Метаболизм железа у человека
железа у человека представляет собой совокупность химических реакций, которые поддерживают гомеостаз железа у человека на Метаболизм системном и клеточном уровне. Железо одновременно необходимо организму и потенциально токсично. Контроль уровня железа в организме является критически важной частью многих аспектов здоровья и болезней человека. Гематологов особенно интересовал системный метаболизм железа , поскольку железо необходимо для эритроцитов , в которых содержится большая часть железа в организме человека. Понимание метаболизма железа также важно для понимания заболеваний, связанных с перегрузкой железом , таких как наследственный гемохроматоз , и дефицита железа , таких как железодефицитная анемия .
Важность регулирования железа
[ редактировать ]
Железо является важным биоэлементом для большинства форм жизни, от бактерий до млекопитающих . Его важность заключается в его способности опосредовать перенос электронов. В двухвалентном состоянии (Fe 2+ ), железо выступает донором электронов , находясь в трехвалентном состоянии (Fe 3+ ) он действует как акцептор . Таким образом, железо играет жизненно важную роль в катализе ферментативных реакций, связанных с переносом электрона (восстановление и окисление, окислительно-восстановительный процесс ). Белки могут содержать железо в составе различных кофакторов , таких как железо-серные кластеры (Fe-S) и гемовые группы, которые собираются в митохондриях .
Клеточное дыхание
[ редактировать ]Человеческие клетки нуждаются в железе для получения энергии в виде АТФ в многоэтапном процессе, известном как клеточное дыхание, а точнее, в результате окислительного фосфорилирования в митохондриальных кристах . Железо присутствует в железо-серном кластере и гемовых группах белков цепи переноса электронов , которые генерируют протонный градиент , позволяющий АТФ-синтазе синтезировать АТФ ( хемиосмос ).
Гемовые группы входят в состав гемоглобина — белка, содержащегося в эритроцитах и служащего для транспортировки кислорода из легких в другие ткани. Гемовые группы также присутствуют в миоглобине для хранения и диффузии кислорода в мышечных клетках.
Транспорт кислорода
[ редактировать ]Человеческому организму железо необходимо для транспортировки кислорода. Кислород (O 2 ) необходим для функционирования и выживания почти всех типов клеток. Кислород транспортируется из легких в остальную часть тела, связанный с гемовой группой гемоглобина в эритроцитах. В мышечных клетках железо связывает кислород с миоглобином , который регулирует его высвобождение.
Токсичность
[ редактировать ]Железо также потенциально токсично. Его способность отдавать и принимать электроны означает, что он может катализировать превращение перекиси водорода в свободные радикалы . Свободные радикалы могут вызвать повреждение самых разных клеточных структур и в конечном итоге убить клетку. [ 1 ]
Железо, связанное с белками или кофакторами, такими как гем, безопасно. Также в клетке практически нет по-настоящему свободных ионов железа, поскольку они легко образуют комплексы с органическими молекулами. Однако некоторая часть внутриклеточного железа связана с комплексами с низким сродством и называется лабильным железом или «свободным» железом. Железо в таких комплексах может вызвать описанные выше повреждения. [ 2 ]
Чтобы предотвратить такого рода повреждения, все формы жизни, использующие железо, связывают атомы железа с белками . Это связывание позволяет клеткам получать пользу от железа, одновременно ограничивая его способность причинять вред. [ 1 ] [ 3 ] Типичные внутриклеточные концентрации лабильного железа у бактерий составляют 10-20 микромолярных. [ 4 ] хотя в анаэробной среде они могут быть в 10 раз выше, [ 5 ] где свободных радикалов и активных форм кислорода меньше. В клетках млекопитающих внутриклеточные концентрации лабильного железа обычно составляют менее 1 микромоляря, что составляет менее 5 процентов от общего количества клеточного железа. [ 2 ]
Бактериальная защита
[ редактировать ]
В ответ на системную бактериальную инфекцию иммунная система инициирует процесс, известный как « удержание железа ». Если бактерии хотят выжить, им необходимо получать железо из окружающей среды. Бактерии, вызывающие заболевания, делают это разными способами, в том числе высвобождают связывающие железо молекулы, называемые сидерофорами , а затем реабсорбируют их для восстановления железа или удаляют железо из гемоглобина и трансферрина . Чем усерднее бактериям приходится работать, чтобы получить железо, тем большую метаболическую цену им приходится платить. Это означает, что бактерии, лишенные железа, размножаются медленнее. Таким образом, контроль уровня железа является важной защитой от многих бактериальных инфекций. Некоторые виды бактерий разработали стратегии, позволяющие обойти эту защиту. Бактерии, вызывающие туберкулез , могут обитать в макрофагах , которые представляют собой среду, богатую железом, а Borrelia burgdorferi использует марганец вместо железа. Люди с повышенным количеством железа, как, например, при гемохроматозе, более восприимчивы к некоторым бактериальным инфекциям. [ 6 ]
Хотя этот механизм является элегантным ответом на кратковременную бактериальную инфекцию, он может вызвать проблемы, если длится так долго, что организм лишается железа, необходимого для производства эритроцитов. Воспалительные цитокины стимулируют печень вырабатывать белок-регулятор метаболизма железа гепсидин , который снижает доступность железа. Если уровни гепсидина повышаются из-за небактериальных источников воспаления, таких как вирусная инфекция, рак, аутоиммунные заболевания или другие хронические заболевания, то может возникнуть анемия хронического заболевания . В этом случае отказ от железа фактически ухудшает здоровье, препятствуя выработке достаточного количества эритроцитов, содержащих гемоглобин. [ 3 ]
Магазины железа для тела
[ редактировать ]
Большинство хорошо питающихся людей в промышленно развитых странах имеют в организме от 4 до 5 граммов железа (~38 мг железа/кг массы тела у женщин и ~50 мг железа/кг массы тела у мужчин). [ 7 ] Из них около 2,5 г содержится в гемоглобине, необходимом для переноса кислорода по крови (около 0,5 мг железа на мл крови), [ 8 ] а большая часть остального (приблизительно 2 грамма у взрослых мужчин и несколько меньше у женщин детородного возраста) содержится в комплексах ферритина , присутствующих во всех клетках, но наиболее часто встречающихся в костном мозге, печени и селезенке . Запасы ферритина в печени являются основным физиологическим источником резервного железа в организме. Запасы железа в промышленно развитых странах, как правило, ниже у детей и женщин детородного возраста, чем у мужчин и пожилых людей. Женщины, которым приходится использовать свои запасы железа для компенсации потери железа во время менструации , беременности или лактации, имеют более низкие запасы негемоглобина в организме, которые могут составлять 500 мг или даже меньше.
