Белок-проводник
Белки-диригенты относятся к классу белков , которые определяют стереохимию соединений, синтезируемых другими ферментами. [1] Первый диригентный белок был обнаружен у Forsythia intermedia . Было обнаружено, что этот белок управляет стереоселективным биосинтезом (+)-пинорезинола из мономеров кониферилового спирта: [2]

лигнанов Биосинтез катализируется ферментами окислительными . [3] В пробирке в результате реакции образуется гетерогенная смесь димерных соединений. [4] Когда во время реакции присутствует прямой белок, один стереоизомер одного соединения сильно обогащается. Диригентные белки, по-видимому, не обладают собственной активностью по образованию окислительных радикалов; в отсутствие окислительного фермента реакция не произойдет. [5]
был идентифицирован второй энантиоклементарный диригентный белок Недавно у Arabidopsis thaliana , который управляет энантиоселективным синтезом (-)- пинорезинола . [6]
Активность
[ редактировать ]При биосинтезе лигнанов окислительные ферменты осуществляют протон-связанный перенос электронов, чтобы удалить атом водорода из монолигнолов , образуя радикальный промежуточный продукт. Эти промежуточные соединения затем соединяются в реакции терминации радикала с образованием одного из множества димеров, известных как лигнаны . [7] in vitro Реакции с конифериловым спиртом (обычным монолигнолом ) в присутствии окислительных ферментов производят большое количество различных димеров в различных концентрациях. [8] диригентный белок из Forsythia intermedia Когда присутствует , производство (+)- пинорезинола значительно увеличивается, а других продуктов гораздо меньше. Поскольку это обогащение настолько выражено, предполагается, что фермент продуцирует исключительно (+)- пинорезинол и конкурирует с небелково-опосредованной реакцией сочетания, которая приводит к образованию гетерологичной смеси продуктов. [9] Это было подтверждено анализом различных смесей, полученных с различными концентрациями присутствующих белков-диригентов. Механизм, с помощью которого достигается такая стереоселективность, в настоящее время недостаточно изучен. Однако, поскольку в отсутствие окислительных ферментов никакая реакция не протекает, диригентный белок сам по себе не катализирует окисление кониферилового спирта с образованием радикалов.
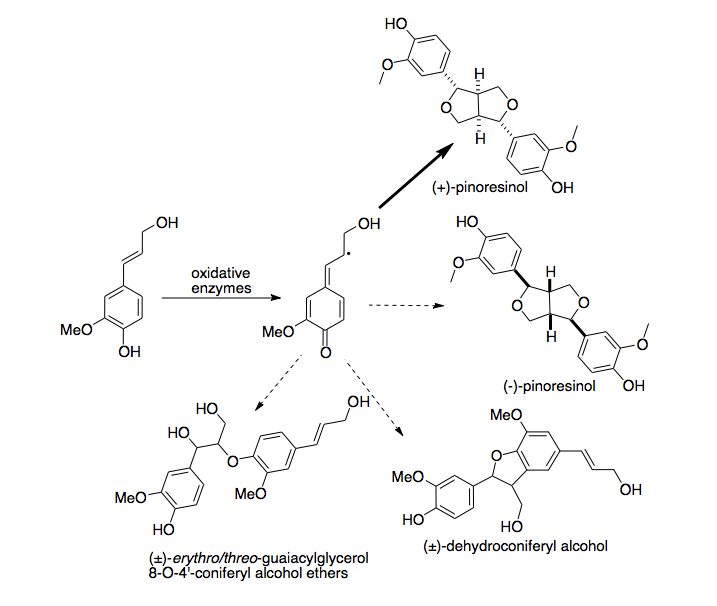
Активность диригентного белка Forsythia intermedia специфична для кониферилового спирта. [10] Когда другие монолигнолы, такие как п-кумариловый спирт и синапиловый спирт , вступают в реакцию in vitro с окислительными ферментами в присутствии диригентного белка, они образуют гетерологичную смесь продуктов, неотличимую от идентичных экспериментов в отсутствие диригентного белка.
Структура
[ редактировать ]Эксперименты с круговым дихроизмом показали, что вторичная структура диригентного белка Forsythia intermedia состоит в основном из β-складчатых листов и петлевых структур. Третичная структура не выяснена, но было подтверждено, что белок является димерным. Каждый димер имеет один сайт связывания кониферилового спирта, всего два сайта связывания. [11] Один конифериловый спирт может связываться с каждым сайтом, так что геометрия реакции между ними ограничена, увеличивая выработку (+)-пинорезинола и подавляя выработку других продуктов.
