белок арабиногалактан
Арабиногалактан-белки ( AGP ) представляют собой высокогликозилированные белки ( гликопротеины ), обнаруженные в клеточных стенках растений . Каждый из них состоит из белка , к которому прикреплены молекулы сахара (который может составлять более 90% общей массы). Они являются членами более широкого класса гликопротеинов клеточной стенки, богатых гидроксипролином (Hyp), - большой и разнообразной группы гликозилированных белков стенки.
АГП обнаружены у многих высших растений в семенах , корнях , стеблях , листьях и соцветиях . АГП составляют лишь небольшую часть клеточной стенки, обычно не более 1% сухой массы первичной стенки. О них также сообщалось в секретах клеточной культуральной среды корней, листьев, эндосперма и тканей эмбриона, а некоторые типы клеток, продуцирующих экссудат, такие как клетки шиловидного канала, способны продуцировать большие количества AGP. Они участвуют в различных аспектах роста и развития растений, включая удлинение корней , соматический эмбриогенез , гормональные реакции , дифференцировку ксилемы , рост и управление пыльцевыми трубками , запрограммированную гибель клеток , расширение клеток, толерантность к соли , взаимодействия хозяина и патогена и передачу клеточных сигналов .
AGP привлекли значительное внимание из-за своей очень сложной структуры и потенциальной роли в передаче сигналов. Кроме того, они находят применение в промышленности и здравоохранении благодаря своим химическим/физическим свойствам (водоудерживающая способность, адгезия и эмульгирование).
Последовательность и классификация
[ редактировать ]
Белковый компонент AGP богат аминокислотами пролином (P), аланином (A), серином (S) и треонином (T), также известными как «PAST», и это смещение аминокислот является одной из особенностей, используемых для идентифицировать их. [ 1 ] [ 2 ] [ 3 ] [ 4 ] AGP по своей сути являются неупорядоченными белками, поскольку они содержат большое количество разупорядочивающих аминокислот, таких как пролин, которые нарушают образование стабильных складчатых структур. AGP, характерные для внутренне неупорядоченных белков, также содержат повторяющиеся мотивы и посттрансляционные модификации . [ 2 ] [ 5 ] Остатки пролина в основной цепи белка могут быть гидроксилированы до гидроксипролина (O) в зависимости от окружающих аминокислот. «Гипотеза смежности Hyp» [ 6 ] [ 2 ] [ 3 ] предсказывает, что когда O встречается несмежным образом, например, последовательность «SOTO», такая как встречается в AGP, это действует как сигнал для O -связанного гликозилирования больших разветвленных полисахаридов арабиногалактана (AG) типа II. [ 7 ] Последовательности, которые направляют гликозилирование AG (SO, TO, AO, VO), называются гликомотивами AGP.
Все скелеты белка AGP содержат минимум 3 кластерных гликомотива AGP и N-концевой сигнальный пептид , который направляет белок в эндоплазматический ретикулум (ER), где начинаются посттрансляционные модификации. [ 8 ] Пролилгидроксилирование P до O осуществляется пролил-4-гидроксилазами (P4Hs), принадлежащими к семейству 2-оксоглутарат-зависимых диоксигеназ. [ 9 ] P4H был идентифицирован как в ER, так и в аппарате Гольджи. [ 10 ] Добавление гликозилфосфатидилинозитол ( GPI)-якоря происходит в большинстве, но не во всех AGP. [ 3 ] [ 4 ]
Семьи
[ редактировать ]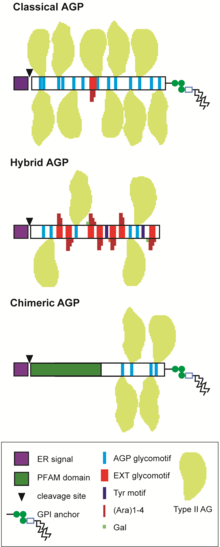
AGP относятся к большим мультигенным семействам и делятся на несколько подгрупп в зависимости от предсказанной последовательности белка. [ 11 ] [ 4 ] [ 12 ] [ 13 ] [ 2 ] [ 14 ] «Классические» AGP включают GPI-AGP, которые состоят из сигнального пептида на N-конце, богатой PAST последовательности из 100-150 аминокислот и гидрофобной области на C-конце, которая управляет добавлением GPI-якоря; не GPI-AGP, у которых отсутствует C-концевая сигнальная последовательность GPI, AGP, богатые лизином (K), которые содержат богатую K область внутри богатого PAST остова, и AG-пептид, который имеет короткий богатый PAST остов из 10-15 аа ( рис. 2 ). Химерные AGP состоят из белков, которые имеют область AGP и дополнительную область с известным доменом семейства белков ( Pfam ). Химерные AGP включают фасциклиноподобные AGP (FLA), фитоцианиноподобные AGP (PAG/PLA, также известные как ранние нодулинподобные белки, ENODL) и ксилогеноподобные AGP (XYLP), которые содержат домены, подобные переносу липидов . [ 1 ] Было идентифицировано несколько других предполагаемых химерных классов AGP, которые включают гликомотивы AG, связанные с протеинкиназой , богатыми лейцином повторами , X8 , FH2 и другими доменами семейства белков . [ 2 ] [ 15 ] [ 16 ] Существуют и другие неклассические AGP, например, содержащие домен, богатый цистеином (C), также называемый доменами PAC, и/или домен, богатый гистидином (H), [ 17 ] [ 18 ] а также многие гибридные HRGP, которые имеют мотивы, характерные для AGP и других членов HRGP, обычно экстенсина и Tyr . мотивы [ 17 ] [ 19 ] [ 1 ] [ 2 ] [ 14 ] AGP эволюционно древние и были идентифицированы у зеленых водорослей , а также у Chromista и Glaucophyta . [ 2 ] [ 14 ] [ 20 ] Предполагается, что наземные растения, обнаруженные во всей линии растений , унаследовали и диверсифицировали существующие гены основной цепи белка AGP, присутствующие в водорослях, с образованием огромного количества гликоформ AGP.
Структура
[ редактировать ]Углеводные фрагменты AGP богаты арабинозой и галактаном , но могут быть обнаружены и другие сахара, такие как L-рамнопираноза (L-Rha p ), D-маннопираноза (Man p ), D-ксилопираноза (Xyl p ), L-фукоза. (Fuc), D-глюкопираноза (Glc p ), D-глюкуроновая кислота (GlcA) и ее 4-O-метиловое производное, а также D-галактуроновая кислота (GalA) и ее 4-O-метиловое производное. [ 21 ] [ 22 ] AG, обнаруженный в AGP, относится к типу II (AG типа II), то есть представляет собой галактановый остов из (1-3)-связанных остатков β-D-галактопиранозы (Gal p ) с разветвлениями (длиной от одного до трех остатков). (1,6)-связанного β-D - Гал с. В большинстве случаев остатки Gal оканчиваются остатками α-L-арабинофуранозы (Ara f ). Некоторые AGP богаты уроновыми кислотами (GlcA), что приводит к образованию заряженного полисахаридного фрагмента, а другие содержат короткие олигосахариды Ara f . [ 23 ] За синтез этого комплекса отвечают специфические наборы гидроксипролин-O-β-галактозилтрансфераз, β-1,3-галактозилтрансфераз, β-1,6-галактозилтрансфераз, α-арабинозилтрансфераз, β-глюкуронозилтрансфераз, α-рамнозилтрансфераз и α-фукозилтрансфераз. структуры. [ 24 ]
Одной из особенностей АГ типа II, особенно (1,3)-связанных остатков β-D-Gal p , является их способность связываться с фенилгликозидами Ярива. Фенилгликозиды Ярива широко используются в качестве цитохимических реагентов для нарушения молекулярных функций AGP, а также для обнаружения, количественного определения, очистки и окрашивания AGP. [ 21 ] Недавно сообщалось, что взаимодействие с Яривом не обнаружено для β-1,6-галактоолигосахаридов любой длины. [ 25 ] Был сделан вывод, что фенилгликозиды Ярива являются специфически связывающими реагентами для цепей β-1,3-галактана длиной более пяти остатков. Семи остатков и более достаточно для перекрестного связывания, что приводит к осаждению гликанов фенилгликозидами Ярива, что наблюдается при связывании классических AGP с красителями β-Ярива. Те же результаты наблюдались, когда в AGP, по-видимому, требовалось как минимум 5–7 β-1,3-связанных единиц Gal для образования агрегатов с реагентом Ярива. [ 26 ]
Биосинтез
[ редактировать ]После трансляции скелеты белков AGP богато украшены сложными углеводами, в первую очередь полисахаридами AG типа II. [ 27 ] Биосинтез зрелого AGP включает расщепление сигнального пептида на N-конце, гидроксилирование остатков P и последующее гликозилирование и во многих случаях добавление GPI-якоря.
Обработка и транспортировка
[ редактировать ]гликозилирование Предполагается, что основной цепи AGP инициируется в ER с добавлением первой Gal с помощью O -галактозилтрансферазы, которая преимущественно локализована во фракциях ER . [ 28 ] Расширение цепи тогда происходит преимущественно в ГА . [ 29 ] Для тех AGP, которые включают привязку GPI, добавление происходит во время котрансляционной миграции в ER.
Боковые цепи арабиногалактана
[ редактировать ]Структура гликанов AG состоит из основной цепи β -1,3-связанной галактозы (Gal), с боковыми цепями β -1,6-связанной Gal и концевых остатков арабинозы (Ara), рамнозы (Rha), Gal, фукозы. (Fuc) и глюкуроновая кислота (GlcA). Эти фрагменты AG-гликанов собираются гликозилтрансферазами (GT). [ 30 ] O -гликозилирование AGP инициируется действием Hyp- O -галактозилтрансфераз (Hyp- O -GalTs), которые добавляют первую Gal к белку. Сложные структуры гликанов затем вырабатываются набором гликозилтрансфераз, большинство из которых биохимически не охарактеризованы. Семейство GT31 является одним из семейств, участвующих в биосинтезе гликанового остова AGP. [ 31 ] [ 32 ] У многих членов семейства GT31 была обнаружена активность Hyp- O -GALT. [ 33 ] [ 34 ] и основная цепь β- (1,3)-галактана, вероятно, также будет синтезироваться семейством GT31. [ 32 ] Члены семейства GT14 участвуют в добавлении β- (1,6)- и β- (1,3) -галактанов к AGP. [ 35 ] [ 36 ] Предполагается, что у Arabidopsis концевые сахара, такие как фукоза, добавляются AtFUT4 (фукозилтрансферазой ) и AtFUT6 в семействе GT37. [ 37 ] [ 38 ] и включение терминального GlcA может катализироваться семейством GT14. [ 35 ] [ 39 ] Ряд GT еще предстоит идентифицировать, например, те, которые отвечают за терминал Rha.
