Лили вирус х
| Лили вирус х | |
|---|---|
| Классификация вирусов | |
| (не вмешательство): | Вирус |
| Область : | Рибовирия |
| Королевство: | Orthornavirae |
| Филум: | Kitrinoviricota |
| Сорт: | Alsuviricetes |
| Заказ: | Tymovirales |
| Семья: | Alphaflexiviridae |
| Род: | Potexvirus |
| Разновидность: | Лили вирус х
|
Вирус Lily X ( LVX ) представляет собой патогенный вирус растений SSRNA (+) семейства Alphaflexiviridae и порядка тимовирал .
Описание
[ редактировать ]LVX описан в 4 -м докладе ICTV (1982). Он встречается в первую очередь в лилиях, хотя больше растений восприимчиво, и, как полагают, вирус передается только механически. Там нет известных векторов; Тем не менее, применение инсектицидов эффективно снижало распространение LVX, из которого можно собрать, что передача вируса опосредована насекомыми. [ 1 ] Симптомы этого вируса пока неизвестны, запутывая способность изучить степень естественной инфекции и распространения.
Структура
[ редактировать ]
LVX-это неразвитый вирус с спиральной симметрией. Считается, что все Potexvirus, в том числе LVX, имеют чуть менее 9 белковых субъединиц на спиральный поворот. [ 3 ] Эта схема образования нуклеокапсида заставляет нуклеокапсид удлиненным, гибким, нитевидным вирусом, как и большинство вирусов растений. В отличие от других potexvirus (средней длины 550 нм), LVX имеет длину 470 нм и ширину 13 нм. [ 4 ] LVX отличается от лилии без симптомов карлавируса (LSV) с помощью серологических тестов, таких как тесты на маркировку иммуноголда. [ 5 ]
Геном
[ редактировать ]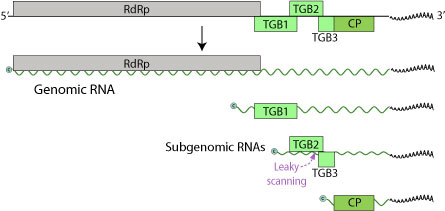
Геном LVX состоит из одной отдельной (+) РНК-последовательности в размере 5,9-7 килобаз. Геном содержит только три открытых кадра считывания (ORF), кодирующие пять белков (RDRP, TGBP1, TGBP2, TGBP3, CP). [ 6 ] 5 'конец ограничен, а 3' терминус полиаденилирован.
Эти белки составляют 24 килодальтона (KDA), 12 кДа и 22 кДа, а третий-белок пальто, а также кодирует РНК-зависимую РНК-полимеразу (RDRP). [ 7 ] Стоп -кодон ORF 24 кДа и начальный кодон 12 кДа ORF перекрывают друг друга. В то время как многие геномы Potexvirus содержат небольшой ORF, который сразу же составляет 5 футов этого белка для пальто, это не обнаруживается в геноме LVX. Эта разница удивительна, так как этот ORF был идентифицирован как минимум в шести других геномах Potexvirus. Однако, глядя на белки, кодируемые LVX и другими potexvirus, между аминокислотными последовательностями было значительное сходство. На 5 -дюймовом конце LVX есть усеченная ORF, которая кодирует для полипептида, который содержит мотив GDD. Этот мотив также обнаружен в С-концевом конце белков, кодируемых другими potexvirus. Исследование белка 24 кДа LVX показывает, что он содержит мотив связывания нуклеозида трифосфата (gxxgxgks/t). Этот мотив также обнаружен в белке без симптомов лилии 25 кДа, а также в белках 25-26 кДа других potexvirus, таких как вирус картофеля и вирус картофеля S.
LVX также содержит предполагаемую последовательность субгеномного промотора Potexvirus (GGTTAAGTT --- GAA) вверх по течению (5 ') белка 24 кДа. Эта последовательность также обнаружена выше по течению кодонов инициации белков пальто в аналогичных вирусах, особенно без симптомов лилии. Исследователи посмотрели на листья, инфицированные LVX, и обнаружили наличие субгеномных РНК. Эти РНК были около 2000 и 850 нуклеотидов в длину, что соответствовало предполагаемым субгеномным промоторам. Исходя из этого, было установлено, что они, вероятно, функционируют как посланники как для белков 24 кДа, так и для 22 кДа (белок пальто). В последовательности LVX существует область между 12 кДа ORF и ORF 22 кДа (белок пальто), которая не переводится. В других последовательностях Potexvirus эта область содержит соответствующие 7 кДа и 11 кДа орф; Однако в этой области последовательности LVX существует непрерывная рамка считывания. Эта рамка считывания начинается в положении нуклеотида 1236 и заканчивается в позиции 1646 года. Если эта область должна была быть переведена, кодируемый выше белок будет аналогичен белкам 7 кДа и 11 кДа, упомянутым выше, заканчивая 18 нуклеотидов внутри белка для покрытия LVX. Перевод этого LVX ORF не переводится, потому что в геноме отсутствует кодон инициации.