Биологическое значение
[ редактировать ]В отсутствие диригентного белка пинорезинол является относительно второстепенным продуктом биосинтеза лигнас. Когда присутствует диригентный белок, он становится основным продуктом. Биологическое значение (+)-пинорезинола в растениях до конца не изучено, но было обнаружено, что он эффективен в качестве средства отпугивания муравьев гусениц капустной бабочки , которые получают это соединение из своего рациона. [12] Это может служить аналогичной защитной цели у Forsythia intermedia . [13]
Ссылки
[ редактировать ]- ^ Николсон, Ральф Л.; Уилфред Вермеррис; Вермеррис Уилфред (2006). Биохимия фенольных соединений . Берлин: Шпрингер. С. 107–108 . ISBN 1-4020-5163-8 .
- ^ Дэвин Л.Б., Ван Х.Б., Кроуэлл А.Л. и др. (1997). «Стереоселективное бимолекулярное связывание фенокси-радикала с помощью вспомогательного (направляющего) белка без активного центра». Наука . 275 (5298): 362–6. дои : 10.1126/science.275.5298.362 . ПМИД 8994027 . S2CID 41957412 .
- ^ Уорд РС (1982). «Синтез лигнанов и неолигнанов» . Обзоры химического общества . 11 (2): 75–125. дои : 10.1039/CS9821100075 .
- ^ Фурнан Д., Катала Б., Лапьер К. (январь 2003 г.). «Начальные этапы катализируемой пероксидазой полимеризации кониферилового спирта и / или синапилового альдегида: исследование влияния pH на капиллярный зональный электрофорез». Фитохимия . 62 (2): 139–46. дои : 10.1016/S0031-9422(02)00573-3 . ПМИД 12482448 .
- ^ Дэвин Л.Б.; Льюис Н.Г. (август 2005 г.). «Сочетание диригентных фенокси-радикалов: достижения и проблемы». Современное мнение в области биотехнологии . 16 (4): 398–406. дои : 10.1016/j.copbio.2005.06.010 . ПМИД 16023845 .
- ^ Пикель Б., Константин М.А., Пфаннштайл Дж., Конрад Дж., Бейфусс Ю., Шаффер А. (март 2007 г.). «Энантиоклементарный диригентный белок для энантиоселективного лакказо-катализируемого окислительного сочетания фенолов» . Ангеванде Хеми . 53 (4): 273–284. дои : 10.1007/s10086-007-0892-x . S2CID 195313754 .
- ^ Сарканен, Симо; Льюис, Норман (1998). Лигнин и биосинтез лигнанов . Колумбус, Огайо: Американское химическое общество. ISBN 0-8412-3566-Х .
- ^ Фурнан Д., Катала Б., Лапьер К. (январь 2003 г.). «Начальные этапы катализируемой пероксидазой полимеризации кониферилового спирта и / или синапилового альдегида: исследование влияния pH на капиллярный зональный электрофорез». Фитохимия . 62 (2): 139–46. дои : 10.1016/S0031-9422(02)00573-3 . ПМИД 12482448 .
- ^ Холлс С.К., Дэвин Л.Б., Крамер Д.М. , Льюис Н.Г. (2004). «Кинетическое исследование связывания радикала кониферилового спирта с (+)-пинорезинолом, образующим диригентный белок». Биохимия . 43 (9): 2587–95. дои : 10.1021/bi035959o . ПМИД 14992596 .
- ^ Ким, МК; Чон Ж.-Х.; Фудзита М.; Дэвин Л.Б.; Льюис Н.Г. (2002). «Семейство DIRIGENT западного красного кедра (Thuja plicata) 8-8' демонстрирует разнообразные модели экспрессии и консервативную специфичность связывания монолинола». Молекулярная биология растений . 49 (2): 199–214. дои : 10.1023/А:1014940930703 . ПМИД 11999375 . S2CID 8042373 .
- ^ Холлс СК, Льюис Н.Г. (июль 2002 г.). «Вторичные и четвертичные структуры (+)-пинорезинол-образующего диригентного белка». Биохимия . 41 (30): 9455–61. дои : 10.1021/bi0259709 . ПМИД 12135368 .
- ^ Шредер ФК, дель Кампо М.Л., Грант Дж.Б. и др. (октябрь 2006 г.). «Пинорезинол: лигнол растительного происхождения, служащий для защиты гусениц» . Учеб. Натл. акад. наук. США . 103 (42): 15497–501. дои : 10.1073/pnas.0605921103 . ПМК 1622851 . ПМИД 17030818 .
- ^ Шмитт Дж., Петерсон М. (2002). «Накопление пинорезинола и матарезинола в суспензионной культуре клеток Forsythia × intermedia». Культура растительных клеток, тканей и органов . 68 (1): 91–98. дои : 10.1023/А:1012909131741 . S2CID 19282302 .