Из общего содержания железа в организме около 400 мг приходится на клеточные белки, которые используют железо для важных клеточных процессов, таких как хранение кислорода (миоглобин) или выполнение окислительно-восстановительных реакций, производящих энергию ( цитохромы ). Относительно небольшое количество (3–4 мг) циркулирует в плазме , связанное с трансферрином. [ 9 ] Из-за своей токсичности свободное растворимое железо сохраняется в организме в низкой концентрации.
Дефицит железа в первую очередь влияет на запасы железа в организме, и считается, что истощение этих запасов протекает относительно бессимптомно, хотя некоторые неопределенные и неспецифические симптомы с ним связаны . Поскольку железо в первую очередь необходимо для гемоглобина, железодефицитная анемия является основным клиническим проявлением дефицита железа. Люди с дефицитом железа страдают или умирают от повреждения органов задолго до того, как в их клетках закончится железо, необходимое для внутриклеточных процессов, таких как транспорт электронов.
Макрофаги ретикулоэндотелиальной системы накапливают железо в процессе расщепления и переработки гемоглобина из поглощенных эритроцитов. Железо также хранится в виде пигмента, называемого гемосидерином , который представляет собой нечеткое отложение белка и железа, создаваемое макрофагами, где присутствует избыток железа, локально или системно, например, у людей с перегрузкой железом из-за частого разрушения клеток крови и необходимые переливания крови, которых требует их состояние. Если корригировать системную перегрузку железом, со временем гемосидерин медленно резорбируется макрофагами.
Механизмы регуляции железа
[ редактировать ]
Гомеостаз железа у человека регулируется на двух разных уровнях. Системный уровень железа уравновешивается контролируемым поглощением пищевого железа энтероцитами , клетками, выстилающими внутреннюю часть кишечника , и неконтролируемой потерей железа в результате отслаивания эпителия, пота, травм и кровопотери. Кроме того, системное железо постоянно перерабатывается. Уровни клеточного железа по-разному контролируются разными типами клеток из-за экспрессии определенных регуляторных и транспортных белков железа.
Системная регуляция железа
[ редактировать ]
Поглощение железа с пищей
[ редактировать ]Всасывание пищевого железа представляет собой переменный и динамичный процесс. Количество абсорбированного железа по сравнению с количеством потребляемого железа обычно невелико, но может варьироваться от 5% до 35% в зависимости от обстоятельств и типа железа. Эффективность поглощения железа варьируется в зависимости от источника. Как правило, лучше всего усваиваемые формы железа поступают из продуктов животного происхождения. Усвоение пищевого железа в форме солей железа (как и в большинстве добавок) несколько варьируется в зависимости от потребности организма в железе и обычно составляет от 10% до 20% потребления железа. Всасывание железа из продуктов животного происхождения и некоторых растительных продуктов происходит в форме гемового железа и является более эффективным, позволяя усваивать от 15% до 35% принятого количества. Гемовое железо у животных поступает из крови и гемсодержащих белков мяса и митохондрий, тогда как у растений гемовое железо присутствует в митохондриях всех клеток, использующих кислород для дыхания.
всасывается в двенадцатиперстной кишке энтероцитами Как и большинство минеральных питательных веществ, большая часть железа, всасываемого из переваренной пищи или добавок , слизистой оболочки двенадцатиперстной кишки. Эти клетки имеют специальные молекулы, которые позволяют им переносить железо в организм. Чтобы абсорбироваться, пищевое железо может усваиваться как часть белка, такого как гем-белок, или железо должно находиться в составе двухвалентного железа. 2+ форма. энтероцитов Фермент редуктаза железа на щеточной кайме , дуоденальный цитохром B ( Dcytb ), восстанавливает железо Fe. 3+ в Фе 2+ . [ 10 ] Белок, называемый транспортером двухвалентных металлов 1 ( DMT1 ), может транспортировать несколько двухвалентных энтероцита металлов через плазматическую мембрану, а затем транспортировать железо через клеточную мембрану в клетку. Если железо связано с гемом, вместо этого оно транспортируется через апикальную мембрану белком-переносчиком гема 1 (HCP1). [ 11 ] Затем гем катаболизируется микросомальной гемоксигеназой в биливердин , высвобождая Fe. 2+ . [ 12 ]
Эти клетки слизистой оболочки кишечника затем могут либо хранить железо в виде ферритина , что достигается с помощью Fe. 2+ связывание с апоферритином (в этом случае железо покидает организм, когда клетка умирает и выделяется с фекалиями ), или клетка может высвободить его в организм через единственный известный у млекопитающих экспортер железа — ферропортин . Гефестин — ферроксидаза , способная окислять железо. 2+ в Фе 3+ и обнаруживается главным образом в тонком кишечнике, помогает ферропортину переносить железо через базолатеральный конец клеток кишечника. При попадании в кровь Fe 3+ связывает трансферрин и циркулирует в тканях. Напротив, ферропортин посттрансляционно репрессируется гепсидином , пептидным гормоном, состоящим из 25 аминокислот. Организм регулирует уровень железа, регулируя каждый из этих этапов. Например, энтероциты синтезируют больше Dcytb, DMT1 и ферропортина в ответ на железодефицитную анемию. [ 13 ] Всасывание железа из рациона увеличивается в присутствии витамина С и снижается при избытке кальция, цинка или марганца. [ 14 ]
Скорость всасывания железа в организме человека, по-видимому, зависит от множества взаимозависимых факторов, включая общие запасы железа, степень выработки костным мозгом новых эритроцитов, концентрацию гемоглобина в крови и содержание кислорода в крови. кровь. Организм также поглощает меньше железа во время воспаления , чтобы лишить бактерии железа. Недавние открытия показывают, что гепсидиновая регуляция ферропортина ответственна за синдром анемии хронических заболеваний.
