Бактериальный рост

Рост бактерий — это распространение бактерии на две дочерние клетки в процессе, называемом бинарным делением . Если не происходит мутации, полученные дочерние клетки генетически идентичны исходной клетке. Следовательно, происходит рост бактерий. Обе дочерние клетки после деления не обязательно выживают. Однако если число выживших в среднем превышает единицу, популяция бактерий подвергается экспоненциальному росту . Измерение экспоненциальной кривой роста бактерий в периодической культуре традиционно было частью обучения всех микробиологов; базовый метод требует подсчета бактерий (подсчета клеток) прямым и индивидуальным (микроскопическим, проточным цитометрическим методом) [ 1 ] ), прямой и групповой (биомасса), косвенный и индивидуальный (подсчет колоний) или косвенный и групповой (наиболее вероятное количество, мутность , поглощение питательных веществ) методы. Модели согласовывают теорию с измерениями. [ 2 ]
Фазы
[ редактировать ]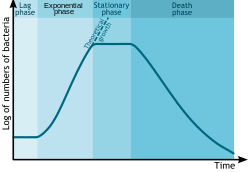
В аутэкологических исследованиях рост бактерий (или других микроорганизмов, таких как простейшие , микроводоросли или дрожжи ) в периодической культуре можно моделировать с помощью четырех различных фаз: лаг-фаза (А), лог-фаза или экспоненциальная фаза (В), стационарная фаза (С). ) и фаза смерти (D). [ 3 ]
- Во время лаг-фазы бактерии адаптируются к условиям роста. Это период, когда отдельные бактерии созревают и еще не способны делиться. Во время лаг-фазы цикла роста бактерий происходит синтез РНК, ферментов и других молекул. Во время лаг-фазы клетки изменяются очень мало, поскольку клетки не сразу размножаются в новой среде. Этот период практически полного отсутствия деления клеток называется лаг-фазой и может длиться от 1 часа до нескольких дней. На этом этапе клетки не находятся в состоянии покоя. [ 4 ]
- Логарифмическая фаза (иногда называемая логарифмической фазой или экспоненциальной фазой ) — это период, характеризующийся удвоением клеток. [ 5 ] Число новых бактерий, появляющихся в единицу времени, пропорционально существующей популяции. Если рост не ограничен, удвоение будет продолжаться с постоянной скоростью, поэтому и количество клеток, и скорость роста популяции удваиваются с каждым последующим периодом времени. Для этого типа экспоненциального роста график зависимости натурального логарифма числа клеток от времени дает прямую линию. Наклон этой линии представляет собой удельную скорость роста организма, которая является мерой количества делений клетки в единицу времени. [ 5 ] Фактическая скорость этого роста (т.е. наклон линии на рисунке) зависит от условий роста, которые влияют на частоту делений клеток и вероятность выживания обеих дочерних клеток. В контролируемых условиях цианобактерии могут удваивать свою популяцию четыре раза в день, а затем утроить свою популяцию. [ 6 ] Однако экспоненциальный рост не может продолжаться бесконечно, поскольку среда вскоре истощается питательными веществами и обогащается отходами.
- Стационарная фаза часто возникает из-за фактора, ограничивающего рост, такого как истощение необходимого питательного вещества и/или образование ингибирующего продукта, такого как органическая кислота. Стационарная фаза возникает в ситуации, когда темпы роста и смертности равны. Количество создаваемых новых клеток ограничено фактором роста, и в результате скорость роста клеток соответствует скорости их гибели. В результате получается «гладкая» горизонтальная линейная часть кривой во время стационарной фазы. Мутации могут возникать во время стационарной фазы . Бриджес и др. (2001) [ 7 ] представили доказательства того, что повреждение ДНК ответственно за многие мутации, возникающие в геномах бактерий, находящихся в стационарной фазе или голодающих. Эндогенно генерируемые активные формы кислорода, по-видимому, являются основным источником таких повреждений. [ 7 ] Бактерии в этой фазе иногда впадают в состояние покоя , используя факторы гибернации для замедления своего метаболизма. [ 8 ]
- На стадии смерти (фаза упадка) бактерии погибают. Это может быть вызвано недостатком питательных веществ, температурой окружающей среды выше или ниже допустимого диапазона для данного вида или другими вредными условиями.