GPI-якорь
[ редактировать ]Биоинформатический анализ предсказывает добавление GPI-якоря на многие AGP. [ 4 ] Ранний синтез фрагмента GPI происходит на цитоплазматической поверхности ЭР, а последующая сборка происходит в просвете ЭР. Они включают сборку триманнозы ( Man), галактозы, не-N-ацетилированного глюкозамина (GlcN) и фосфата этаноламина с образованием зрелого фрагмента GPI. [ 40 ] [ 41 ] AGP подвергаются добавлению якоря GPI во время котрансляционной миграции в ER, и эти два процесса наконец сходятся. Впоследствии комплекс трансамидазы одновременно расщепляет коровый белок на С-конце, когда он распознает сайт расщепления ω, и переносит полностью собранный якорь GPI на аминокислотный остаток на С-конце белка. Эти события происходят до гидроксилирования и гликозилирования пролила. [ 42 ] [ 10 ] Основной гликановой структурой якорей GPI является Man- α -1,2-Man- α -1,6-Man- α -1,4-GlcN-инозитол (Man: манноза, GlcN: глюкозаминил), который консервативен во многих эукариоты . [ 41 ] [ 43 ] [ 44 ] [ 40 ] [ 10 ] [ 45 ] Единственная якорная структура GPI растений, охарактеризованная на сегодняшний день, - это GPI-заякоренный AGP из клеток Pyrus communis , культивируемых в суспензии. [ 40 ] Это показало частично модифицированный гликановый фрагмент по сравнению с ранее охарактеризованными якорями GPI, поскольку он содержал β -1,4-Gal. Предполагается, что путь синтеза якоря GPI и сборки белка консервативен у млекопитающих и растений. [ 10 ] Интеграция GPI-якоря позволяет прикрепить белок к мембране ЭР, проходящей через ГА, что приводит к секреции во внешний листок плазматической мембраны, обращенный к стенке. [ 46 ] По предложению Оксли и Бачича, [ 40 ] GPI-заякоренные AGP, вероятно, высвобождаются посредством расщепления некоторыми фосфолипазами (PL) (C или D) и секретируются во внеклеточный компартмент.
Функционально охарактеризованные гены, участвующие в гликозилировании AGP
[ редактировать ]Биоинформатический анализ с использованием последовательностей β-1,3-галактозилтрансферазы (GalT) млекопитающих в качестве матриц показал участие семейства гликозилтрансфераз (GT) 31 , активирующих углеводы (CAZy), в синтезе галактановых цепей основной цепи AG. [ 47 ] Члены семейства GT31 были сгруппированы в 11 клад, из которых четыре клады являются специфичными для растений: клады 1, 7, 10 и 11. Домены и мотивы клад 1 и 11 четко не определены; в то время как клады 7 и 10 имеют сходство доменов с белками с известной функцией GalT в системах млекопитающих. [ 47 ] Белки клады 7 содержат домены как GalT, так и галектина, тогда как белки клады 10 содержат GalT-специфичный домен. [ 48 ] Предполагается, что домен галектина позволяет GalT связываться с первым остатком Gal на полипептидном остове AGP; таким образом, определяя положение последующих остатков Gal на основной цепи белка, аналогично активности белков, содержащих домен галектина человека. [ 47 ]
Восемь ферментов, принадлежащих к семейству GT31, продемонстрировали способность размещать первый остаток Gal на остатках Hyp в коровых белках AGP. Эти ферменты называются GALT2, GALT3, GALT4, GALT5, GALT6. [ 49 ] которые являются членами клада 7, а также HPGT1, HPGT2 и HPGT3, [ 50 ] которые являются членами Клада 10. Предварительные исследования субстратной специфичности фермента показали, что другой фермент GT31 клады 10, At1g77810, обладает активностью β-1,3-GalT. [ 47 ] Ген GT31 клады 10, KNS4/UPEX1 , кодирует β-1,3-GalT, способный синтезировать связи β-1,3-Gal, обнаруженные в AG типа II, присутствующих в AGP и/или пектиновом рамногалактуронане I (RG-I). [ 51 ] Другой член клады 10 GT31, названный GALT31A , кодирует β-1,6-GalT при гетерологичной экспрессии в E. coli и Nicotiana benthamiana , а также удлиненные боковые цепи β-1,6-галактана гликанов AGP. [ 52 ] GALT29A, член семейства GT29, был идентифицирован как экспрессирующийся совместно с GALT31A, действующий совместно и образующий комплексы. [ 53 ]
Сообщалось, что три члена GT14, названные GlcAT14A, GlcAT14B и GlcAT14C, добавляли GlcA к цепям β-1,6- и β-1,3-Gal в ферментном анализе in vitro после гетерологичной экспрессии в Pichia Pastoris . [ 54 ] Два гена α-фукозилтрансферазы, FUT4 и FUT6 , оба принадлежащие к семейству GT37, кодируют ферменты, которые добавляют остатки α-1,2-фукозы к AGP. [ 55 ] [ 56 ] Они кажутся частично избыточными, поскольку демонстрируют несколько разные специфичности субстрата AGP. [ 55 ] Было обнаружено, что член семейства GT77, REDUCED ARABINOSE YARIV (RAY1), представляет собой β-арабинозилтрансферазу, которая присоединяет β-Ara f к метиловой β-Gal полимера стенки, осаждаемого Яривом. [ 57 ] Ожидается, что дополнительные исследования позволят функционально идентифицировать другие гены, участвующие в гликозилировании AGP, и их взаимодействие с другими компонентами клеточной стенки растений.
Биологические роли
[ редактировать ]Использование человеком AGP включает использование гуммиарабика в пищевой и фармацевтической промышленности из-за его природных свойств загущать и эмульгировать . [ 58 ] [ 59 ] AGP в зернах зерновых имеют потенциальное применение в биофортификации . [ 60 ] в качестве источников пищевых волокон для поддержания кишечных бактерий [ 61 ] и защитные средства от токсичности этанола . [ 62 ]
AGP обнаруживаются в широком спектре растительных тканей, в секретах клеточной культуральной среды корней , листьев , эндосперма и эмбриона тканей , а также в некоторых типах клеток, продуцирующих экссудат , таких как клетки шиловидного канала . [ 19 ] [ 63 ] Было показано, что AGP регулируют многие аспекты роста и развития растений , включая распознавание мужских и женских особей в органах размножения, деление и дифференцировку клеток в эмбриональном и постэмбриональном развитии , слизи семян развитие клеточной стенки , толерантность корней к соли и взаимодействие корневых микробов . [ 64 ] [ 10 ] [ 65 ] Эти исследования показывают, что они многофункциональны, подобно тому, что обнаружено в протеогликанах/гликопротеинах млекопитающих. [ 66 ] [ 67 ] [ 68 ] Традиционные методы изучения функций AGP включают использование β -гликозиловых (обычно глюкозильных) реагентов Ярива и моноклональных антител (мАт). Реагенты β -гликозил Ярива представляют собой синтетические фенилазогликозидные зонды, которые специфически, но не ковалентно, связываются с AGP и могут использоваться для осаждения AGP из раствора. [ 69 ] Их также обычно используют в качестве гистохимических красителей для определения местоположения и распределения AGP. [ 70 ] [ 71 ] Ряд исследований показал, что добавление реагентов β -Ярива в питательную среду растений может ингибировать рост проростков, удлинение клеток, блокировать соматический эмбриогенез и накопление массы свежих клеточных стенок. [ 72 ] [ 73 ] [ 74 ] Использование моноклональных антител, которые специфически связываются с углеводными эпитопами AGP, также применялось для определения функций на основе местоположения и структуры эпитопов AGP. [ 75 ] Обычно используемые mAb против AGP включают CCRC-M7, LM2, JIM8, JIM13 и JIM14. [ 76 ]
Функция отдельных AGP в значительной степени была установлена посредством исследований мутантов . Например, арабидопсиса было показано, что специфичный для корня AtAGP30 необходим для регенерации корня in vitro, что указывает на его функцию в регенерации корня путем модуляции активности фитогормонов . [ 77 ] Исследования мутантов agp6 и agp11 у Arabidopsis продемонстрировали важность этих AGP для предотвращения неконтролируемого образования пыльцевого зерна и для нормального роста пыльцевой трубки . [ 78 ] [ 79 ] Функциональные механизмы AGP в передаче сигналов в клетках недостаточно изучены. Одна предложенная модель предполагает, что AGP могут взаимодействовать и контролировать высвобождение кальция из AG-гликана (через остатки GlcA), запуская нижестоящие сигнальные пути, опосредованные кальцием . [ 80 ] [ 81 ] [ 82 ] Другой возможный механизм, в основном основанный на изучении FLA, предполагает, что комбинация домена фасциклина и гликанов AG может опосредовать межклеточную адгезию. [ 83 ] [ 84 ]
| Биологическая роль | АГП [ а ] [ б ] | Местоположение(а) | Функция(и) | Ссылки |
|---|---|---|---|---|
| Эмбриогенез | GhPLA1 | Соматические эмбрионы | Содействие соматическому эмбриогенезу | [ 85 ] |
| ДкАГП | Соматические эмбрионы | Содействие соматическому эмбриогенезу | [ 86 ] | |
| AtAGP | Эмбрионы | Развитие и дифференциация эмбрионов | [ 87 ] | |
| NtAGP | Эмбрионы | Развитие эмбриона | [ 88 ] | |
| BgAGP | Соматические эмбрионы | Скорость развития и морфология соматических эмбрионов | [ 89 ] | |
| БнАГП | Эмбрионы | Развитие эмбриона | [ 21 ] | |
| МаАГП | Соматические эмбрионы | Содействие соматическому эмбриогенезу | [ 90 ] | |
| ПсАГП | Содействие соматическому эмбриогенезу | [ 91 ] | ||
| ФсАГП | Эмбрионы | Образование и стабильность клеточной стенки | [ 20 ] | |
| ВКАЛГАЛ-САМ | эмбрионы | Адгезия эмбриональных клеток | [ 83 ] | |
| ВК ISG | эмбрионы | Инверсия эмбриона | [ 92 ] | |
| Воспроизведение | AtAGP4 (ЯГГЕР) | Пестик | Закупорка пыльцевой трубки | [ 93 ] |
| AtAGP6, AtAGP11 | Тычинка, пыльцевое зерно и пыльцевая трубка | Развитие пыльцевого зерна и рост пыльцевой трубки | [ 94 ] [ 95 ] | |
| AtAGP18 | яйцеклетка | Выбор мегаспор | [ 96 ] [ 97 ] | |
| AtFLA3 | Пыльцевое зерно и пыльцевая трубка | Развитие микроспор | [ 98 ] | |
| AtENODL11-15 | Микропиляр | Прием пыльцевой трубки | [ 99 ] [ 100 ] | |
| БкмМФ8 | Пыльцевое зерно и пыльцевая трубка | Развитие пыльцевой стенки и рост пыльцевой трубки | [ 101 ] | |
| БкмМФ18 | Пыльцевое зерно | Развитие пыльцевого зерна, образование интины | [ 102 ] | |
| НтТТС | Пестик | Рост пыльцевой трубки и руководство | [ 103 ] | |
| Нп/На120кД | Пестик | S-специфическое отторжение пыльцы (самонесовместимость) | [ 104 ] | |
| ОсМТР1 | Мужские репродуктивные клетки | Развитие пыльников и фертильность пыльцы | [ 105 ] | |
| Развитие завода | AtAGP19 | Стебель, цветок, корень и лист | Деление и расширение клеток, развитие листьев и размножение | [ 106 ] |
| AtAGP57C | Розеточный лист, кремень, семя, цветок и верхушка побега стебля соцветия. | Поддержание структуры клеточной стенки | [ 107 ] | |
| AtFLA1 | Устьица, трихомы, сосудистая сеть листа, кончик главного корня и боковой корень. | Развитие бокового корня и регенерация побегов | [ 108 ] | |
| AtFLA4 (SOS5) | Цветок, лист, стебель, корень, кремень | Устойчивость корней к солевому стрессу; прилипание семенной слизи | [ 109 ] [ 110 ] [ 111 ] [ 112 ] [ 113 ] | |
| PpAGP1 | Апикальные клетки | Расширение апикальных клеток | [ 114 ] | |
| AtAGP30 | Корень | Регенерация корней и прорастание семян | [ 47 ] | |
| BcrFLA1 | Корень | Удлинение корневых волос | [ 115 ] | |
| Развитие вторичной стенки | AtFLA11, AtFLA12 | Ствол и ветвь | Синтез/паттерн вторичной клеточной стенки | [ 116 ] |
| AtXYP1, AtXYP2 | Клеточные стенки дифференцирующихся элементов трахеи | Развитие и формирование сосудистой ткани | [ 117 ] | |
| GhAGP4 | Хлопковое волокно | Инициирование и удлинение хлопкового волокна | [ 98 ] | |
| GhFLA1 | Хлопковое волокно | Инициация и удлинение волокон | [ 52 ] | |
| PtFLA6 | Стеблевое ксилемное волокно | Синтез/паттерн вторичной клеточной стенки | [ 118 ] | |
| Оборона | СлаттАГП | Место атаки паразитов | Способствует приживанию паразитов | [ 119 ] |
| Растительно-микробное взаимодействие | AtAGP17 | Корень | Agrobacterium tumefaciens Трансформация корня | [ 120 ] |
- ^ Gh: Gossypium hirsutum , Dc: Daucus carota , At: Arabidopsis thaliana , Nt: Nicotiana tabacum , Bg: Bactris gasipaes , Bn: Brassica napus , Ma: Musa spp. AAA , Ps: Pelargonium sidoides , Fs: Fucus serratus , Vc: Volvox carteri , Bcm: Brassica Campestris , Np: Nicotiana Plumbaginifolia , Na: Nicotiana alata , Os: Oryza sativa , Pp: Physcomitrella patens , Bcr: Brassica carinata , Pt: Populus trichocarpa , Sl: Solanum licopersicum .