Переработка и потеря железа
[ редактировать ]Большая часть железа в организме накапливается и перерабатывается ретикулоэндотелиальной системой, которая разрушает старые эритроциты. железа не существует В отличие от поглощения и рециркуляции железа, физиологического механизма регуляции выведения . Люди теряют небольшое, но постоянное количество из-за кровопотери в желудочно-кишечном тракте, потоотделения и отшелушивания клеток кожи и слизистой оболочки желудочно-кишечного тракта . Общий объем потерь для здоровых людей в развитых странах составляет в среднем 1 мг в день для мужчин и 1,5–2 мг в день для женщин с регулярными менструациями. [ 15 ] Люди с желудочно-кишечными паразитарными инфекциями, чаще встречающимися в развивающихся странах, часто теряют больше. [ 1 ] Те, кто не может достаточно хорошо регулировать всасывание, страдают от перегрузки железом. При этих заболеваниях токсичность железа начинает подавлять способность организма связывать и хранить его. [ 16 ]
Регуляция клеточного железа
[ редактировать ]Импорт железа
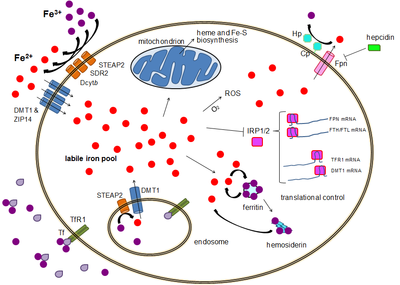
[ редактировать ]Большинство типов клеток поглощают железо в основном посредством рецептор-опосредованного эндоцитоза через рецептор трансферрина 1 (TFR1), рецептор трансферрина 2 (TFR2) и GAPDH . TFR1 имеет в 30 раз более высокое сродство к железу, связанному с трансферрином, чем TFR2, и, таким образом, является основным игроком в этом процессе. [ 17 ] [ 18 ] Многофункциональный гликолитический фермент высшего порядка глицеральдегид-3-фосфатдегидрогеназа (ГАФДГ) также действует как рецептор трансферрина. [ 19 ] [ 20 ] Связанное с трансферрином трехвалентное железо распознается этими рецепторами трансферрина, вызывая конформационные изменения, вызывающие эндоцитоз. Затем железо поступает в цитоплазму из эндосомы через импортер DMT1 после восстановления до двухвалентного состояния редуктазой семейства STEAP. [ 21 ]
Альтернативно, железо может проникать в клетку напрямую через импортеры двухвалентных катионов плазматической мембраны, такие как DMT1 и ZIP14 (Zrt-Irt-подобный белок 14). [ 22 ] Опять же, железо попадает в цитоплазму в двухвалентном состоянии после восстановления во внеклеточном пространстве редуктазой, такой как STEAP2, STEAP3 (в эритроцитах), Dcytb (в энтероцитах) и SDR2. [ 21 ]
Импорт железа в некоторые раковые клетки
[ редактировать ]Железо также может проникать в клетки через CD44 в комплексах, связанных с гиалуроновой кислотой во время эпителиально-мезенхимального перехода (ЕМТ). В этом процессе эпителиальные клетки трансформируются в мезенхимальные клетки с отрывом от базальной мембраны , к которой они обычно прикреплены, открывая путь для вновь дифференцированных подвижных мезенхимальных клеток, чтобы начать миграцию от эпителиального слоя. [ 23 ] [ 24 ]
Хотя ЕМТ играет решающую роль в физиологических процессах, таких как имплантация , когда она позволяет эмбриону проникать в эндометрий, чтобы облегчить прикрепление плаценты , ее нарушение регуляции также может способствовать злокачественных распространению опухолей , позволяя им проникать в окружающие ткани и образовывать отдаленные колонии ( метастазирование ). [ 24 ]
Злокачественные клетки часто демонстрируют повышенную потребность в железе, что способствует их переходу в более инвазивное мезенхимальное состояние. Это железо необходимо для экспрессии мезенхимальных генов, например тех, которые кодируют трансформирующий фактор роста бета (TGF-β), имеющий решающее значение для ЕМТ. Примечательно, что уникальная способность железа катализировать белков и ДНК деметилирование играет жизненно важную роль в этом процессе экспрессии генов. [ 23 ]
Обычные пути поглощения железа, такие как те, которые используют рецептор трансферрина 1 (TfR1), часто оказываются недостаточными для удовлетворения этих повышенных потребностей в железе в раковых клетках. В результате различные цитокины и факторы роста вызывают активацию CD44, поверхностной молекулы, способной усваивать железо, связанное с гиалуронановым комплексом. Этот альтернативный путь, основанный на CD44-опосредованном эндоцитозе, становится доминирующим механизмом поглощения железа по сравнению с традиционным TfR1-зависимым путем. [ 23 ] [ 24 ]
Пул лабильного железа
[ редактировать ]В цитоплазме двухвалентное железо находится в растворимом, хелатируемом состоянии, которое составляет пул лабильного железа (~0,001 мМ). [ 25 ] Считается, что в этом пуле железо связано с соединениями малой массы, такими как пептиды, карбоксилаты и фосфаты, хотя некоторые из них могут находиться в свободной гидратированной форме ( аква-ионы ). [ 25 ] Альтернативно, ионы железа могут быть связаны со специализированными белками, известными как металлочапероны . [ 26 ] В частности, поли-r(C)-связывающие белки PCBP1 и PCBP2, по-видимому, опосредуют перенос свободного железа в ферритин (для хранения) и негемовые ферменты железа (для использования в катализе). [ 22 ] [ 27 ] Пул лабильного железа потенциально токсичен из-за способности железа генерировать активные формы кислорода. Железо из этого пула может поглощаться митохондриями через митоферрин для синтеза кластеров Fe-S и гемовых групп. [ 21 ]