Эта базовая модель роста периодической культуры выделяет и подчеркивает аспекты роста бактерий, которые могут отличаться от роста макрофауны. Он подчеркивает клональность, бесполое бинарное деление, короткое время развития по сравнению с самой репликацией, кажущуюся низкую смертность, необходимость перехода от спящего состояния к репродуктивному состоянию или кондиционирования среды и, наконец, тенденцию к появлению лабораторно адаптированных штаммов. исчерпать свои питательные вещества. В действительности, даже в периодической культуре четыре фазы четко не определены. Клетки не размножаются синхронно без явной и постоянной подсказки (как в экспериментах со стебельчатыми бактериями). [ 9 ] ), и их экспоненциальный фазовый рост часто не всегда является постоянной скоростью, а вместо этого медленно затухающей скоростью, постоянной стохастической реакцией на давление как на воспроизводство, так и на спячку перед лицом снижения концентрации питательных веществ и увеличения концентрации отходов.
Уменьшение числа бактерий может даже стать логарифмическим. Следовательно, эту фазу роста можно также назвать фазой отрицательного логарифма или отрицательной экспоненциальной фазы роста. [1]
Ближе к концу логарифмической фазы периодической культуры способность к естественной генетической трансформации может быть индуцирована , как у Bacillus subtilis. [ 10 ] и у других бактерий. Естественная генетическая трансформация — это форма переноса ДНК, которая, по-видимому, является адаптацией для восстановления повреждений ДНК.
Периодическое культивирование — наиболее распространенный метод лабораторного выращивания, с помощью которого изучается рост бактерий, но это лишь один из многих. В идеале оно пространственно неструктурировано и структурировано во времени. Бактериальную культуру инкубируют в закрытом сосуде с одной порцией среды. В некоторых экспериментальных режимах часть бактериальной культуры периодически удаляют и добавляют в свежую стерильную среду. В крайнем случае это приводит к постоянному обновлению питательных веществ. Это хемостат , также известный как непрерывная культура. В идеале он неструктурирован в пространстве и во времени, находится в устойчивом состоянии, определяемом скоростью поступления питательных веществ и роста бактерий. По сравнению с периодической культурой бактерии поддерживаются в фазе экспоненциального роста, и скорость роста бактерий известна. Сопутствующие устройства включают турбидостаты и ауксостаты . Когда Escherichia coli растет очень медленно со временем удвоения 16 часов в хемостате, большинство клеток имеют одну хромосому. [ 1 ]
Рост бактерий можно подавить с помощью бактериостатов , не обязательно убивая бактерии. Определенные токсины можно использовать для подавления роста бактерий или уничтожения бактерий. Антибиотики (или, точнее, антибактериальные препараты) — это препараты, используемые для уничтожения бактерий; они могут иметь побочные эффекты или даже вызывать побочные реакции у людей, однако не относятся к токсинам. В синэкологической , естественной ситуации, в которой присутствует более одного вида бактерий, рост микробов более динамичен и непрерывен.
Жидкость – не единственная лабораторная среда для роста бактерий. Пространственно структурированные среды, такие как биопленки или поверхности агара , представляют собой дополнительные сложные модели роста.
5-я фаза: Долгосрочная стационарная фаза.
[ редактировать ]Долгосрочная стационарная фаза, в отличие от ранней стационарной фазы (когда деление клеток незначительно), представляет собой высокодинамичный период, в котором уровни рождаемости и смертности сбалансированы. Доказано, что после фазы смерти E. coli можно поддерживать в периодической культуре в течение длительного времени без добавления питательных веществ. [ 11 ] [ 12 ] Предоставляя стерильную дистиллированную воду для поддержания объема и осмолярности, аэробно выращенные культуры можно поддерживать при плотности ~10 6 колониеобразующих единиц (КОЕ) на мл в течение более 5 лет без добавления питательных веществ в периодической культуре. [ 13 ]
Условия окружающей среды
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( октябрь 2016 г. ) |
Факторы окружающей среды влияют на скорость роста бактерий, такие как кислотность (pH), температура, активность воды, макро- и микроэлементы, уровень кислорода и токсины. Условия, как правило, относительно одинаковы между бактериями, за исключением экстремофилов . Бактерии имеют оптимальные условия роста, при которых они процветают, но за пределами этих условий стресс может привести либо к замедлению, либо к остановке роста, состоянию покоя (например, образования спор ) или смерти. Поддержание неоптимальных условий роста является ключевым принципом сохранения продуктов питания .
