут выселять сыновей, имея возможность сделать им рога. [28] Полигинные системы спаривания также влияют на внутриполовую конкуренцию между самцами: в тех случаях, когда самцы могут охранять нескольких самок и проявлять свое доминирование, подчиненные самцы часто вынуждены расходиться по другим ненатальным группам.
Когда виды применяют альтернативные механизмы предотвращения инбридинга, они могут косвенно влиять на то, рассеется ли вид. Их выбор в пользу ненатальной группы самцов затем приводит к расселению самцов.
Задержка созревания
[ редактировать ]Задержка полового созревания потомства в присутствии родителей — еще один механизм, с помощью которого особи избегают инбридинга. Сценарии задержки созревания могут включать удаление исходного родителя противоположного пола, как в случае со львами-самками, у которых течка проявляется раньше после замены их отцов новыми самцами. Другая форма задержки созревания включает присутствие родителей, которое подавляет репродуктивную активность, например, у зрелых игрунок потомство репродуктивно подавляется в присутствии родителей противоположного пола, братьев и сестер в их социальных группах. [12] Репродуктивное подавление происходит, когда половозрелые особи в группе не могут размножаться из-за поведенческих или химических раздражителей со стороны других членов группы, которые подавляют репродуктивное поведение. [33] Социальные сигналы окружающей среды часто диктуют, когда репродуктивная активность подавляется, и включают взаимодействие между взрослыми людьми одного пола. Если текущие условия для воспроизводства неблагоприятны, например, когда в качестве средства воспроизводства используется только инбридинг, люди могут увеличить свой репродуктивный успех в течение всей жизни , приурочивая свои репродуктивные попытки к более благоприятным условиям. Этого можно достичь путем подавления особями своей репродуктивной активности в плохих условиях воспроизводства.
Избегание инбридинга между филопатрическим потомством и их родителями/братьями и сестрами серьезно ограничивает возможности размножения подчиненных, живущих в их социальных группах. Исследование О'Риайна и соавт. (2000) исследовали социальные группы сурикатов и факторы, влияющие на подавление репродуктивной функции у подчиненных самок. Они обнаружили, что в семейных группах отсутствие доминирующей особи любого пола приводило к репродуктивному спокойствию . Репродуктивная деятельность возобновилась только после того, как другая половозрелая самка получила доминирование, а также иммиграция неродственного самца. Для воспроизводства требовалось присутствие неродственного партнера противоположного пола, который действовал как соответствующий стимул на репродуктивно подавленных подчиненных, которые находились в состоянии покоя в присутствии исходной доминирующей особи. [33]

Внепарные совокупления
[ редактировать ]У различных видов самки получают выгоду от спаривания с несколькими самцами, производя таким образом больше потомства с более высоким генетическим разнообразием и потенциально качественным. Самки, которые связаны парой с самцом плохого генетического качества, как это может быть в случае инбридинга, с большей вероятностью будут участвовать в внепарных совокуплениях, чтобы улучшить свой репродуктивный успех и выживаемость своего потомства. [34] Это улучшенное качество потомства возникает либо за счет внутреннего воздействия хороших генов , либо за счет взаимодействия между совместимыми генами родителей. При инбридинге потеря гетерозиготности способствует общему снижению репродуктивного успеха, но когда особи участвуют в внепарных совокуплениях, спаривание между генетически несходными особями приводит к увеличению гетерозиготности. [35]
Внепарные совокупления влекут за собой ряд затрат и выгод как для самцов, так и для самок. Для самцов внепарное совокупление предполагает проведение большего времени вдали от исходной пары в поисках других самок. Это рискует оплодотворить исходную самку другими самцами, пока первоначальный самец ищет партнеров, что приведет к потере отцовства. Компромисс этих затрат полностью зависит от того, сможет ли самец оплодотворить яйца других самок при совокуплении дополнительной пары. Для самок совокупление дополнительных пар гарантирует оплодотворение яйцеклеток и обеспечивает улучшенное генетическое разнообразие с совместимыми сперматозоидами, что позволяет избежать экспрессии повреждающих рецессивных генов, возникающих при инбридинге. [36] Посредством внепарного спаривания самки могут максимизировать генетическую изменчивость своего потомства, обеспечивая защиту от изменений окружающей среды, которые в противном случае могут быть нацелены на более гомозиготные популяции, которые часто возникают в результате инбридинга. [37]
Вступает ли самка в совокупление дополнительной пары ради предотвращения инбридинга, зависит от того, перевешивают ли затраты на совокупление дополнительной пары затраты на инбридинг. При внепарных совокуплениях необходимо учитывать как затраты на инбридинг, так и потерю самцов парной связи (ведущую к потере отцовской заботы) с учетом преимуществ репродуктивного успеха, которые обеспечивает внепарное совокупление. Когда отцовская забота отсутствует или мало влияет на выживаемость потомства, самкам обычно выгодно участвовать в спаривании вне пары, чтобы повысить репродуктивный успех и избежать инбридинга. [34]
Пробелы
[ редактировать ]Избегание инбридинга изучалось тремя основными методами: (1) наблюдение за индивидуальным поведением в присутствии и отсутствии близких родственников, (2) сопоставление издержек избегания с издержками терпимости к близкому инбридингу и (3) сравнение наблюдаемых и случайных частот близких родственников. инбридинг. [38] Ни один метод не является совершенным, что вызывает вопросы о полноте и непротиворечивости гипотезы избежания инбридинга. [38] [39] Хотя первый вариант — индивидуальное наблюдение за поведением — предпочтительнее и наиболее широко используется, до сих пор ведутся споры о том, может ли он предоставить убедительные доказательства предотвращения инбридинга.