Железный бассейн для хранения
[ редактировать ]Железо может храниться в ферритине в виде трехвалентного железа благодаря ферроксидазной активности тяжелой цепи ферритина. [ 28 ] Дисфункциональный ферритин может накапливаться в виде гемосидерина , что может быть проблематичным в случае перегрузки железом. [ 29 ] Пул запасенного железа ферритина намного больше, чем пул лабильного железа, и его концентрация варьируется от 0,7 до 3,6 мМ. [ 25 ]
Экспорт железа
[ редактировать ]Экспорт железа происходит в различных типах клеток, включая нейроны , эритроциты, макрофаги и энтероциты. Последние два особенно важны, поскольку от них зависит системный уровень железа. Существует только один известный экспортер железа — ферропортин . [ 30 ] Он транспортирует двухвалентное железо из клетки, обычно при помощи церулоплазмина и/или гефестина (в основном в энтероцитах), которые окисляют железо до трехвалентного состояния, чтобы оно могло связывать ферритин во внеклеточной среде. [ 21 ] Гепсидин вызывает интернализацию ферропортина, снижая экспорт железа. Кроме того, гепсидин, по-видимому, подавляет как TFR1, так и DMT1 посредством неизвестного механизма. [ 31 ] Еще одним игроком, помогающим ферропортину осуществлять экспорт железа из клеток, является GAPDH. [ 32 ] Специфическая посттрансляционно модифицированная изоформа GAPDH рекрутируется на поверхность клеток, нагруженных железом, где она рекрутирует апо-трансферрин в непосредственной близости от ферропортина, чтобы быстро хелатировать экструдированное железо. [ 33 ]

- Путь трансцитоза (показан в правом верхнем сегменте изображения), где комплекс «Fe 3+ -трансферрин-трансферриновый рецептор 1 (TfR1)» подвергается эндоцитозу и экзоцитозу от люминального полюса к церебральному внеклеточному матриксу (ECM) и интерстициальной жидкости .
- Облегченный путь транспортера, при котором эндотелиальные клетки интернализуют комплекс «Fe 3+ -трансферрин-трансферриновый рецептор 1 (TfR1)» в эндосоме, снижает содержание железа в железе 3+ ион в двухвалентное железо 2+ ион ферментом STEAP3, а затем Fe 2+ ион пересекает эндосомальную мембрану благодаря DMT1. Фе 2+ затем экспортируется во внеклеточный матрикс (ECM) и интерстициальную жидкость через ферропортин в сочетании с церулоплазмином.
Экспрессия гепсидина, который происходит только в определенных типах клеток, таких как гепатоциты , жестко контролируется на уровне транскрипции и представляет собой связь между клеточным и системным гомеостазом железа благодаря роли гепсидина как «привратника» высвобождения железа из энтероцитов в остальные клетки. тела. [ 21 ] Эритробласты вырабатывают эритроферрон — гормон, который ингибирует гепсидин и тем самым увеличивает доступность железа, необходимого для синтеза гемоглобина. [ 35 ]
Трансляционный контроль клеточного железа
[ редактировать ]Хотя некоторый контроль существует на уровне транскрипции, регуляция уровня клеточного железа в конечном итоге контролируется на уровне трансляции с помощью железо-чувствительных белков, связывающих элементы IRP1 и особенно IRP2. [ 36 ] Когда уровень железа низкий, эти белки способны связываться с железо-чувствительными элементами (IRE). IRE представляют собой структуры «стебель-петля» в нетранслируемых областях (UTR) мРНК. [ 21 ]
И ферритин, и ферропортин содержат IRE в своих 5'-UTR, так что при дефиците железа их трансляция подавляется с помощью IRP2, предотвращая ненужный синтез запасного белка и вредный экспорт железа. Напротив, TFR1 и некоторые варианты DMT1 содержат 3'-UTR IRE, которые связывают IRP2 при дефиците железа, стабилизируя мРНК, что гарантирует синтез импортеров железа. [ 21 ]
Патология
[ редактировать ]Дефицит железа
[ редактировать ]
Функциональный или фактический дефицит железа может возникнуть по разным причинам. Эти причины можно сгруппировать в несколько категорий:
- Повышенная потребность в железе, которую диета не может удовлетворить.
- Повышенная потеря железа (обычно за счет кровопотери).
- Дефицит питания. Это может произойти из-за недостатка железа в рационе или употребления продуктов, которые препятствуют всасыванию железа. Наблюдалось ингибирование всасывания, вызванное фитатами в отрубях . [ 37 ] кальций из добавок или молочных продуктов, [ 38 ] и дубильные вещества из чая, [ 39 ] хотя во всех трех исследованиях эффект был небольшим, и авторы цитируемых исследований, касающихся отрубей и чая, отмечают, что эффект, вероятно, будет иметь заметное влияние только в том случае, если большая часть железа будет получена из растительных источников.