Блок 1 Triple Gene (TGBP1) представляет собой многофункциональный белок. Он действует, чтобы способствовать трансляции вирусных РНК, действуя как РНК- геликаза , разделяя двухцепочечную РНК для функций RDRP. Более того, он может выступать в качестве супрессора РНК -интерференции , который является иммунной защитой от накопления вирусных РНК. [ 8 ] TGBP1 транспортирует вирусный геном в соседние растительные клетки непосредственно через Plasmosdesmata , что позволяет эффективно распространение путем обхода клеточной стенки хозяина. [ 9 ] TGBP1 также увеличивает пределы исключения размеров плазмодесмального размера, позволяя вирусному геному легче перемещаться от одной клетки в другую. Наконец, он подавляет молчание РНК, предотвращая отключение генома РНК.
TGBP2 и TGBP3 являются мембранными связывающими белками, участвующими в прикреплении и входе. TGBP3 экспрессируется посредством протекающей сканирования субгеномной мРНК TGBP2. ORF TGBP2 имеет слабый кодон инициации, который иногда пропускается рибосомой в инициации перевода. В этом случае рибосомная субъединица 40 -х годов продолжает сканироваться, пока не столкнется с кодоном инициации TGBP3 и не начинает перевод.
CP является белком Coat LVX, который образует рибонуклеопротеиновые комплексы наряду с TGBP1 и вирусной мРНК.
«Нуклеотидная последовательность LVX, по -видимому, является уникальной среди potexvirus, поскольку в ней, по -видимому, не хватает небольшой открытой рамки считывания , 5 футов для белка Coat Cistron, общего для всех других potexvirus». [ 10 ] Кроме того, в области TGBP3 генома не хватало нормального начального кодона. [ 11 ] Область из 5'-некодирования начинается с GGAAAA, тогда как у других секвенированных potexvirus, вероятно, все начинаются с GAAAA. [ 12 ] Филогенетический анализ кодирующей последовательности LVX показал, что LVX был наиболее тесно связан с вирусом из мягкого желтого края клубники . [ 13 ]

Репликация
[ редактировать ]LVX не имеет известного вектора, но он, скорее всего, распространяется и входит в ячейку посредством механической инокуляции насекомыми. Репликация LVX, как и другие вирусы ssRNA (+), происходит в цитоплазме клеток. Как только вирус попадает в клетку -хозяина, вирус не покрыт покрытием и выпускает РНК вирусного генома в цитоплазму. Вирусная моноцистронная РНК затем транслируется в РНК-зависимую РНК-полимеразу, кодируемой 5'-проксимальным ORF. Репликация LVX происходит на вирусных фабриках, которые организованы белком TGBP1. [ 15 ] TGBP1 работает, чтобы переставить актин и эндомбраны хозяина и создает сборку спиральных аранжировок. Эти спиральные структуры окружены эндомбранами хозяина, которые создают область, где вирус может реплицироваться более эффективно. [ 16 ]
Репликация вируса ssRNA (+) генерирует вирус дцРНК (+), который затем дополнительно транскрибируется и реплицируется для создания большего количества мРНК LVX и геномов SSRNA (+) соответственно. Субгеномные промоторы на вирусе приводят к трансляции SgrNAS, что приводит к образованию капсида и движений. Они используются для распоряжения структуры вириона и организовать вирусные геномы. Затем новые частицы вируса могут быть получены и собраны. С помощью белков с тройным геном LVX эти завершенные частицы могут пройти большие расстояния среди Plasmodesmata и более коротких межклеточных пространств, чтобы заразить другие клетки -хозяина. [ 17 ]
Связанная болезнь
[ редактировать ]LVX не имеет известных связанных заболеваний.