Большая часть литературы по предотвращению инбридинга была опубликована не менее 15 лет назад, что позволяет расширять и развивать исследования с помощью современных экспериментальных методов и технологий. Молекулярные методы, такие как дактилоскопия ДНК, стали более совершенными и доступными, что повышает эффективность и точность измерения родства. [12] Изучение избегания инбридинга у плотоядных животных вызвало повышенный интерес благодаря продолжающейся работе по объяснению их социального поведения. [40]
Ссылки
[ редактировать ]- ^ де Бур, Раиса А.; Вега-Трехо, Регина; Котршал, Александр; Фицпатрик, Джон Л. (июль 2021 г.). «Метааналитические доказательства того, что животные редко избегают инбридинга» . Экология и эволюция природы . 5 (7): 949–964. Бибкод : 2021NatEE...5..949D . дои : 10.1038/s41559-021-01453-9 . ISSN 2397-334X . ПМИД 33941905 . S2CID 233718913 .
- ^ Васер П.М., Остад С.Н., Кин Б. (1986). «Когда животные должны терпеть инбридинг?». Американский натуралист . 128 (4): 529–537. дои : 10.1086/284585 . S2CID 84806782 .
- ^ Jump up to: а б Арчи Э.А., Холлистер-Смит Дж.А., Пул Дж.Х., Ли ПК, Мосс СиДжей, Мальдонадо Дж.Э., Альбертс С.К. (2007). «Избегание поведенческого инбридинга у диких африканских слонов». Молекулярная экология . 16 (19): 4138–4148. Бибкод : 2007MolEc..16.4138A . дои : 10.1111/j.1365-294x.2007.03483.x . ПМИД 17784925 . S2CID 1535829 .
- ^ Хименес Х.А., Хьюз К.А. , Алакс Г., Грэм Л., Лейси Р.К. (октябрь 1994 г.). «Экспериментальное исследование инбредной депрессии в естественной среде обитания». Наука . 266 (5183): 271–273. Бибкод : 1994Sci...266..271J . дои : 10.1126/science.7939661 . ПМИД 7939661 .
- ^ Мохаммад Афзал (январь 1983 г.). «Влияние кровного родства на коэффициент интеллекта и неонатальное поведение детей-мусульман ансари» .
- ^ Соммер, С. (2005). «Важность изменчивости иммунных генов (MHC) в эволюционной экологии и сохранении природы» . Границы в зоологии . 2:16 . дои : 10.1186/1742-9994-2-16 . ПМЦ 1282567 . ПМИД 16242022 .
- ^ Чарльзворт Д., Уиллис Дж. Х. (2009). «Генетика инбредной депрессии». Нат. Преподобный Жене . 10 (11): 783–96. дои : 10.1038/nrg2664 . ПМИД 19834483 . S2CID 771357 .
- ^ Бернштейн Х., Хопф Ф.А., Мишод Р.Э. (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–70. дои : 10.1016/s0065-2660(08)60012-7 . ISBN 9780120176243 . ПМИД 3324702 .