- ^ PLA: фитоцианиноподобный AGP. ALGAL-CAM: молекула адгезии клеток водорослей. ISG: гликопротеин, специфичный для инверсии. FLA: фасциклин, подобный AGP. ЭНОДЛ: Похоже на узелковые образования. МФ: мужская фертильность. TTS: передача тканеспецифической. MTR: регулятор микроспор и тапетума. SOS: соль слишком чувствительна. XYP: белок ксилоген. attAGP: AGP вложения.
Функции AGP в процессах роста и развития растений во многом зависят от невероятного разнообразия их гликановых и белковых фрагментов основной цепи. [ 121 ] В частности, именно полисахариды AG, скорее всего, будут участвовать в развитии. [ 122 ] Большая часть биологической роли AGP была идентифицирована посредством характеристики инсерционных мутантов Т-ДНК генов или ферментов, участвующих в гликозилировании AGP, в первую очередь у Arabidopsis thaliana . Одиночные мутанты galt2-6 выявили некоторые физиологические фенотипы при нормальных условиях роста, включая уменьшение длины и плотности корневых волосков, уменьшение завязывания семян, уменьшение прилипшей слизи семенной оболочки и преждевременное старение. [ 123 ] Однако двойные мутанты galt2galt5 показали более тяжелые и плейотропные физиологические фенотипы, чем одиночные мутанты, в отношении длины и плотности корневых волосков и слизи семенной оболочки. [ 123 ] Аналогичным образом, тройные мутанты hpgt1hpgt2hpgt3 показали несколько плейотропных фенотипов, включая более длинные боковые корни, увеличенную длину и плотность корневых волосков, более толстые корни, меньшие розеточные листья, более короткие черешки, более короткие стебли соцветий, пониженную плодовитость и более короткие кремнеземы. [ 50 ] В случае GALT31A было обнаружено, что он необходим для развития эмбрионов Arabidopsis . Вставка Т-ДНК в 9-й экзон GALT31A привела к гибели эмбрионов этой мутантной линии. [ 52 ] Между тем, нокаутные мутанты по KNS4/UPEX1 имеют коллапс пыльцевых зерен и аномальную структуру и морфологию пыльцевой экзины. [ 124 ] Кроме того, одиночные мутанты kns4 демонстрировали пониженную фертильность, подтверждая, что KNS4/UPEX1 имеет решающее значение для жизнеспособности и развития пыльцы. [ 51 ] Нокаутные мутанты по FUT4 и FUT6 показали сильное ингибирование роста корней в условиях соли. [ 56 ] в то время как нокаутные мутанты по GlcAT14A , GlcAT14B и GlcAT14C показали повышенную скорость удлинения клеток в выращенных в темноте гипокотилях и выращенных на свету корнях во время роста проростков. [ 125 ] В случае сеянцев с мутантом ray1 повлияла на длину первичного корня , выращенных на вертикальных чашках, мутация RAY1 . Кроме того, первичный корень мутантов ray1 рос медленнее по сравнению с арабидопсисом дикого типа . [ 57 ] В совокупности эти исследования доказывают, что правильное гликозилирование AGP важно для функционирования AGP в процессе роста и развития растений.
Человеческое использование
[ редактировать ]Использование AGP человеком включает использование гуммиарабика в пищевой и фармацевтической промышленности из-за его естественных свойств загущения и эмульгирования . [ 58 ] [ 59 ] AGP в зернах зерновых имеют потенциальное применение в биофортификации . [ 60 ] в качестве источников пищевых волокон для поддержания кишечных бактерий [ 61 ] и защитные средства от токсичности этанола . [ 62 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]![]() Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2021 г. ) ( отчеты рецензента ):
Инсюань Ма; Ким Джонсон (15 января 2021 г.). «Арабиногалактан-белки» (PDF) . Викижурнал науки . 4 (1): 2. doi : 10.15347/WJS/2021.002 . ISSN 2470-6345 . Викиданные Q99557488 .
Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2021 г. ) ( отчеты рецензента ):
Инсюань Ма; Ким Джонсон (15 января 2021 г.). «Арабиногалактан-белки» (PDF) . Викижурнал науки . 4 (1): 2. doi : 10.15347/WJS/2021.002 . ISSN 2470-6345 . Викиданные Q99557488 .
- ^ Jump up to: а б с Шоуолтер, Аллан М.; Кепплер, Брайан; Лихтенберг, Йенс; Гу, Дачжан; Уэлч, Лонни Р. (15 апреля 2010 г.). «Биоинформатический подход к идентификации, классификации и анализу гликопротеинов, богатых гидроксипролином» . Физиология растений . 153 (2): 485–513. дои : 10.1104/стр.110.156554 . ISSN 0032-0889 . ПМЦ 2879790 . ПМИД 20395450 .
- ^ Jump up to: а б с д и ж г Джонсон, Ким Л.; Кассен, Эндрю М.; Лонсдейл, Эндрю; Бачич, Антоний; Доблин, Моника С.; Шульц, Кэролайн Дж. (26 апреля 2017 г.). «Конвейер для идентификации гликопротеинов, богатых гидроксипролином» . Физиология растений . 174 (2): 886–903. дои : 10.1104/стр.17.00294 . ISSN 0032-0889 . ПМК 5462032 . ПМИД 28446635 .
- ^ Jump up to: а б с Джонсон, Ким Л.; Кассен, Эндрю М.; Лонсдейл, Эндрю; Вонг, Гейн Ка-Шу; Солтис, Дуглас Э.; Майлз, Николас В.; Мелконян, Михаил; Мелконян, Барбара; Дейхолос, Майкл К. (26 апреля 2017 г.). «Эволюция богатых гидроксипролином гликопротеинов на основе 1000 растительных транскриптомов» . Физиология растений . 174 (2): 904–921. дои : 10.1104/стр.17.00295 . ISSN 0032-0889 . ПМК 5462033 . ПМИД 28446636 .
- ^ Jump up to: а б с д Шульц, Кэролайн Дж.; Румсевич, Майкл П.; Джонсон, Ким Л.; Джонс, Брайан Дж.; Гаспар, Иоланда М.; Бачич, Энтони (1 августа 2002 г.). «Использование геномных ресурсов для определения направлений исследований. Семейство генов белка арабиногалактана как тестовый пример» . Физиология растений . 129 (4): 1448–1463. дои : 10.1104/стр.003459 . ISSN 0032-0889 . ПМК 166734 . ПМИД 12177459 .
- ^ Шафи, Томас; Бачич, Антоний; Джонсон, Ким (01 августа 2020 г.). Вилке, Клаус (ред.). «Эволюция неупорядоченных областей с разнообразной последовательностью в семействе белков: порядок внутри хаоса» . Молекулярная биология и эволюция . 37 (8): 2155–2172. дои : 10.1093/molbev/msaa096 . ISSN 0737-4038 . ПМИД 32359163 .
- ^ Келишевский, Марсия Дж.; Лэмпорт, Дерек Т.А. (февраль 1994 г.). «Экстенсин: повторяющиеся мотивы, функциональные сайты, посттрансляционные коды и филогения» . Заводской журнал . 5 (2): 157–172. дои : 10.1046/j.1365-313x.1994.05020157.x . ISSN 0960-7412 . PMID 8148875 .
- ^ Тан, Ли; Лейкам, Джозеф Ф.; Келишевский, Марсия Дж. (12 июня 2003 г.). «Мотивы гликозилирования, которые направляют добавление арабиногалактана к белкам арабиногалактана» . Физиология растений . 132 (3): 1362–1369. дои : 10.1104/стр.103.021766 . ISSN 0032-0889 . ПМК 167076 . ПМИД 12857818 .
- ^ Шульц, Кэролайн; Гилсон, Пол; Оксли, Дэвид; Юл, Джоэлиан; Бачич, Энтони (ноябрь 1998 г.). «GPI-якоря на арабиногалактановых белках: значение для передачи сигналов в растениях» . Тенденции в науке о растениях . 3 (11): 426–431. дои : 10.1016/s1360-1385(98)01328-4 . ISSN 1360-1385 .
- ^ Коски, М. Кристиан; Хиета, Рейя; Бёлльнер, Клаудия; Кивирикко, Кари И.; Мюллихарью, Йоханна; Веренга, Рик К. (21 декабря 2007 г.). «Активный центр пролил-4-гидроксилазы водорослей обладает большой структурной пластичностью» . Журнал биологической химии . 282 (51): 37112–37123. дои : 10.1074/jbc.M706554200 . ISSN 0021-9258 . ПМИД 17940281 .
- ^ Jump up to: а б с д и Эллис, Мириам; Эгелунд, Джек; Шульц, Кэролайн Дж.; Бачич, Энтони (13 апреля 2010 г.). «Арабиногалактан-белки: ключевые регуляторы на поверхности клетки?» . Физиология растений . 153 (2): 403–419. дои : 10.1104/стр.110.156000 . ISSN 0032-0889 . ПМЦ 2879789 . ПМИД 20388666 .