Симптомы
[ редактировать ]Нет никаких известных выдающихся симптомов, которые возникают в естественном диапазоне LVX с лилиемной формосаной ; Однако Ян (1997) обнаружил, что инфицированные LVX лилии растут медленнее и имеют тенденцию быть меньше, чем неинфицированные образцы. Механическая инокуляция других восприимчивых видов растений, таких как тетрагония тетрагоиды и Chenopodium murale , приводят к хлоротическим местным поражениям. [ 18 ]
Тропизм
[ редактировать ]Несмотря на то, что они не видны в природе, лабораторные тесты показали, что LVX способен заразить несколько других растений за пределами лилиевой формы . [ 19 ] Другие виды, которые, как было показано, восприимчивы: [ 20 ]
- Chenopodium Head
- Chenopodium murale
- Chenopodium Quinoa
- Gomphrena Globose
- Лили Формосанум
- Никотиана Бентамана
- Никотиана Кливлендий
- Тетрагония тетрагоноид
LVX способен заразить все ткани растения лилии. Вирионы были цитопатологически обнаружены во всех частях растений -хозяев и содержат приблизительно 5% нуклеиновой кислоты и 95% белка, без липидов. [ 21 ] Ожидается, что режим передачи будет механической инокуляцией через векторы насекомых, поскольку распространение вируса ингибировалось инсектицидами, но не спрея минерала-масла. [ 22 ]
Ссылки
[ редактировать ]- ^ Asjes, CJ (1991). Контроль воздушного полевого распространения вируса, разбивающего тюльпан, бессмысленного вируса и вируса лилии, в лилиях минеральными маслами, синтетическими пиретроидами и нематида в Нидерландах Нидерланды Журнал Plate Path, 97 (3), 129-138.
- ^ Кендалл, А., Макдональд, М., Биан, В., Боулз, Т., Баумгартен, С.С., Ши, Дж., ... Стаббс, Г. (2008). Структура гибких нитевидных вирусов растений. Journal of Virology, 82 (19), 9546–9554.
- ^ Кендалл, А., Макдональд, М., Биан, В., Боулз, Т., Баумгартен, С.С., Ши, Дж., ... Стаббс, Г. (2008). Структура гибких нитевидных вирусов растений. Journal of Virology, 82 (19), 9546–9554.
- ^ Стоун, Ом (1980). Два новых Potexvirus от Monocotyledons. Acta Hort. 110, 59-63.
- ^ Ян, Т. (1997). Цитологические характеристики и обнаружение вирусов лилии SPP. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Memelink, J., van der Vlugt, Cim, Linthorst, HJM, Derks, Aflm, Asjes, CJ, Bol, JF (1990). Гомологии между геномами карлавируса (бессмысленный вирус лилии) и Potexvirus (вирус Lily x) из растений лилии. Журнал общей вирусологии, 71, 917-924.
- ^ Memelink, J., van der Vlugt, Cim, Linthorst, HJM, Derks, Aflm, Asjes, CJ, Bol, JF (1990). Гомологии между геномами карлавируса (бессмысленный вирус лилии) и Potexvirus (вирус Lily x) из растений лилии. Журнал общей вирусологии, 71, 917-924.
- ^ Lubicz-Verchot, J. (2005). Новая модель транспорта клетки к клеткам для Potexviruses. Молекулярные взаимодействия растений-микробов, 18 (4), 283-290.
- ^ Lubicz-Verchot, J. (2005). Новая модель транспорта клетки к клеткам для Potexviruses. Молекулярные взаимодействия растений-микробов, 18 (4), 283-290.
- ^ Ян, Т. (1997). Цитологические характеристики и обнаружение вирусов лилии SPP. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Chen, J., Shi, Y.-H., Adams, MJ, Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архивы вирусологии, 150 (4), 825-832.
- ^ Chen, J., Shi, Y.-H., Adams, MJ, Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архивы вирусологии, 150 (4), 825-832.
- ^ Chen, J., Shi, Y.-H., Adams, MJ, Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архивы вирусологии, 150 (4), 825-832.
- ^ Chen, J., Shi, Y.-H., Adams, MJ, Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архивы вирусологии, 150 (4), 825-832.
- ^ Ян, Т. (1997). Цитологические характеристики и обнаружение вирусов лилии SPP. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Lubicz-Verchot, J. (2005). Новая модель транспорта клетки к клеткам для Potexviruses. Молекулярные взаимодействия растений-микробов, 18 (4), 283-290.
- ^ Ян, Т. (1997). Цитологические характеристики и обнаружение вирусов лилии SPP. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Стоун, Ом (1980). Два новых Potexvirus от Monocotyledons. Acta Hort. 110, 59-63.
- ^ Стоун, Ом (1980). Два новых Potexvirus от Monocotyledons. Acta Hort. 110, 59-63.
- ^ Филлипс, S (1986). "Лили X Potexvirus" . Вирусы растений онлайн . Архивировано из оригинала 8 декабря 2015 года . Получено 7 декабря 2015 года .
- ^ Стоун, Ом (1980). Два новых Potexvirus от Monocotyledons. Acta Hort. 110, 59-63.
- ^ Asjes, CJ (1991). Контроль воздушного поля распространяется вирусом, разбивающим тюльпан, бессмысленным вирусом лилии и вирусом лилии в лилий минеральными маслами, синтетическими пиретроидами и нематидом в Нидерландах. Нидерландов Журнал Plant Path, 97 (3), 129-138.