- ^ Мишод, RE (1994). «Эрос и эволюция: естественная философия секса», Издательство Addison-Wesley, Ридинг, Массачусетс. ISBN 978-0201442328
- ^ Црнокрак П., Рофф Д.А. (1999). «Инбредная депрессия в дикой природе» . Наследственность . 83 (3): 260–270. дои : 10.1038/sj.hdy.6885530 . ПМИД 10504423 .
- ^ О'Брайен С.Дж., Ролке М.Е., Маркер Л. (1985). «Генетическая основа видовой уязвимости гепарда». Наука . 227 (4693): 1428–1434. Бибкод : 1985Sci...227.1428O . дои : 10.1126/science.2983425 . ПМИД 2983425 . S2CID 14341795 .
- ^ Jump up to: а б с д и ж г час я дж к Пьюзи А., Вольф М. (1996). «Избежание инбридинга у животных». Тенденции Эколь Эвол . 11 (5): 201–206. Бибкод : 1996TEcoE..11..201P . дои : 10.1016/0169-5347(96)10028-8 . ПМИД 21237809 .
- ^ Jump up to: а б Симмонс, Л.В. (1989) Распознавание родственников и его влияние на брачные предпочтения полевого сверчка Gryffus bimaculatus (de Geer), Anim. Поведение. 38,68-77
- ^ Jump up to: а б с Краков, С. и Матушак, Б. (1991) Выбор партнера для неродственных братьев и сестер у диких домашних мышей: данные теста выбора и репродуктивного теста , Этология 88,99-108
- ^ Боллинджер, EK et al. (1991)Избежание инбридинга у луговой полевки (Microtus pennsylvanicus), .I Mammal. 72, 419-421
- ^ Кин, Б. (1990) Влияние родства на репродуктивный успех и выбор партнера у белоногой мыши Peromyscus leucopus, Anim. Поведение. 39,264-273
- ^ Вольф, AP Вестермарк Redivivus. Ежегодный обзор антропологии 22: 157–175, 1993 г.
- ^ Холмс В.Г., Шерман П.В. (1982) Онтогенез распознавания родства у двух видов сусликов . Американский зоолог , 22 491–517.
- ^ Пьюзи, А.Е. (1990) Механизмы предотвращения инбридинга у приматов, не являющихся человеком, в книге «Педофилия: биосоциальные измерения» (Фейрман, Дж.Р., ред.), стр. 201-220, Springer-Verlag.
- ^ Матео Дж. М., Джонстон Р. Е. (2000) Распознавание родственников и «эффект подмышки»: свидетельства самореферентного совпадения фенотипов . Труды Лондонского королевского общества. Серия Б, Биологические науки , 267, 695–700.
- ^ Джеррам Л. Браун и Эми Эклунд «Американский натуралист», том. 143, № 3 (март 1994 г.), стр. 435–461. Опубликовано: The University of Chicago Press.
- ^ Мэннинг С.Дж., Уэйкленд Э.К., Поттс В.К. (1992). «Схемы коллективного гнездования у мышей указывают на участие генов MHC в распознавании родственников». Природа . 360 (6404): 581–583. Бибкод : 1992Natur.360..581M . дои : 10.1038/360581a0 . ПМИД 1461279 . S2CID 4335928 .
- ^ Вайсфельд Г.Е., Чилли Т., Филлипс К.А., Галл Дж.А., Лихтман С.М. (2003). «Возможные механизмы распознавания родственников человека и предотвращения инбридинга, основанные на обонянии». J Exp Детская Психология . 85 (3): 279–95. дои : 10.1016/s0022-0965(03)00061-4 . ПМИД 12810039 .
- ^ Фирман Р.К., Симмонс Л.В. (2015). «Гаметические взаимодействия способствуют предотвращению инбридинга у домашних мышей». Экол. Летт . 18 (9): 937–43. Бибкод : 2015EcolL..18..937F . дои : 10.1111/ele.12471 . ПМИД 26154782 .
- ^ Тейшейра С., Ферстер К., Бернаскони Г. (2009). «Доказательства инбридинговой депрессии и отбора после опыления против инбридинга у двудомного растения Silene latifolia» . Наследственность (Эдинб) . 102 (2): 101–12. дои : 10.1038/hdy.2008.86 . ПМИД 18698334 .