- Лекарства, снижающие кислотность. Лекарства, снижающие кислотность, снижают всасывание пищевого железа. Эти лекарства обычно используются при гастрите, рефлюксной болезни и язвах. Ингибиторы протонной помпы (ИПП), антигистаминные препараты H2 и антациды снижают метаболизм железа. [ 40 ]
- Повреждение слизистой оболочки кишечника. Примеры причин такого рода повреждений включают хирургическое вмешательство на двенадцатиперстной кишке или такие заболевания, как болезнь Крона или целиакия , которые значительно уменьшают площадь поверхности, доступную для абсорбции. Инфекции Helicobacter pylori также снижают доступность железа. [ 41 ]
- Воспаление, приводящее к индуцированному гепсидином ограничению высвобождения железа из энтероцитов (см. выше).
- Это также частое явление у беременных женщин и у растущих подростков из-за неправильного питания.
- Острая кровопотеря или острый цирроз печени приводят к недостатку трансферрина, что приводит к выделению железа из организма.
Железная перегрузка
[ редактировать ]Организм способен существенно уменьшить количество железа, которое оно поглощает через слизистую оболочку. Похоже, что полностью остановить процесс транспортировки железа не удастся. Кроме того, в ситуациях, когда избыток железа повреждает саму слизистую оболочку кишечника (например, когда дети едят большое количество таблеток железа, произведенных для потребления взрослыми), еще больше железа может попасть в кровоток и вызвать потенциально смертельный синдром перегрузки железом. Большое количество свободного железа в кровообращении приведет к повреждению критически важных клеток печени, сердца и других метаболически активных органов.
Токсичность железа возникает, когда количество циркулирующего железа превышает количество трансферрина, доступного для его связывания, но организм может энергично регулировать усвоение железа. Таким образом, токсичность железа при приеме внутрь обычно является результатом чрезвычайных обстоятельств, таких как чрезмерное потребление таблеток железа. [1] [ 42 ] а не изменения в диете . Острая токсичность, вызванная приемом железа, вызывает, помимо других проблем, серьезное повреждение слизистой оболочки желудочно-кишечного тракта.
Избыток железа связан с более высокими показателями заболеваемости и смертности. Например, пациенты с раком молочной железы с низкой экспрессией ферропортина (что приводит к более высоким концентрациям внутриклеточного железа) в среднем выживают в течение более короткого периода времени, тогда как высокая экспрессия ферропортина предсказывает 90% 10-летнюю выживаемость у пациентов с раком молочной железы. [ 43 ] Аналогичным образом, генетические вариации генов-переносчиков железа, которые, как известно, повышают уровень железа в сыворотке, также сокращают продолжительность жизни и среднее количество лет, проведенных в добром здравии. [ 44 ] Было высказано предположение, что мутации, которые увеличивают абсорбцию железа, например, ответственные за гемохроматоз (см. Ниже), были выбраны во времена неолита , поскольку они обеспечивали селективное преимущество против железодефицитной анемии. [ 45 ] Увеличение системного уровня железа становится патологическим в пожилом возрасте, что подтверждает представление о том, что антагонистическая плейотропия или «гиперфункция» приводит к старению человека. [ 44 ]
Хроническая токсичность железа обычно является результатом более хронических синдромов перегрузки железом, связанных с генетическими заболеваниями, повторными переливаниями крови или другими причинами. В таких случаях запасы железа у взрослого могут достигать 50 граммов (в 10 раз больше нормального общего количества железа в организме) и более. Наиболее распространенными заболеваниями, связанными с перегрузкой железом, являются наследственный гемохроматоз (HH), вызванный мутациями в гене HFE , и более тяжелое заболевание ювенильный гемохроматоз (JH), вызванное мутациями либо гемоювелина ( HJV ), либо ювенильного гемохроматоза (JH ). [ 46 ] или гепсидин ( HAMP ). Точные механизмы большинства различных форм гемохроматоза взрослых, которые составляют большую часть генетических нарушений перегрузки железом, остаются невыясненными. Итак, хотя исследователи смогли идентифицировать генетические мутации, вызывающие несколько вариантов гемохроматоза у взрослых, теперь они должны обратить свое внимание на нормальное функционирование этих мутировавших генов.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Конрад М.Е., Умбрейт Дж.Н. (апрель 2000 г.). «Нарушения обмена железа». Медицинский журнал Новой Англии . 342 (17): 1293–4. дои : 10.1056/NEJM200004273421716 . ПМИД 10787338 .
- ^ Jump up to: а б Кахлон О., Кабанчик З.И. (2002). «Лабильный пул железа: характеристика, измерение и участие в клеточных процессах». Свободнорадикальная биология и медицина . 33 (8): 1037–1046. дои : 10.1016/s0891-5849(02)01006-7 . ПМИД 12374615 .
- ^ Jump up to: а б Эндрюс, Северная Каролина (декабрь 1999 г.). «Нарушения обмена железа». Медицинский журнал Новой Англии . 341 (26): 1986–95. дои : 10.1056/NEJM199912233412607 . ПМИД 10607817 .
- ^ Ян Ю, Уэйт-Кьюсик Дж. Г., Куппусами П., Юсеф А. Е. (январь 2013 г.). «Внутриклеточное свободное железо и его потенциальная роль в инактивации Escherichia coli, вызванной сверхвысоким давлением» . Прикладная и экологическая микробиология . 79 (2): 722–724. Бибкод : 2013ApEnM..79..722Y . дои : 10.1128/aem.02202-12 . ПМЦ 3553779 . ПМИД 23124235 .
- ^ Ямамото Ю, Фукуи К, Коджин Н, Охья Х, Кимура К, Камио Ю (2004). «Регуляция внутриклеточного пула свободного железа с помощью Dpr обеспечивает толерантность к кислороду Streptococcus mutans» . Журнал бактериологии . 186 (18): 5997–6002. дои : 10.1128/jb.186.18.5997-6002.2004 . ПМЦ 515136 . ПМИД 15342568 .
- ^ Ганц Т. (август 2003 г.). «Гепцидин — ключевой регулятор обмена железа и медиатор анемии воспаления» . Кровь . 102 (3): 783–8. дои : 10.1182/кровь-2003-03-0672 . ПМИД 12663437 .