- ^ Шоуолтер, AM (сентябрь 2001 г.). «Арабиногалактан-белки: структура, выражение и функции» . Клеточные и молекулярные науки о жизни . 58 (10): 1399–1417. дои : 10.1007/pl00000784 . ISSN 1420-682X . ПМИД 11693522 . S2CID 206858189 .
- ^ Гаспар, Иоланда Мария; Нам, Джесон; Шульц, Кэролин Джейн; Ли, Лань-Ин; Гилсон, Пол Р.; Гелвин, Стэнтон Б.; Бачич, Энтони (30 июля 2004 г.). «Характеристика мутанта арабидопсиса, богатого лизином арабиногалактан-белка AtAGP17 (крыса1), который приводит к снижению эффективности трансформации агробактерий» . Физиология растений . 135 (4): 2162–2171. дои : 10.1104/стр.104.045542 . ISSN 0032-0889 . ПМК 520787 . ПМИД 15286287 .
- ^ Хуан, Гэн-Цин; Гонг, Си-Ин; Сюй, Вэнь-Лян; Ли, Вэнь; Ли, Пэн; Чжан, Чао-Цзюнь; Ли, Дэн-Ди; Чжэн, Юн; Ли, Фу-Гуан (24 января 2013 г.). «Фасциклиноподобный белок арабиногалактана, GhFLA1, участвует в инициировании волокон и удлинении хлопка» . Физиология растений . 161 (3): 1278–1290. дои : 10.1104/стр.112.203760 . ISSN 0032-0889 . ПМЦ 3585596 . ПМИД 23349362 .
- ^ Jump up to: а б с Джонсон, Ким Л.; Кассен, Эндрю М.; Лонсдейл, Эндрю; Вонг, Гейн Ка-Шу; Солтис, Дуглас Э.; Майлз, Николас В.; Мелконян, Михаил; Мелконян, Барбара; Дейхолос, Майкл К. (26 апреля 2017 г.). «Эволюция богатых гидроксипролином гликопротеинов на основе 1000 растительных транскриптомов» . Физиология растений . 174 (2): 904–921. дои : 10.1104/стр.17.00295 . ISSN 0032-0889 . ПМК 5462033 . ПМИД 28446636 .
- ^ Драгичевич, Милан Б; Паунович, Даниела М; Богданович, Милица Д; Тодорович, Сладжана I; Симонович, Ана Д (01 января 2020 г.). «ragp: Трубопровод для добычи растительных гликопротеинов, богатых гидроксипролином, с реализацией в R» . Гликобиология . 30 (1): 19–35. дои : 10.1093/гликоб/cwz072 . ISSN 1460-2423 . ПМИД 31508799 .
- ^ Пфайфер, Лукас; Шафи, Томас; Джонсон, Ким Л.; Бачич, Антоний; Классен, Биргит (декабрь 2020 г.). «Арабиногалактан-белки Zostera marina L. содержат уникальные гликановые структуры и позволяют понять процессы адаптации к соленой среде» . Научные отчеты . 10 (1): 8232. Бибкод : 2020NatSR..10.8232P . дои : 10.1038/s41598-020-65135-5 . ISSN 2045-2322 . ПМЦ 7237498 . ПМИД 32427862 .
- ^ Jump up to: а б Болдуин, Тимоти К.; Доминго, Конча; Шиндлер, Томас; Ситхараман, Гури; Стейси, Никола; Робертс, Кейт (2001). «DcAGP1, секретируемый белок арабиногалактан, относится к семейству основных белков, богатых пролином» . Молекулярная биология растений . 45 (4): 421–435. дои : 10.1023/А:1010637426934 . hdl : 2436/17032 . ПМИД 11352461 . S2CID 8322072 .
- ^ Нгуен-Ким, Хуан; Сан-Клементе, Элен; Лаймер, Йозеф; Лакнер, Питер; Гадермайер, Габриэле; Дюнан, Кристоф; Жаме, Элизабет (03 апреля 2020 г.). «Белки домена PAC клеточной стенки (богатые пролином, арабиногалактановыми белками, консервативными цистеинами) консервативны в зеленой линии» . Международный журнал молекулярных наук . 21 (7): 2488. doi : 10.3390/ijms21072488 . ISSN 1422-0067 . ПМЦ 7177597 . ПМИД 32260156 .
- ^ Jump up to: а б Гаспар, Ю.; Джонсон, КЛ; Маккенна, Дж.А.; Бачич, А.; Шульц, CJ (сентябрь 2001 г.). «Сложные структуры белков арабиногалактана и путь к пониманию их функции» . Молекулярная биология растений . 47 (1–2): 161–176. дои : 10.1023/А:1010683432529 . ISSN 0167-4412 . ПМИД 11554470 . S2CID 19541545 .
- ^ Jump up to: а б Эрве, Сесиль; Симеон, Амандин; Джем, Мюриэль; Кассен, Эндрю; Джонсон, Ким Л.; Салмеан, Армандо А.; Уиллатс, Уильям Г.Т.; Доблин, Моника С.; Бачич, Энтони (15 декабря 2015 г.). «Белки арабиногалактана имеют глубокие корни у эукариот: идентификация генов и эпитопов у бурых водорослей и их роль в развитии эмбриона Fucus serratus» . Новый фитолог . 209 (4): 1428–1441. дои : 10.1111/nph.13786 . ISSN 0028-646X . ПМИД 26667994 .
- ^ Jump up to: а б с Финчер, Великобритания; Стоун, бакалавр; Кларк, А.Е. (1983). «Арабиногалактан-белки: структура, биосинтез и функции». Ежегодный обзор физиологии растений . 34 (1): 47–70. дои : 10.1146/annurev.pp.34.060183.000403 .
- ^ Инаба, Михо; Йошими, Ёсихиса; Мацуока, Кодзи; Трифона, Теодора; Цумурая, Йоичи (13 октября 2015 г.) . -белок в листьях редиса» . Carbodrive Research . 415 : 1–11. doi : /j.carres.2015.07.002 . ISSN 0008-6215 . PMC 4610949. 10.1016 PMID 26267887 .
- ^ Ду, Он; Кларк, Эдриенн Э.; Бачич, Энтони (1 ноября 1996 г.). «Арабиногалактан-белки: класс протеогликанов внеклеточного матрикса, участвующих в росте и развитии растений». Тенденции в клеточной биологии . 6 (11): 411–414. дои : 10.1016/S0962-8924(96)20036-4 . ISSN 0962-8924 . ПМИД 15157509 .
- ^ Шоуолтер, Аллан М.; Басу, Дебарати (2016). «Биосинтез белка экстенсина и арабиногалактана: гликозилтрансферазы, проблемы исследования и биосенсоры» . Границы в науке о растениях . 7 : 814. doi : 10.3389/fpls.2016.00814 . ISSN 1664-462X . ПМК 4908140 . ПМИД 27379116 .
- ^ Китадзава, Киминари; Трифона, Феодора; Ёшими, Ёсихиса; Хаяси, Ёсихиро; Каваучи, Сусуму; Антонов, Людмил; Танака, Хироши; Такахаси, Такаши; Канеко, Сатоши (01 марта 2013 г.). «Реагент β-галактозил Ярива связывается с β-1,3-галактаном белков арабиногалактана» . Физиология растений . 161 (3): 1117–1126. дои : 10.1104/стр.112.211722 . ISSN 0032-0889 . ПМЦ 3585584 . ПМИД 23296690 .
- ^ Полсен, бакалавр наук; Крейк, диджей; Данстан, Делавэр; Стоун, бакалавр; Бачич, А. (15 июня 2014 г.). «Реагент Ярива: поведение в различных растворителях и взаимодействие с белком арабиногалактана гуммиарабика». Углеводные полимеры . 106 : 460–468. дои : 10.1016/j.carbpol.2014.01.009 . ISSN 0144-8617 . ПМИД 24721102 .
- ^ Лян, Ян; Басу, Дебарати; Паттатил, Сивакумар; Сюй, Вэньлян; Венетос, Александра; Мартин, Стэнтон Л.; Фаик, Ахмед; Хан, Майкл Г.; Шоуолтер, Аллан М. (14 октября 2013 г.). «Биохимическая и физиологическая характеристика мутантов fut4 и fut6, дефектных в фукозилировании белка арабиногалактана у арабидопсиса» . Журнал экспериментальной ботаники . 64 (18): 5537–5551. дои : 10.1093/jxb/ert321 . ISSN 1460-2431 . ПМЦ 3871811 . ПМИД 24127514 .
- ^ Ока, Такудзи; Сайто, Фуми; Шимма, Ёичи; Ёко-о, Такэхико; Номура, Ёсиюки; Мацуока, Кен; Дзигами, Ёсифуми (18 ноября 2009 г.). «Характеристика UDP-d-галактозы, локализованной в эндоплазматическом ретикулуме: гидроксипролин-О-галактозилтрансферазы с использованием синтетических пептидных субстратов арабидопсиса» . Физиология растений . 152 (1): 332–340. дои : 10.1104/стр.109.146266 . ISSN 0032-0889 . ПМЦ 2799367 . ПМИД 19923238 .
- ^ Като, Хидеаки; Такеучи, Ёшими; Цумурая, Ёичи; Хасимото, Ёхичи; Накано, Хирофуми; Ковач, Павол (11 февраля 2003 г.). «Биосинтез галактанов in vitro с помощью мембраносвязанной галактозилтрансферазы из проростков редиса (Raphanus sativus L.)» . Планта . 217 (2): 271–282. Бибкод : 2003Завод.217..271К . дои : 10.1007/s00425-003-0978-7 . ISSN 0032-0935 . ПМИД 12783335 . S2CID 5783849 .
- ^ Шоуолтер, Аллан М.; Басу, Дебарати (15 июня 2016 г.). «Биосинтез белка экстенсина и арабиногалактана: гликозилтрансферазы, проблемы исследования и биосенсоры» . Границы в науке о растениях . 7 : 814. doi : 10.3389/fpls.2016.00814 . ISSN 1664-462X . ПМК 4908140 . ПМИД 27379116 .
- ^ Эгелунд, Джек; Обель, Николай; Ульвсков, Петр; Геши, Наоми; Поли, Маркус; Бачич, Антоний; Петерсен, Бент Ларсен (31 марта 2007 г.). «Молекулярная характеристика двух мутантов гликозилтрансферазы Arabidopsis thaliana, rra1 и rra2, которые имеют пониженное остаточное содержание арабинозы в полимере, тесно связанном с остатком целлюлозной стенки» . Молекулярная биология растений . 64 (4): 439–451. дои : 10.1007/s11103-007-9162-y . ISSN 0167-4412 . ПМИД 17401635 . S2CID 11643754 .
- ^ Jump up to: а б Цюй, Юнмей; Эгелунд, Джек; Гилсон, Пол Р.; Хоутон, Фиона; Глисон, Пол А.; Шульц, Кэролайн Дж.; Бачич, Энтони (12 июня 2008 г.). «Идентификация новой группы предполагаемых β-(1,3)-галактозилтрансфераз Arabidopsis thaliana» . Молекулярная биология растений . 68 (1–2): 43–59. дои : 10.1007/s11103-008-9351-3 . ISSN 0167-4412 . ПМИД 18548197 . S2CID 25896609 .