- ^ Нельсон-Флауэр М.Дж., Хоккей П.А., О'Райан С., Ридли А.Р. (2012). «Механизмы предотвращения инбридинга: динамика расселения при совместном разведении южных пестрых болтунов» . Журнал экологии животных . 81 (4): 876–883. Бибкод : 2012JAnEc..81..876N . дои : 10.1111/j.1365-2656.2012.01983.x . ПМИД 22471769 .
- ^ Jump up to: а б с Перрен Н., Мазалов В. (1999). «Рассредоточение и предотвращение инбридинга». Американский натуралист . 154 (3): 282–292. дои : 10.1086/303236 . ПМИД 10506544 . S2CID 42975760 .
- ^ Jump up to: а б с д Пьюзи А.Е. (1987). «Рассредоточение по признаку пола и предотвращение инбридинга у птиц и млекопитающих». Тенденции в экологии и эволюции . 2 (10): 295–299. Бибкод : 1987TEcoE...2..295P . дои : 10.1016/0169-5347(87)90081-4 . ПМИД 21227869 .
- ^ Леманн Л., Перрин Н. (2003). «Избегание инбридинга посредством признания родства: разборчивые самки способствуют расселению самцов» . Американский натуралист . 162 (5): 638–652. дои : 10.1086/378823 . ПМИД 14618541 . S2CID 25356698 .
- ^ Jump up to: а б Клаттон-Брок, TH (1989). «Передача самок и предотвращение инбридинга у социальных млекопитающих». Природа . 337 (6202): 70–72. Бибкод : 1989Natur.337...70C . дои : 10.1038/337070a0 . ПМИД 2909891 . S2CID 4343108 .
- ^ Пьюзи А.Е. (1980). «Избежание инбридинга у шимпанзе». Поведение животных . 28 (2): 543–552. дои : 10.1016/s0003-3472(80)80063-7 . S2CID 53164905 .
- ^ Кокберн А., Скотт, член парламента, Скоттс-ди-джей (1985). «Избегание инбридинга и натальное расселение по признаку самцов у Antechinus spp. (Marsupialia: Dasyuridae)». Поведение животных . 33 (3): 908–915. дои : 10.1016/s0003-3472(85)80025-7 . S2CID 53206203 .
- ^ Jump up to: а б О'Риэйн М.Дж., Беннетт, Северная Каролина, Браттон П.Н., Макилрат Г.М., Клаттон-Брок, Т.Х. (2000b). «Подавление репродукции и предотвращение инбридинга в диких популяциях совместно размножающихся сурикатов Suricata suricatta». Поведение. Экол. Социобиол . 48 (6): 471–477. дои : 10.1007/s002650000249 . S2CID 25928636 .
- ^ Jump up to: а б Петри М., Кемпенаерс Б. (1998). «Внепарное отцовство у птиц: объяснение различий между видами и популяциями». Тенденции экологии и эволюции . 13 (2): 52–57. Бибкод : 1998TEcoE..13...52P . дои : 10.1016/s0169-5347(97)01232-9 . ПМИД 21238200 .
- ^ Ферстер К., Дели К., Джонсен А., Лифьелд Дж.Т., Кемпенаерс Б. (октябрь 2003 г.). «Самки повышают гетерозиготность и приспособленность потомства за счет дополнительных парных спариваний». Природа . 425 (6959): 714–7. Бибкод : 2003Natur.425..714F . дои : 10.1038/nature01969 . ПМИД 14562103 . S2CID 4311810 .
- ^ Алкок, Джон. 1998. Поведение животных. Шестое издание. 429-519.
- ^ Морелл С. (1998). «Новый взгляд на моногамию». Наука . 281 (5385): 1982–1983. дои : 10.1126/science.281.5385.1982 . ПМИД 9767050 . S2CID 31391458 .
- ^ Jump up to: а б Парт, Т. (1996). Проблемы с тестированием предотвращения инбридинга: случай мухоловки с ошейником. Эволюция, 1625-1630 гг.
- ^ Мур Дж., Али Р. (1984). «Связаны ли расселение и предотвращение инбридинга?». Поведение животных . 32 (1): 94–112. CiteSeerX 10.1.1.584.5226 . дои : 10.1016/s0003-3472(84)80328-0 . S2CID 7674864 .
- ^ Кин Б., Крил С.Р., Васер П.М. (1996). «Нет свидетельств избегания инбридинга или инбредной депрессии у социального хищника» . Поведенческая экология . 7 (4): 480–489. дои : 10.1093/beheco/7.4.480 .