- ^ Гроппер, Сарин С.; Смит, Джек Л. (2013). Продвинутое питание и обмен веществ человека (6-е изд.). Бельмонт, Калифорния: Уодсворт. п. 481. ИСБН 978-1133104056 .
- ^ Трусвелл, А. Стюарт (15 июля 2010 г.). Азбука питания . Джон Уайли и сыновья. п. 52. ИСБН 9781444314229 .
- ^ Камашелла С., Шриер С.Л. (07 ноября 2011 г.). «Регуляция баланса железа» . До настоящего времени . Проверено 11 марта 2012 г.
- ^ Маккай А.Т., Барроу Д., Латунде-Дада Г.О., Рольфс А., Сагер Г., Мудали Е., Мудали М., Ричардсон С., Барлоу Д., Бомфорд А., Питерс Т.Дж., Раджа К.Б., Ширали С., Хедигер М.А., Фарзане Ф., Симпсон Р.Дж. ( март 2001 г.). «Железорегулируемая редуктаза железа, связанная с усвоением пищевого железа» . Наука . 291 (5509): 1755–9. Бибкод : 2001Sci...291.1755M . дои : 10.1126/science.1057206 . ПМИД 11230685 . S2CID 44351106 .
- ^ Руо, Трейси А. (9 сентября 2005 г.). «Обнаружен кишечный переносчик гема» . Клетка . 122 (5): 649–651. дои : 10.1016/j.cell.2005.08.027 . ISSN 0092-8674 . ПМИД 16143096 . S2CID 9180328 .
- ^ Пшибышевская, Юстина; Жекановская, Ева (2014). «Роль гепсидина, ферропортина, белка HCP1 и DMT1 в всасывании железа в пищеварительном тракте человека» . Обзор гастроэнтерологии . 4 (4): 208–213. дои : 10.5114/стр.2014.45102 . ПМК 4178046 . ПМИД 25276251 .
- ^ Флеминг Р.Э., Бэкон Б.Р. (апрель 2005 г.). «Оркестровка гомеостаза железа». Медицинский журнал Новой Англии . 352 (17): 1741–4. дои : 10.1056/NEJMp048363 . ПМИД 15858181 .
- ^ Аббаспур, Назанин (февраль 2014 г.). «Обзор железа и его значения для здоровья человека» . J Res Med Sci . 19 (2): 164–174. ПМЦ 3999603 . ПМИД 24778671 .
- ^ Джанет, Р. Хант (июнь 2009 г.). «Выведение железа из организма здоровыми мужчинами и женщинами» . Американский журнал клинического питания . 89 (6): 1792–1798. дои : 10.3945/ajcn.2009.27439 . ПМИД 19386738 .
- ^ Шриер С.Л., Бэкон Б.Р. (07 ноября 2011 г.). «Синдромы перегрузки железом, кроме наследственного гемохроматоза» . До настоящего времени . Проверено 11 марта 2012 г.
- ^ Кавабата Х., Жермен Р.С., Вуонг П.Т., Накамаки Т., Саид Дж.В., Кёффлер Х.П. (июнь 2000 г.). «Рецептор трансферрина 2-альфа поддерживает рост клеток как в культивируемых клетках, хелатированных железом, так и in vivo» . Журнал биологической химии . 275 (22): 16618–25. дои : 10.1074/jbc.M908846199 . ПМИД 10748106 .
- ^ Вест А.П., Беннетт М.Дж., Селлерс В.М., Эндрюс Н.С., Эннс Калифорния, Бьоркман П.Дж. (декабрь 2000 г.). «Сравнение взаимодействий рецептора трансферрина и рецептора трансферрина 2 с трансферрином и наследственным белком гемохроматоза HFE» . Журнал биологической химии . 275 (49): 38135–8. дои : 10.1074/jbc.C000664200 . ПМИД 11027676 .
- ^ Кумар С., Шеоканд Н., Мхадешвар М.А., Радже С.И., Радже М. (январь 2012 г.). «Характеристика глицеральдегид-3-фосфатдегидрогеназы как нового рецептора трансферрина». Международный журнал биохимии и клеточной биологии . 44 (1): 189–99. дои : 10.1016/j.biocel.2011.10.016 . ПМИД 22062951 .
- ^ Шеоканд Н., Кумар С., Малхотра Х., Тиллу В., Радже С.И., Радже М. (июнь 2013 г.). «Секретируемая глицеральдегие-3-фосфат [ sic ] дегидрогеназа представляет собой многофункциональный аутокринный рецептор трансферрина для усвоения клеточного железа». Biochimica et Biophysical Acta (BBA) – Общие предметы . 1830 (6): 3816–27. дои : 10.1016/j.bbagen.2013.03.019 . ПМИД 23541988 .
- ^ Jump up to: а б с д и ж г Хентце М.В., Мукенталер М.Ю., Гали Б., Камашелла С. (июль 2010 г.). «Двое в танго: регуляция обмена железа у млекопитающих» . Клетка . 142 (1): 24–38. дои : 10.1016/j.cell.2010.06.028 . ПМИД 20603012 . S2CID 23971474 .
- ^ Jump up to: а б Лейн, DJR; Мерло, AM; Хуанг, ML-H.; Бэ, Д.-Х.; Янссон, П.Дж.; Сахни, С.; Калиновский, Д.С.; Ричардсон, доктор медицинских наук (май 2015 г.). «Поглощение, транспортировка и метаболизм клеточного железа: ключевые молекулы и механизмы, а также их роль в заболеваниях». Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1853 (5): 1130–1144. дои : 10.1016/j.bbamcr.2015.01.021 . ПМИД 25661197 .