- ^ Басу, Дебарати; Тиан, Лу; Ван, Вуда; Боббс, Шауни; Херок, Хейли; Трэверс, Эндрю; Шоуолтер, Аллан М. (декабрь 2015 г.). «Небольшое мультигенное семейство гидроксипролин-О-галактозилтрансфераз участвует в гликозилировании, росте и развитии арабиногалактан-белка» . Биология растений BMC . 15 (1): 295. дои : 10.1186/s12870-015-0670-7 . ISSN 1471-2229 . ПМЦ 4687291 . ПМИД 26690932 .
- ^ Огава-Охниши, Мари; Мацубаяси, Ёсикацу (24 февраля 2015 г.). «Идентификация трех мощных гидроксипролин-О-галактозилтрансфераз у арабидопсиса» . Заводской журнал . 81 (5): 736–746. дои : 10.1111/tpj.12764 . ISSN 0960-7412 . ПМИД 25600942 .
- ^ Jump up to: а б Кнох, Ева; Дилопимол, Адифол; Трифона, Феодора; Поульсен, Кристиан П.; Сюн, Гуанъянь; Хархолт, Джеспер; Петерсен, Бент Л.; Вольфсков, Питер; Хади, Масуд З. (29 ноября 2013 г.). «β-глюкуронозилтрансфераза из Arabidopsis thaliana, участвующая в биосинтезе арабиногалактана II типа, играет роль в удлинении клеток во время роста проростков» . Заводской журнал . 76 (6): 1016–1029. дои : 10.1111/tpj.12353 . ISSN 0960-7412 . ПМИД 24128328 .
- ^ Дилокпимол, Адифол; Геши, Наоми (16 апреля 2014 г.). «Arabidopsis thalianaguronosyltransferase в семействе GT14» . Сигнализация и поведение растений . 9 (6): e28891. Бибкод : 2014PlSiB...9E8891D . дои : 10.4161/psb.28891 . ISSN 1559-2324 . ПМК 4091549 . ПМИД 24739253 .
- ^ У, Инъин; Уильямс, Мэтью; Бернар, Софи; Дриуич, Азеддин; Шоуолтер, Аллан М.; Фаик, Ахмед (30 апреля 2010 г.). «Функциональная идентификация двух неизбыточных альфа-(1,2)фукозилтрансфераз Arabidopsis, специфичных к белкам арабиногалактана» . Журнал биологической химии . 285 (18): 13638–13645. дои : 10.1074/jbc.M110.102715 . ISSN 1083-351X . ПМЦ 2859526 . ПМИД 20194500 .
- ^ Трифона, Феодора; Тейс, Тина Э.; Вагнер, Таня; Стотт, Кэтрин; Кигстра, Кеннет; Дюпри, Поль (25 марта 2014 г.). «Характеристика α-(1 → 2)-фукозилтрансфераз FUT4 и FUT6 показывает, что отсутствие фукозилирования корневого арабиногалактана увеличивает чувствительность корня арабидопсиса к солям» . ПЛОС ОДИН . 9 (3): e93291. Бибкод : 2014PLoSO...993291T . дои : 10.1371/journal.pone.0093291 . ISSN 1932-6203 . ПМЦ 3965541 . ПМИД 24667545 .
- ^ Дилокпимол, Адифол; Поульсен, Кристиан; Вереб, Дьёрдь; Канеко, Сатоши; Шульц, Александр; Геши, Наоми (2014). «Галактозилтрансферазы Arabidopsis thaliana в биосинтезе арабиногалактана II типа: молекулярное взаимодействие усиливает активность фермента» . Биология растений BMC . 14 (1): 90. дои : 10.1186/1471-2229-14-90 . ISSN 1471-2229 . ПМЦ 4234293 . ПМИД 24693939 .
- ^ Jump up to: а б с д Оксли, Д.; Бачич, А. (7 декабря 1999 г.). «Структура гликозилфосфатидилинозитолового якоря белка арабиногалактана из клеток Pyrus communis, культивируемых в суспензии» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (25): 14246–14251. Бибкод : 1999PNAS...9614246O . дои : 10.1073/pnas.96.25.14246 . ISSN 0027-8424 . ПМК 24422 . ПМИД 10588691 .
- ^ Jump up to: а б Йейтс, Тревор Х.; Бачич, Антоний; Джонсон, Ким Л. (август 2018 г.). «Растительные белки, заякоренные на гликозилфосфатидилинозитоле, в связи с плазматической мембраной и клеточной стенкой: растительные белки, заякоренные с GPI» . Журнал интегративной биологии растений . 60 (8): 649–669. дои : 10.1111/jipb.12659 . hdl : 11343/283902 . ISSN 1744-7909 . ПМИД 29667761 . S2CID 4949024 .
- ^ Имхоф, Изабелла; Флюри, Изабель; Вионне, Кристина; Рубати, Кэрол; Эггер, Дайан; Конзельманн, Андреас (7 мая 2004 г.). «Гликозилфосфатидилинозитол (GPI) белки Saccharomyces cerevisiae содержат фосфатные группы этаноламина на альфа1,4-связанной маннозе якоря GPI» . Журнал биологической химии . 279 (19): 19614–19627. дои : 10.1074/jbc.M401873200 . ISSN 0021-9258 . ПМИД 14985347 .
- ^ Фергюсон, М.; Хоманс, С.; Двек, Р.; Радемахер, Т. (12 февраля 1988 г.). «Гликозил-фосфатидилинозитоловый фрагмент, который прикрепляет поверхностный гликопротеин варианта Trypanosoma brucei к мембране» . Наука . 239 (4841): 753–759. Бибкод : 1988Sci...239..753F . дои : 10.1126/science.3340856 . ISSN 0036-8075 . ПМИД 3340856 .
- ^ Фергюсон, Массачусетс (сентябрь 1999 г.). «Структура, биосинтез и функции гликозилфосфатидилинозитоловых якорей, а также вклад исследований трипаносом» . Журнал клеточной науки . 112 (17): 2799–2809. дои : 10.1242/jcs.112.17.2799 . ISSN 0021-9533 . ПМИД 10444375 .
- ^ Штрассер, Ричард (23 февраля 2016 г.). «Гликозилирование растительных белков» . Гликобиология . 26 (9): 926–939. дои : 10.1093/гликоб/cww023 . ISSN 0959-6658 . ПМЦ 5045529 . ПМИД 26911286 .
- ^ Мунис, М.; Сурзоло, К. (6 июня 2014 г.). «Сортировка GPI-заякоренных белков от дрожжей до млекопитающих – общие пути в разных местах?» . Журнал клеточной науки . 127 (13): 2793–2801. дои : 10.1242/jcs.148056 . ISSN 0021-9533 . ПМИД 24906797 .
- ^ Jump up to: а б с д и Цюй, Юнмей; Эгелунд, Джек; Гилсон, Пол Р.; Хоутон, Фиона; Глисон, Пол А.; Шульц, Кэролайн Дж.; Бачич, Энтони (1 сентября 2008 г.). «Идентификация новой группы предполагаемых β-(1,3)-галактозилтрансфераз Arabidopsis thaliana». Молекулярная биология растений . 68 (1–2): 43–59. дои : 10.1007/s11103-008-9351-3 . ISSN 0167-4412 . ПМИД 18548197 . S2CID 25896609 .
- ^ Эгелунд, Джек; Эллис, Мириам; Доблин, Моника; Цюй, Юнмей; Бачич, Энтони (2010). Ульвсков, Петр (ред.). Ежегодные обзоры растений . Уайли-Блэквелл. стр. 213–234. дои : 10.1002/9781444391015.ch7 . ISBN 9781444391015 .
- ^ Басу, Дебарати; Лян, Ян; Лю, Сяо; Химмельдирк, Клаус; Фаик, Ахмед; Келишевский, Марсия; Держись, Майкл; Шоуолтер, Аллан М. (5 апреля 2013 г.). «Функциональная идентификация гидроксипролин-О-галактозилтрансферазы, специфичной для биосинтеза белка арабиногалактана у Arabidopsis» . Журнал биологической химии . 288 (14): 10132–10143. дои : 10.1074/jbc.m112.432609 . ISSN 0021-9258 . ПМЦ 3617256 . ПМИД 23430255 .
- ^ Jump up to: а б Огава-Охниши, Мари; Мацубаяси, Ёсикацу (01 марта 2015 г.). «Идентификация трех мощных гидроксипролин-О-галактозилтрансфераз у арабидопсиса» . Заводской журнал . 81 (5): 736–746. дои : 10.1111/tpj.12764 . ISSN 1365-313X . ПМИД 25600942 .
- ^ Jump up to: а б Сузуки, Тошия; Нарцисс, Джоан; Цзэн, Вэй; Мин, Эллисон ван де; Ясутоми, Масаюки; Такемура, Сюнсукэ; Молния, Эдвин Р.; Доблин, Моника С.; Бачич, Энтони (01 января 2017 г.). «KNS4 / UPEX1: арабиногалактан β-(1,3)-галактозилтрансфераза типа II, необходимая для развития экзины пыльцы» . Физиология растений . 173 (1): 183–205. дои : 10.1104/стр.16.01385 . ISSN 0032-0889 . ПМК 5210738 . ПМИД 27837085 .
- ^ Jump up to: а б с Геши, Наоми; Йохансен, Йорунн Н.; Дилокпимол, Адифол; Роллан, Аурелия; Белкрам, Катя; Верже, Стефан; Котаке, Тошихиса; Цумурая, Ёичи; Канеко, Сатоши (01 октября 2013 г.). «Галактозилтрансфераза, действующая на гликаны белка арабиногалактана, необходима для развития эмбрионов Arabidopsis». Заводской журнал . 76 (1): 128–137. дои : 10.1111/tpj.12281 . ISSN 1365-313X . ПМИД 23837821 .
- ^ Дилокпимол, Адифол; Поульсен, Кристиан Питер; Вереб, Дьёрдь; Канеко, Сатоши; Шульц, Александр; Геши, Наоми (3 апреля 2014 г.). «Галактозилтрансферазы Arabidopsis thaliana в биосинтезе арабиногалактана II типа: молекулярное взаимодействие усиливает активность фермента» . Биология растений BMC . 14:90 . дои : 10.1186/1471-2229-14-90 . ISSN 1471-2229 . ПМЦ 4234293 . ПМИД 24693939 .
- ^ Дилокпимол, Адифол; Геши, Наоми (01 июня 2014 г.). «Глюкуронозилтрансфераза Arabidopsis thaliana в семействе GT14» . Сигнализация и поведение растений . 9 (6): e28891. Бибкод : 2014PlSiB...9E8891D . дои : 10.4161/psb.28891 . ПМК 4091549 . ПМИД 24739253 .