- ^ Jump up to: а б с Мюллер, Себастьян; Синдикубвабо, Фабьен; Каньеке, Татьяна; Лафон, Энн; Версини, Антуан; Ломбард, Беранжер; Лоу, Дамарис; Ву, Тинг-Ди; Жинестье, Кристоф; Шараф-Жофре, Эммануэль; Дюран, Аделина; Валло, Селин; Бауланд, Сильвен; Слуга Николас; Родригес, Рафаэль (1 октября 2020 г.). «CD44 регулирует эпигенетическую пластичность, опосредуя эндоцитоз железа» . Природная химия . 12 (10): 929–938. Бибкод : 2020НатЧ..12..929М . дои : 10.1038/s41557-020-0513-5 . ISSN 1755-4330 . ПМЦ 7612580 . ПМИД 32747755 .
- ^ Jump up to: а б с Каллури, Рагху; Вайнберг, Роберт А. (1 июня 2009 г.). «Основы эпителиально-мезенхимального перехода» . Журнал клинических исследований . 119 (6): 1420–1428. дои : 10.1172/JCI39104 . ISSN 0021-9738 . ПМК 2689101 . ПМИД 19487818 .
- ^ Jump up to: а б с Иегуда С., Мостофский Д.И., ред. (2010). Дефицит железа и перегрузка: от фундаментальной биологии к клинической медицине . Питание и здоровье. Нью-Йорк, штат Нью-Йорк: Humana Press . п. 230. дои : 10.1007/978-1-59745-462-9 . ISBN 9781934115220 .
- ^ Финни Лос-Анджелес, O'Halloran TV (май 2003 г.). «Видование переходных металлов в клетке: идеи химии рецепторов ионов металлов». Наука . 300 (5621): 931–6. Бибкод : 2003Sci...300..931F . дои : 10.1126/science.1085049 . ПМИД 12738850 . S2CID 14863354 .
- ^ Филпотт, Кэролайн С.; Рю, Мун-Сун (22 июля 2014 г.). «Специальная доставка: распределение железа в цитозоле клеток млекопитающих» . Границы в фармакологии . 5 : 173. дои : 10.3389/fphar.2014.00173 . ПМК 4106451 . ПМИД 25101000 .
- ^ Арозио П., Леви С. (август 2002 г.). «Ферритин, гомеостаз железа и окислительное повреждение». Свободно-радикальная биология и медицина . 33 (4): 457–63. дои : 10.1016/s0891-5849(02)00842-0 . ПМИД 12160928 .
- ^ Руо Т.А., Куперман С. (сентябрь 2006 г.). «Обмен железа в мозгу». Семинары по детской неврологии . 13 (3): 142–8. дои : 10.1016/j.spen.2006.08.002 . ПМИД 17101452 .
- ^ Ганц Т. (март 2005 г.). «Клеточное железо: ферропортин – единственный выход» . Клеточный метаболизм . 1 (3): 155–7. дои : 10.1016/j.cmet.2005.02.005 . ПМИД 16054057 .
- ^ Ду Ф, Цянь С, Цянь ЗМ, Ву XM, Се Х, Юнг ВХ, Кэ Ю (июнь 2011 г.). «Гепцидин напрямую ингибирует экспрессию рецептора трансферрина 1 в астроцитах через путь циклической АМФ-протеинкиназы А». Глия . 59 (6): 936–45. дои : 10.1002/glia.21166 . ПМИД 21438013 . S2CID 25678955 .
- ^ Мошенничество, Вишант Махендра; Радже, Манодж; Радже, Чаая Айенгар (1 декабря 2014 г.). «Подработка белка в метаболизме железа: глицеральдегид-3-фосфатдегидрогеназа (ГАФДГ)». Труды Биохимического общества . 42 (6): 1796–1801. дои : 10.1042/BST20140220 . ПМИД 25399609 .
- ^ Шеоканд Н., Малхотра Х., Кумар С., Тиллу В.А., Чаухан А.С., Радже С.И., Радже М. (октябрь 2014 г.). «Подрабатывающая GAPDH на клеточной поверхности рекрутирует апотрансферрин, чтобы обеспечить выход железа из клеток млекопитающих» . Журнал клеточной науки . 127 (Часть 19): 4279–91. дои : 10.1242/jcs.154005 . ПМИД 25074810 .
- ^ Берту, Кристиан; Илиу, Жан Поль; Барба, Денис (5 декабря 2021 г.). «Железо, нейробиодоступность и депрессия» . Э.Дж.Хем . 3 (1): 263–275. дои : 10.1002/jha2.321 . ISSN 2688-6146 . ПМЦ 9175715 . ПМИД 35846210 .
- ^ Каутц Л., Юнг Г., Валоре Э.В., Ривелла С., Немет Э., Ганц Т. (июль 2014 г.). «Идентификация эритроферрона как эритроидного регулятора обмена железа» . Природная генетика . 46 (7): 678–84. дои : 10.1038/ng.2996 . ПМК 4104984 . ПМИД 24880340 .
- ^ Мукенталер М.Ю., Гали Б., Хентце М.В. (2008). «Системный гомеостаз железа и регуляторная сеть железо-чувствительного элемента/железо-регуляторного белка (IRE/IRP)». Ежегодный обзор питания . 28 : 197–213. дои : 10.1146/annurev.nutr.28.061807.155521 . ПМИД 18489257 .
- ^ Халлберг Л. (1987). «Пшеничная клетчатка, фитаты и всасывание железа». Scand J Гастроэнтерол Suppl . 129 : 73–9. дои : 10.3109/00365528709095855 . ПМИД 2820048 .
- ^ Линч С.Р. (2000). «Влияние кальция на всасывание железа» . Нутр Рес Преподобный . 13 (2): 141–58. дои : 10.1079/095442200108729043 . ПМИД 19087437 .
- ^ Дислер П.Б., Линч С.Р., Чарльтон Р.В., Торранс Дж.Д., Ботвелл Т.Х., Уокер Р.Б. и др. (1975). «Влияние чая на всасывание железа» . Гут . 16 (3): 193–200. дои : 10.1136/gut.16.3.193 . ПМЦ 1410962 . ПМИД 1168162 .