- ^ Jump up to: а б У, Инъин; Уильямс, Мэтью; Бернар, Софи; Дриуич, Азеддин; Шоуолтер, Аллан М.; Фаик, Ахмед (30 апреля 2010 г.). «Функциональная идентификация двух неизбыточных α(1,2)фукозилтрансфераз Arabidopsis, специфичных для белков арабиногалактана» . Журнал биологической химии . 285 (18): 13638–13645. дои : 10.1074/jbc.m110.102715 . ISSN 0021-9258 . ПМЦ 2859526 . ПМИД 20194500 .
- ^ Jump up to: а б Лян, Ян; Басу, Дебарати; Паттатил, Сивакумар; Сюй, Вэньлян; Венетос, Александра; Мартин, Стэнтон Л.; Фаик, Ахмед; Хан, Майкл Г.; Шоуолтер, Аллан М. (1 декабря 2013 г.). «Биохимическая и физиологическая характеристика мутантов fut4 и fut6, дефектных в фукозилировании белка арабиногалактана у арабидопсиса» . Журнал экспериментальной ботаники . 64 (18): 5537–5551. дои : 10.1093/jxb/ert321 . ISSN 0022-0957 . ПМЦ 3871811 . ПМИД 24127514 .
- ^ Jump up to: а б Гилле, Саша; Шарма, Вайшали; Байду, Эдвард ЭК; Кислинг, Джей Д.; Шеллер, Хенрик Вибе; Поли, Маркус (01 июля 2013 г.). «Арабинозилирование осаждаемого Яривом полимера клеточной стенки влияет на рост растений на примере мутанта гликозилтрансферазы Arabidopsis ray1» . Молекулярный завод . 6 (4): 1369–1372. дои : 10.1093/mp/sst029 . ISSN 1674-2052 . ПМИД 23396039 .
- ^ Jump up to: а б Саха, Дипджьоти; Бхаттачарья, Сувенду (декабрь 2010 г.). «Гидроколлоиды как загустители и гелеобразователи в пищевых продуктах: критический обзор» . Журнал пищевой науки и технологий . 47 (6): 587–597. дои : 10.1007/s13197-010-0162-6 . ISSN 0022-1155 . ПМЦ 3551143 . ПМИД 23572691 .
- ^ Jump up to: а б Барклай, Томас Г.; Дэй, Кэндис Минту; Петровский, Николай; Гарг, Санджай (октябрь 2019 г.). «Обзор доставки функциональных лекарств на основе полисахаридных частиц» . Углеводные полимеры . 221 : 94–112. дои : 10.1016/j.carbpol.2019.05.067 . ПМК 6626612 . ПМИД 31227171 .
- ^ Jump up to: а б Айзат, Ван М.; Пройсс, Джеймс М.; Джонсон, Александр А.Т.; Тестер, Марк А.; Шульц, Кэролин Дж. (ноябрь 2011 г.). «Исследование богатого His белка арабиногалактана для биообогащения микроэлементами зерна злаков» . Физиология Плантарум . 143 (3): 271–286. дои : 10.1111/j.1399-3054.2011.01499.x . ПМИД 21707638 .
- ^ Jump up to: а б Фудзита, Киётака; Сасаки, Юки; Китахара, Канефуми (сентябрь 2019 г.). «Деградация растительных белков арабиногалактана кишечными бактериями: характеристики и функции участвующих ферментов» . Прикладная микробиология и биотехнология . 103 (18): 7451–7457. дои : 10.1007/s00253-019-10049-0 . ISSN 0175-7598 . ПМИД 31384991 . S2CID 199451378 .
- ^ Jump up to: а б Сингха, Праджжал К.; Рой, Соменат; Дей, Сатьяхари (апрель 2007 г.). «Защитная активность белков андрографолида и арабиногалактана из Andrographis метельчатой Nees. против токсичности, вызванной этанолом, у мышей» . Журнал этнофармакологии . 111 (1): 13–21. дои : 10.1016/j.jep.2006.10.026 . ПМИД 17127022 .
- ^ Финчер, Великобритания; Стоун, бакалавр; Кларк, А.Э. (1 июня 1983 г.). «Арабиногалактан-белки: структура, биосинтез и функции» . Ежегодный обзор физиологии растений . 34 (1): 47–70. дои : 10.1146/annurev.pp.34.060183.000403 . ISSN 0066-4294 .
- ^ Ма, Инсюань; Цзэн, Вэй; Бачич, Антоний; Джонсон, Ким (2018). «AGP сквозь время и пространство». Ежегодные обзоры растений онлайн . Том. 3. С. 767–804. дои : 10.1002/9781119312994.apr0608 . ISBN 9781119312994 . ISSN 2639-3832 . S2CID 104384164 .
- ^ Нгема-Она, Эрик; Викре-Жибуэн, Майте; Каннезан, Марк-Антуан; Дриуич, Азеддин (август 2013 г.). «Белки арабиногалактана во взаимодействии корней и микробов» . Тенденции в науке о растениях . 18 (8): 440–449. doi : 10.1016/j.tplants.2013.03.006 . ISSN 1878-4372 . ПМИД 23623239 . S2CID 8085638 .
- ^ Фильмус, Хорхе; Капурро, Мариана; Раст, Джонатан (2008). «Глипиканы» . Геномная биология . 9 (5): 224. doi : 10.1186/gb-2008-9-5-224 . ISSN 1465-6906 . ПМЦ 2441458 . ПМИД 18505598 .
- ^ Шефер, Лилиана; Шефер, Роланд М. (10 июня 2009 г.). «Протеогликаны: от структурных соединений к сигнальным молекулам» . Исследования клеток и тканей . 339 (1): 237–246. дои : 10.1007/s00441-009-0821-y . ISSN 0302-766X . ПМИД 19513755 . S2CID 20358779 .
- ^ Тан, Ли; Шоуолтер, Аллан М.; Эгелунд, Джек; Эрнандес-Санчес, Арианна; Доблин, Моника С.; Бачич, Энтони (2012). «Арабиногалактан-белки и проблемы исследования этих загадочных протеогликанов поверхности растительных клеток» . Границы в науке о растениях . 3 : 140. дои : 10.3389/fpls.2012.00140 . ISSN 1664-462X . ПМК 3384089 . ПМИД 22754559 .
- ^ Китадзава, Киминари; Трифона, Феодора; Ёшими, Ёсихиса; Хаяси, Ёсихиро; Каваучи, Сусуму; Антонов, Людмил; Танака, Хироши; Такахаси, Такаши; Канеко, Сатоши (март 2013 г.). «Реагент β-галактозил Ярива связывается с β-1,3-галактаном белков арабиногалактана» . Физиология растений . 161 (3): 1117–1126. дои : 10.1104/стр.112.211722 . ISSN 0032-0889 . ПМЦ 3585584 . ПМИД 23296690 .
- ^ Ярив, Дж; Раппорт, ММ; Граф, Л. (1 ноября 1962 г.). «Взаимодействие гликозидов и сахаридов с антителами к соответствующим фенилазогликозидам» . Биохимический журнал . 85 (2): 383–388. дои : 10.1042/bj0850383 . ISSN 0006-2936 . ПМЦ 1243744 . ПМИД 14002491 .
- ^ Тан, Х.-С. (07.07.2006). «Роль белков арабиногалактана, связывающихся с реагентами Yariv, в инициировании, судьбе развития клеток и поддержании эмбриогенеза микроспор у Brassica napus L. сорта Topas» . Журнал экспериментальной ботаники . 57 (11): 2639–2650. дои : 10.1093/jxb/erl027 . ISSN 0022-0957 . ПМИД 16829548 .
- ^ Уиллатс, Уильям Г.Т.; Нокс, Дж. Пол (июнь 1996 г.). «Роль арабиногалактановых белков в размножении растительных клеток: данные исследований взаимодействия бета-глюкозилового реагента Ярива с проростками Arabidopsis thaliana» . Заводской журнал . 9 (6): 919–925. дои : 10.1046/j.1365-313x.1996.9060919.x . ISSN 0960-7412 . ПМИД 8696368 .
- ^ Чепмен, Одри; Блервак, Анн-Софи; Вассер, Жак; Гильберт, Жан-Луи (10 августа 2000 г.). «Арабиногалактан-белки в соматическом эмбриогенезе Cichorium: влияние β-глюкозил-реагента Ярива и локализация эпитопа во время развития эмбриона» . Планта . 211 (3): 305–314. Бибкод : 2000Plant.211..305C . дои : 10.1007/s004250000299 . ISSN 0032-0935 . ПМИД 10987548 . S2CID 23116408 .
- ^ Загорчев Л; Стойнева Р.; Оджакова, М (2013). «Изменения в белках арабиногалактана во время соматического эмбриогенеза. В суспензии культур In vitro Dactylis glomerata L». (PDF) . Болгарский журнал сельскохозяйственных наук . 17 (2): 35–38. ISSN 1310-0351 .
- ^ Рупрехт, Колин; Бартецко, Макс П.; Сенф, Дебора; Даллабернадина, Пьетро; Боос, Ирен; Андерсен, Матиас КФ; Котаке, Тошихиса; Нокс, Дж. Пол; Хан, Майкл Г. (ноябрь 2017 г.). «Синтетический гликановый микрочип позволяет картировать эпитопы антител, направленных на гликан клеточной стенки растений» . Физиология растений . 175 (3): 1094–1104. дои : 10.1104/стр.17.00737 . ISSN 0032-0889 . ПМК 5664464 . ПМИД 28924016 .
- ^ Зайферт, Георг Дж.; Робертс, Кейт (июнь 2007 г.). «Биология белков арабиногалактана» . Ежегодный обзор биологии растений . 58 (1): 137–161. doi : 10.1146/annurev.arplant.58.032806.103801 . ISSN 1543-5008 . ПМИД 17201686 .
- ^ ван Хенгель, Арджон Дж.; Робертс, Кейт (октябрь 2003 г.). «AtAGP30, белок арабиногалактан в клеточных стенках первичного корня, играет роль в регенерации корня и прорастании семян» . Заводской журнал . 36 (2): 256–270. дои : 10.1046/j.1365-313x.2003.01874.x . ISSN 0960-7412 . ПМИД 14535889 .
- ^ КОИМБРА, Сильвия; КОСТА, Марио; МЕНДЕС, Марта Аделина; ПЕРЕЙРА, Ана Марта; ПИНТО, Жуан; ПЕРЕЙРА, Луис Густаво (17 февраля 2010 г.). «Раннее поколение шкива Arabidopsis в двойном нулевом мутанте по генам белка арабиногалактана AGP6 и AGP11» . Размножение половых растений . 23 (3): 199–205. дои : 10.1007/s00497-010-0136-x . ISSN 0934-0882 . ПМИД 20162305 . S2CID 32823162 .
- ^ Сузуки, Тошия; Нарцисс, Джоан; Цзэн, Вэй; ван де Мин, Эллисон; Ясутоми, Масаюки; Такемура, Сюнсукэ; Молния, Эдвин Р.; Доблин, Моника С.; Бачич, Энтони (9 ноября 2016 г.). «KNS4 / UPEX1: арабиногалактан β-(1,3)-галактозилтрансфераза типа II, необходимая для развития экзины пыльцы» . Физиология растений . 173 (1): 183–205. дои : 10.1104/стр.16.01385 . ISSN 0032-0889 . ПМК 5210738 . ПМИД 27837085 .
- ^ Лэмпорт, Дерек Т.А.; Варнаи, Питер (январь 2013 г.). «Периплазматические арабиногалактановые гликопротеины действуют как кальциевый конденсатор, регулирующий рост и развитие растений» . Новый фитолог . 197 (1): 58–64. дои : 10.1111/nph.12005 . ISSN 0028-646X . ПМИД 23106282 .
- ^ Лэмпорт, Дерек Т.А.; Тан, Ли; Держись, Майкл; Келишевский, Марсия Дж. (09 февраля 2020 г.). «Филлотаксис открывает новый лист — новая гипотеза» . Международный журнал молекулярных наук . 21 (3): 1145. doi : 10.3390/ijms21031145 . ISSN 1422-0067 . ПМК 7037126 . ПМИД 32050457 .
- ^ Лопес-Эрнандес, Федерико; Трифона, Феодора; Рицца, Анналиса; Ю, Сяолань Л.; Харрис, Мэтью О.Б.; Уэбб, Алекс А.Р.; Котаке, Тошихиса; Дюпри, Поль (октябрь 2020 г.). «Связывание кальция полисахаридами арабиногалактана важно для нормального развития растений» . Растительная клетка . 32 (10): 3346–3369. дои : 10.1105/tpc.20.00027 . ISSN 1040-4651 . ПМЦ 7534474 . ПМИД 32769130 .
- ^ Jump up to: а б Хубер, О.; Сампер, М. (15 сентября 1994 г.). «Водорослевые САМ: изоформы молекулы клеточной адгезии в эмбрионах водоросли Volvox, гомологичные фасциклину I дрозофилы» . Журнал ЭМБО . 13 (18): 4212–4222. дои : 10.1002/j.1460-2075.1994.tb06741.x . ISSN 0261-4189 . ПМЦ 395348 . ПМИД 7925267 .
- ^ Зайферт, Георг Дж. (31 мая 2018 г.). «Увлекательные фасциклины: удивительно широко распространенное семейство белков, которые опосредуют взаимодействие между внешней и клеточной поверхностью» . Международный журнал молекулярных наук . 19 (6): 1628. doi : 10.3390/ijms19061628 . ISSN 1422-0067 . ПМК 6032426 . ПМИД 29857505 .
- ^ Пун, Саймон; Хит, Робин Луиза; Кларк, Эдриен Элизабет (2 августа 2012 г.). «Химерный белок арабиногалактан способствует соматическому эмбриогенезу в культуре хлопковых клеток» . Физиология растений . 160 (2): 684–695. дои : 10.1104/стр.112.203075 . ISSN 0032-0889 . ПМЦ 3461548 . ПМИД 22858635 .
- ^ Тунен, Марсель А.Дж.; Шмидт, Эд Д.Л.; ван Каммен, Аб; де Врис, Сакко К. (26 сентября 1997 г.). «Стимулирующее и ингибирующее действие различных белков арабиногалактана на соматический эмбриогенез Daucus carota L.» . Растение . 203 (2): 188–195. Бибкод : 1997Завод.203..188Т . дои : 10.1007/s004250050181 . ISSN 0032-0935 . S2CID 35053257 .
- ^ Ху, Ин; Цинь, Юань; Чжао, Цзе (06 октября 2006 г.). «Локализация эпитопа белка арабиногалактана и эффекты фенилгликозида Ярива во время развития зиготических эмбрионов Arabidopsis thaliana» . Протоплазма . 229 (1): 21–31. дои : 10.1007/s00709-006-0185-z . ISSN 0033-183X . ПМИД 17019527 . S2CID 9707077 .
- ^ Цинь, Ю. (31 января 2006 г.). «Локализация белков арабиногалактана в яйцеклетках, зиготах и двухклеточных проэмбрионах и влияние реагента -D-глюкозил Ярив на оплодотворение яйцеклеток и деление зигот у Nicotiana tabacum L». Журнал экспериментальной ботаники . 57 (9): 2061–2074. дои : 10.1093/jxb/erj159 . ISSN 0022-0957 . ПМИД 16720612 .
- ^ Штайнмахер, Дуглас А.; Сааре-Сурмински, Катя; Либерей, Рейнхард (19 июня 2012 г.). «Белки арабиногалактана и поверхностная сеть внеклеточного матрикса во время соматического эмбриогенеза персиковой пальмы» . Физиология Плантарум . 146 (3): 336–349. дои : 10.1111/j.1399-3054.2012.01642.x . ISSN 0031-9317 . ПМИД 22574975 .
- ^ Пан, Сяо; Ян, Сяо; Линь, Гуймэй; Цзоу, Ру; Чен, Хубин; Самадж, Джозеф; Сюй, Чуньсян (24 мая 2011 г.). «Ультраструктурные изменения и распределение белков арабиногалактана во время соматического эмбриогенеза банана (Musa spp. AAA сорт 'Yueyoukang 1')» . Физиология Плантарума . 142 (4): 372–389. дои : 10.1111/j.1399–3054.2011.01478.x . ISSN 0031-9317 . ПМИД 21496030 .
- ^ Духов, Стефани; Дальке, Рената И.; Геске, Томас; Блашек, Вольфганг; Классен, Биргит (ноябрь 2016 г.). «Арабиногалактан-белки стимулируют соматический эмбриогенез и размножение растений Pelargonium sidoides» . Углеводные полимеры . 152 : 149–155. дои : 10.1016/j.carbpol.2016.07.015 . ISSN 0144-8617 . ПМИД 27516259 .
- ^ Халлманн, А.; Кирк, Д.Л. (декабрь 2000 г.). «Регулируемый в процессе развития гликопротеин ЕСМ ISG играет важную роль в организации ЕСМ и ориентации клеток вольвокса» . Журнал клеточной науки . 113 (24): 4605–4617. дои : 10.1242/jcs.113.24.4605 . ISSN 0021-9533 . ПМИД 11082052 .
- ^ Перейра, Ана Марта; Лопес, Ана Люсия; Коимбра, Сильвия (14 июля 2016 г.). «JAGGER, AGP, необходимый для стойкой синергидной дегенерации и блокады политубея у арабидопсиса» . Сигнализация и поведение растений . 11 (8): e1209616. Бибкод : 2016PlSiB..11E9616P . дои : 10.1080/15592324.2016.1209616 . ISSN 1559-2324 . ПМК 5022411 . ПМИД 27413888 .
- ^ Левитин, Белла; Рихтер, Джганит; Маркович, Инбал; Зик, Мория (ноябрь 2008 г.). «Белки арабиногалактана 6 и 11 необходимы для функционирования тычинок и пыльцы арабидопсиса» . Заводской журнал . 56 (3): 351–363. дои : 10.1111/j.1365-313x.2008.03607.x . ISSN 0960-7412 . ПМИД 18644001 .
- ^ Коимбра, С.; Коста, М.; Джонс, Б.; Мендес, Массачусетс; Перейра, LG (11 мая 2009 г.). «У арабидопсиса с нулевыми мутантами agp6 agp11 нарушено развитие пыльцевого зерна» . Журнал экспериментальной ботаники . 60 (11): 3133–3142. дои : 10.1093/jxb/erp148 . ISSN 0022-0957 . ПМК 2718217 . ПМИД 19433479 .
- ^ Акоста-Гарсия, Херардо; Виэль-Кальсада, Жан-Филипп (17 сентября 2004 г.). «Классический белок арабиногалактан необходим для инициации женского гаметогенеза у Arabidopsis» . Растительная клетка . 16 (10): 2614–2628. дои : 10.1105/tpc.104.024588 . ISSN 1040-4651 . ПМК 520959 . ПМИД 15377758 .
- ^ Демеса-Аревало, Эдгар; Виэль-Кальсада, Жан-Филипп (апрель 2013 г.). «Классический белок арабиногалактан AGP18 опосредует селекцию мегаспор у Arabidopsis» . Растительная клетка . 25 (4): 1274–1287. дои : 10.1105/tpc.112.106237 . ISSN 1040-4651 . ПМЦ 3663267 . ПМИД 23572547 .
- ^ Jump up to: а б Ли, Юньцзин; Лю, Дицю; Ту, Лили; Чжан, Сяньлун; Ван, Ли; Чжу, Лунфу; Тан, Цзяфу; Дэн, Фэнлинь (30 декабря 2009 г.). «Подавление экспрессии гена GhAGP4 подавляло инициацию и удлинение хлопкового волокна» . Отчеты о растительных клетках . 29 (2): 193–202. дои : 10.1007/s00299-009-0812-1 . ISSN 0721-7714 . ПМИД 20041253 . S2CID 1341378 .
- ^ Масигучи, Киёси; Асами, Тадао; Сузуки, Ёшихито (23 ноября 2009 г.). «Полногеномная идентификация, исследование структуры и экспрессии, а также сбор мутантов 22 генов ранних нодулиноподобных белков у Arabidopsis» . Бионауки, биотехнологии и биохимия . 73 (11): 2452–2459. дои : 10.1271/bbb.90407 . ISSN 0916-8451 . ПМИД 19897921 . S2CID 27449840 .
- ^ Хоу, Иннань; Го, Синьян; Киприс, Филипп; Чжан, Ин; Блекманн, Андреа; Кай, Ле; Хуан, Цинпей; Ло, Ю; Гу, Хунья (сентябрь 2016 г.). «Материнские ENODL необходимы для приема пыльцевой трубки у арабидопсиса» . Современная биология . 26 (17): 2343–2350. Бибкод : 2016CBio...26.2343H . дои : 10.1016/j.cub.2016.06.053 . ISSN 0960-9822 . ПМК 5522746 . ПМИД 27524487 .
- ^ Лин, Сью; Донг, Хэн; Чжан, Фанг; Цю, Линь; Ван, Фанчжан; Цао, Цзяшу; Хуан, Ли (31 января 2014 г.). «BcMF8, предполагаемый ген, кодирующий белок арабиногалактана, способствует развитию пыльцевых стенок, образованию отверстий и росту пыльцевых трубок у Brassica Campestris» . Анналы ботаники . 113 (5): 777–788. дои : 10.1093/aob/mct315 . ISSN 1095-8290 . ПМЦ 3962243 . ПМИД 24489019 .
- ^ Лин, Сью; Юэ, Сяоянь; Мяо, Инцзин; Ю, Юцзян; Донг, Хэн; Хуан, Ли; Цао, Цзяшу (09 марта 2018 г.). «Отличные функции двух классических белков арабиногалактана BcMF8 и BcMF18 во время развития стенок пыльцы у Brassica Campestris» . Заводской журнал . 94 (1): 60–76. дои : 10.1111/tpj.13842 . ISSN 0960-7412 . ПМИД 29385650 .
- ^ Чунг, Элис Ю ; Ван, Хун; У, Хэнь-мин (август 1995 г.). «Цветочный передающий тканеспецифичный гликопротеин привлекает пыльцевые трубки и стимулирует их рост» . Клетка . 82 (3): 383–393. дои : 10.1016/0092-8674(95)90427-1 . ISSN 0092-8674 . ПМИД 7634328 . S2CID 17604437 .