- ^ "До настоящего времени" .
- ^ Аннибале, Бруно; Капурсо, Габриэле; Мартино, Джина; Гросси, Кристина; Делле Фаве, Джанфранко (декабрь 2000 г.). «Железодефицитная анемия и инфекция Helicobacter pylori». Международный журнал противомикробных средств . 16 (4): 515–519. дои : 10.1016/s0924-8579(00)00288-0 . ПМИД 11118871 .
- ^ Компакт-диск Рудольфа (2003). Педиатрия Рудольфа . Нью-Йорк: МакГроу-Хилл, Медицинский паб. Разделение. ISBN 978-0-07-112457-7 .
- ^ Пинникс, Зандра К.; Миллер, Лэнс Д.; Ван, Вэй; Д'Агостино, Ральф; Куте, Тим; Уиллингем, Марк К.; Хэтчер, Хизер; Тесфай, Лия; Суй, Гуанчао (4 августа 2010 г.). «Регуляция ферропортина и железа в прогрессировании и прогнозе рака молочной железы» . Наука трансляционной медицины . 2 (43): 43ра56. doi : 10.1126/scitranslmed.3001127 . ISSN 1946-6234 . ПМЦ 3734848 . ПМИД 20686179 .
- ^ Jump up to: а б Тиммерс, Пол Р.Х.Дж.; Уилсон, Джеймс Ф.; Джоши, Питер К.; Дилен, Йорис (июль 2020 г.). «Многовариантное геномное сканирование позволяет выявить новые локусы и метаболизм гема в старении человека» . Природные коммуникации . 11 (3570): 3570. Бибкод : 2020NatCo..11.3570T . дои : 10.1038/s41467-020-17312-3 . ПМЦ 7366647 . ПМИД 32678081 .
- ^ Раметта, Рафаэла; Мерони, Марика; Донджованни, Паола (15 мая 2020 г.). «От окружающей среды к геному и обратно: урок мутаций HFE» . Международный журнал молекулярных наук . 21 (10): 3505. doi : 10.3390/ijms21103505 . ПМК 7279025 . ПМИД 32429125 .
- ^ Северин С.Дж., Шинде У., Ротвейн П. (сентябрь 2009 г.). «Молекулярная биология, генетика и биохимия семейства молекул отталкивающего направления» . Биохимический журнал . 422 (3): 393–403. дои : 10.1042/BJ20090978 . ПМЦ 4242795 . ПМИД 19698085 .
Дальнейшее чтение
[ редактировать ]- Эндрюс С., Нортон И., Салунхе А.С., Гудлак Х., Али В.С., Мурад-Ага Х., Корнелис П. (2013). «Глава 7, Контроль метаболизма железа в бактериях». В Banci L (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. стр. 203–39. дои : 10.1007/978-94-007-5561-1_7 . ISBN 978-94-007-5560-4 . ПМИД 23595674 . электронная книга ISBN 978-94-007-5561-1 ISSN 1559-0836 электронный- ISSN 1868-0402
- Эндрюс, Северная Каролина (май 2004 г.). «Анемия воспаления: цитокин-гепсидиновая связь» . Журнал клинических исследований . 113 (9): 1251–3. дои : 10.1172/JCI21441 . ПМЦ 398435 . ПМИД 15124013 .
- Камашелла С (декабрь 2005 г.). «Понимание гомеостаза железа посредством генетического анализа гемохроматоза и связанных с ним нарушений» . Кровь . 106 (12): 3710–7. дои : 10.1182/кровь-2005-05-1857 . ПМИД 16030190 .
- Фрейзер Д.М., Андерсон Дж.Дж. (октябрь 2005 г.). «Импорт железа. I. Кишечная абсорбция железа и ее регуляция». Американский журнал физиологии. Физиология желудочно-кишечного тракта и печени . 289 (4): G631–5. дои : 10.1152/ajpgi.00220.2005 . ПМИД 16160078 .
- Инсел П., Росс Д., МакМахон К., Бернштейн М. (2011). "Железо" . Питание (4-е изд.). Садбери, Массачусетс: Издательство Jones and Bartlett . стр. 510–514. ISBN 978-0-7637-7663-3 . Проверено 25 июня 2012 г. См. особенно. стр. 513-514.
- Ламми-Киф CJ, Couch SC, Philipson EH, ред. (2008). «Диетическая диверсификация и модификация железа» . Справочник по питанию и беременности . Питание и здоровье. Тотова, Нью-Джерси: Humana Press . стр. 350–351. дои : 10.1007/978-1-59745-112-3 . ISBN 978-1-59745-112-3 . Проверено 25 июня 2012 г.
- Группа экспертов по микроэлементам; Подкомитеты по верхним референтным уровням питательных веществ, а также по интерпретации и использованию эталонных норм потребления с пищей; Постоянный комитет по научной оценке эталонного потребления пищевых продуктов (2001 г.). "Железо" . Рекомендуемая диетическая норма витамина А, витамина К, мышьяка, бора, хрома, меди, йода, железа, марганца, молибдена, никеля, кремния, ванадия и цинка . Вашингтон, округ Колумбия: Совет по продовольствию и питанию, Медицинский институт . стр. 290–393. ISBN 978-0-309-07279-3 . Проверено 25 июня 2012 г.
- Рейли С. (2004). "Железо" . Пищевые микроэлементы . Оксфорд, Великобритания и Эймс, Айова: Blackwell Publishing . стр. 35–81. ISBN 978-1-4051-1040-2 . Проверено 25 июня 2012 г.
Внешние ссылки
[ редактировать ]- Подробный информационный бюллетень NIH по железу и питанию
- Институт заболеваний железа: некоммерческая группа, занимающаяся расстройствами железа; сайт содержит полезные ссылки и информацию о заболеваниях, связанных с железом.
- Интерактивный портал медицинского обучения по метаболизму железа
- Информация о железе вне организма