- ^ Натан Хэнкок, К.; Кент, Лия; МакКлюр, Брюс А. (8 августа 2005 г.). «Стилярный гликопротеин массой 120 кДа необходим для S-специфического отторжения пыльцы у Никотианы» . Заводской журнал . 43 (5): 716–723. дои : 10.1111/j.1365-313x.2005.02490.x . ISSN 0960-7412 . ПМИД 16115068 .
- ^ Тан, Гексин; Лян, Ваньци; Ху, Цзяньпин; Чжан, Дабин (июнь 2012 г.). «MTR1 кодирует секреторный гликопротеин фасциклин, необходимый для мужского репродуктивного развития в рисе» . Развивающая клетка . 22 (6): 1127–1137. дои : 10.1016/j.devcel.2012.04.011 . ISSN 1534-5807 . ПМИД 22698279 .
- ^ Ян, Цзе; Сардар, Харджиндер С.; Макговерн, Кэтлин Р.; Чжан, Ичжу; Шоуолтер, Аллан М. (8 января 2007 г.). «Богатый лизином белок арабиногалактан в арабидопсисе необходим для роста и развития растений, включая деление и размножение клеток» . Заводской журнал . 49 (4): 629–640. дои : 10.1111/j.1365-313x.2006.02985.x . ISSN 0960-7412 . ПМИД 17217456 .
- ^ Тан, Ли; Эберхард, Стефан; Паттатил, Сивакумар; Уордер, Клейтон; Глушка, Джон; Юань, Чуньхуа; Хао, Чжанъин; Чжу, Сян; Авчи, Утку (январь 2013 г.). «Протеогликан клеточной стенки арабидопсиса состоит из пектина и арабиноксилана, ковалентно связанных с белком арабиногалактана» . Растительная клетка . 25 (1): 270–287. дои : 10.1105/tpc.112.107334 . ISSN 1040-4651 . ПМЦ 3584541 . ПМИД 23371948 .
- ^ Джонсон, Ким Л.; Киббл, Натали Эй Джей; Бачич, Антоний; Шульц, Кэролайн Дж. (22 сентября 2011 г.). «Мутант Arabidopsis thaliana, fla1, подобный фасциклину-белку арабиногалактана (FLA), демонстрирует дефекты регенерации побегов» . ПЛОС ОДИН . 6 (9): e25154. Бибкод : 2011PLoSO...625154J . дои : 10.1371/journal.pone.0025154 . ISSN 1932-6203 . ПМЦ 3178619 . ПМИД 21966441 .
- ^ Ши, Хуачжун; Ким, ЁнгСиг; Го, Ян; Стивенсон, Бекки; Чжу, Цзянь-Кан (13 декабря 2002 г.). «Локус SOS5 Arabidopsis кодирует предполагаемый белок адгезии к поверхности клеток и необходим для нормального расширения клеток» . Растительная клетка . 15 (1): 19–32. дои : 10.1105/tpc.007872 . ISSN 1040-4651 . ПМЦ 143448 . ПМИД 12509519 .
- ^ Харпаз-Саад, Смадар; Макфарлейн, Хизер Э.; Сюй, Шулин; Диви, Удай К.; Нападающий, Бронвен; Вестерн, Тамара Л.; Кибер, Джозеф Дж. (10 октября 2011 г.). «Синтез целлюлозы по пути FEI2 RLK/SOS5 и ЦЕЛЛЮЛОЗНО-СИНТАЗА 5 необходим для структуры слизи семенной оболочки арабидопсиса» . Заводской журнал . 68 (6): 941–953. дои : 10.1111/j.1365-313x.2011.04760.x . ISSN 0960-7412 . ПМИД 21883548 .
- ^ Гриффитс, Джонатан С.; Цай, Аллен И-Лун; Сюэ, Хуэй; Войничюк, Кэтэлин; Шола, Крешимир; Зайферт, Георг Дж.; Мэнсфилд, Шон Д.; Хон, Джордж В. (7 мая 2014 г.). «Чрезмерная чувствительность к соли5 опосредует прилипание и организацию слизистой оболочки семени арабидопсиса посредством пектинов» . Физиология растений . 165 (3): 991–1004. дои : 10.1104/стр.114.239400 . ISSN 0032-0889 . ПМК 4081351 . ПМИД 24808103 .
- ^ Гриффитс, Джонатан С.; Крепо, Мари-Жанна; Рале, Мари-Кристин; Зайферт, Георг Дж.; Норт, Хелен М. (29 июля 2016 г.). «Рассеивание прилипания семенной слизи, опосредованное FEI2 и SOS5» . Границы в науке о растениях . 7 : 1073. дои : 10.3389/fpls.2016.01073 . ISSN 1664-462X . ПМЦ 4965450 . ПМИД 27524986 .
- ^ Сюэ, Хуэй; Вейт, Кристиана; Абас, Линди; Трифона, Феодора; Мареш, Дэниел; Рикарди, Мартиниано М.; Эстевес, Хосе Мануэль; Штрассер, Ричард; Зайферт, Георг Дж. (13 июня 2017 г.). «FLA4 Arabidopsis thaliana действует как стабилизированный гликанами растворимый фактор через свой карбокси-проксимальный домен фасциклина 1» . Заводской журнал . 91 (4): 613–630. дои : 10.1111/tpj.13591 . ISSN 0960-7412 . ПМЦ 5575511 . ПМИД 28482115 .
- ^ Ли, Киран Джей Ди; Саката, Ёичи; Мау, Шайо-Лим; Петтолино, Филомена; Бачич, Антоний; Кватрано, Ральф С.; Найт, Селия Д.; Нокс, Дж. Пол (30 сентября 2005 г.). «Белки арабиногалактана необходимы для удлинения апикальных клеток мха Physcomitrella patens» . Растительная клетка . 17 (11): 3051–3065. дои : 10.1105/tpc.105.034413 . ISSN 1040-4651 . ПМК 1276029 . ПМИД 16199618 .
- ^ Киршнер, Томас В.; Нихаус, Маркус; Дебенер, Томас; Шенк, Манфред К.; Херде, Марко (22 сентября 2017 г.). «Эффективное создание мутаций, опосредованных CRISPR/Cas9, в системе трансформации волосатых корней Brassica carinata» . ПЛОС ОДИН . 12 (9): e0185429. Бибкод : 2017PLoSO..1285429K . дои : 10.1371/journal.pone.0185429 . ISSN 1932-6203 . ПМК 5609758 . ПМИД 28937992 .
- ^ Макмиллан, Коллин П.; Мэнсфилд, Шон Д.; Стачурский, Збигнев Х.; Эванс, Роб; Саутертон, Саймон Г. (24 февраля 2010 г.). «Фасциклиноподобные белки арабиногалактана: специализация в области биомеханики стебля и архитектуры клеточной стенки арабидопсиса и эвкалипта» . Заводской журнал . 62 (4): 689–703. дои : 10.1111/j.1365-313x.2010.04181.x . ISSN 0960-7412 . ПМИД 20202165 .
- ^ Мотосэ, Хироясу; Сугияма, Мунетака; Фукуда, Хироо (июнь 2004 г.). «Протеогликан опосредует индуктивное взаимодействие во время развития сосудов растений» . Природа . 429 (6994): 873–878. Бибкод : 2004Natur.429..873M . дои : 10.1038/nature02613 . ISSN 0028-0836 . ПМИД 15215864 . S2CID 4393158 .
- ^ Ван, Хайхай; Цзян, Чунмей; Ван, Куитинг; Ян, Ян; Ян, Лей; Гао, Сяоянь; Чжан, Хунся (26 ноября 2014 г.). «Антисмысловая экспрессия гена фасциклиноподобного белка арабиногалактана FLA6 у Populus ингибирует экспрессию его гомологичных генов и изменяет биомеханику стебля и состав клеточной стенки у трансгенных деревьев» . Журнал экспериментальной ботаники . 66 (5): 1291–1302. дои : 10.1093/jxb/eru479 . ISSN 1460-2431 . ПМЦ 4339592 . ПМИД 25428999 .
- ^ Альберт, Маркус; Беластеги-Макадам, Ксана; Кальденхофф, Ральф (ноябрь 2006 г.). «Атака растительного паразита Cuscuta reflexa индуцирует экспрессию attAGP, белка прикрепления томата-хозяина» . Заводской журнал . 48 (4): 548–556. дои : 10.1111/j.1365-313x.2006.02897.x . ISSN 0960-7412 . ПМИД 17076801 .
- ^ Гаспар, Иоланда Мария; Нам, Джесон; Шульц, Кэролин Джейн; Ли, Лань-Ин; Гилсон, Пол Р.; Гелвин, Стэнтон Б.; Бачич, Энтони (август 2004 г.). «Характеристика мутанта арабидопсиса, богатого лизином арабиногалактан-белка AtAGP17 (крыса1), который приводит к снижению эффективности трансформации агробактерий» . Физиология растений . 135 (4): 2162–2171. дои : 10.1104/стр.104.045542 . ISSN 0032-0889 . ПМК 520787 . ПМИД 15286287 .
- ^ Тан, Ли; Шоуолтер, Аллан М.; Эгелунд, Джек; Эрнандес-Санчес, Арианна; Доблин, Моника С.; Бачич, Энтони (2012). «Арабиногалактан-белки и проблемы исследования этих загадочных протеогликанов поверхности растительных клеток» . Границы в науке о растениях . 3 : 140. дои : 10.3389/fpls.2012.00140 . ISSN 1664-462X . ПМК 3384089 . ПМИД 22754559 .
- ^ Джонсон, Ким Л.; Джонс, Брайан Дж.; Бачич, Антоний; Шульц, Кэролин Дж. (1 декабря 2003 г.). «Фаскиклин-подобные арабиногалактановые белки арабидопсиса. Мультигенное семейство предполагаемых молекул клеточной адгезии» . Физиология растений . 133 (4): 1911–1925. дои : 10.1104/стр.103.031237 . ISSN 0032-0889 . ПМК 300743 . ПМИД 14645732 .
- ^ Jump up to: а б Басу, Дебарати; Тиан, Лу; Ван, Вуда; Боббс, Шауни; Херок, Хейли; Трэверс, Эндрю; Шоуолтер, Аллан М. (21 декабря 2015 г.). «Небольшое мультигенное семейство гидроксипролин-О-галактозилтрансфераз участвует в гликозилировании, росте и развитии арабиногалактан-белка» . Биология растений BMC . 15 : 295. дои : 10.1186/s12870-015-0670-7 . ISSN 1471-2229 . ПМЦ 4687291 . ПМИД 26690932 .
- ^ Ли, Вэньхуа Л.; Лю, Юаньюань; Дуглас, Карл Дж. (01 января 2017 г.). «Роль гликозилтрансфераз в формировании примексина пыльцевой стенки и формировании рисунка экзины» . Физиология растений . 173 (1): 167–182. дои : 10.1104/стр.16.00471 . ISSN 0032-0889 . ПМК 5210704 . ПМИД 27495941 .
- ^ Кнох, Ева; Дилокпимол, Адифол; Геши, Наоми (2014). «Белки арабиногалактана: фокус на углеводно-активных ферментах» . Границы в науке о растениях . 5 : 198. doi : 10.3389/fpls.2014.00198 . ISSN 1664-462X . ПМК 4052742 . ПМИД 24966860 .