Температура
[ редактировать ]Низкие температуры имеют тенденцию замедлять темпы роста, что привело к тому, что охлаждение играет важную роль в сохранении продуктов питания. В зависимости от температуры бактерии подразделяются на:
- Психрофилы
Психрофилы — экстремофильные холодолюбивые бактерии или археи с оптимальной температурой роста около 15 °С и ниже (максимальная температура роста — 20 °С, минимальная температура роста — 0 °С и ниже). Психрофилы обычно встречаются в чрезвычайно холодных экосистемах Земли, таких как полярные ледяные шапки, вечная мерзлота, полярная поверхность и глубокие океаны. [ 14 ]
- Мезофилы
Мезофилы — это бактерии, которые процветают при умеренных температурах, лучше всего растут при температуре от 20° до 45°C. Эти температуры соответствуют естественной температуре тела человека, поэтому многие патогены человека являются мезофилами. [ 15 ]
- термофилы
Выживают при температуре 45–80 °C. [ 16 ]
Кислотность
[ редактировать ]Оптимальная кислотность для бактерий обычно составляет от 6,5 до 7,0, за исключением ацидофилов . Некоторые бактерии могут изменять pH, например, выделяя кислоту, что приводит к созданию неоптимальных условий. [ 17 ]
Водная активность
[ редактировать ]Кислород
[ редактировать ]Бактерии могут быть аэробами и анаэробами . В зависимости от степени потребности в кислороде бактерии делятся на следующие классы:
- факультативные анаэробы, то есть аэротолерантное отсутствие или минимальное количество кислорода, необходимого для их роста.
- облигатные анаэробы растут только при полном отсутствии кислорода
- факультативные аэробы – могут расти как в присутствии, так и в минимальном количестве кислорода.
- облигатные аэробы – растут только в присутствии кислорода
Микронутриенты
[ редактировать ]Достаточное количество питательных веществ
Токсичные соединения
[ редактировать ]Токсичные соединения, такие как этанол, могут препятствовать росту или убивать бактерии. Это полезно для дезинфекции и консервации пищевых продуктов .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Скарстад К., Стин Х.Б., Бой Э (1983). «Параметры клеточного цикла медленно растущей Escherichia coli B/r, изученные методом проточной цитометрии» . Дж. Бактериол. 154 (2): 656–62. дои : 10.1128/jb.154.2.656-662.1983 . ПМК 217513 . ПМИД 6341358 .
- ^ Цвиеринг М.Х., Йонгенбургер И., Ромбоутс Ф.М., ван Т Рит К. (1990). «Моделирование кривой роста бактерий» . Прикладная и экологическая микробиология . 56 (6): 1875–1881. Бибкод : 1990ApEnM..56.1875Z . doi : 10.1128/aem.56.6.1875-1881.1990 . ПМК 184525 . ПМИД 16348228 .
- ^ Фанкхаузер Д.Б. (17 июля 2004 г.). «Кривая роста бактерий» . Клермонтский колледж Университета Цинциннати. Архивировано из оригинала 13 февраля 2016 года . Проверено 29 декабря 2015 г.
- ^ Дело C, Функе Б, Тортора Г (2010). Микробиология. Введение (Десятое изд.). ISBN 978-0-321-55007-1 .
- ^ Перейти обратно: а б «Рост бактерий» . Проект БАКАНОВА . Европейская комиссия. Архивировано из оригинала 24 октября 2007 года . Проверено 7 мая 2008 г.
- ^ Куттс Д.А. (15 декабря 2011 г.). «Маршалл Т. Сэвидж - экспоненциалистский взгляд» . Знаменитые экспоненциалисты . Домашняя страница экспоненциализма.
- ^ Перейти обратно: а б Бриджес Б.А., Фостер П.Л., Тиммс А.Р. (2001). «Влияние эндогенных каротиноидов на «адаптивную» мутацию Escherichia coli FC40» . Мутат. Рез . 473 (1): 109–19. дои : 10.1016/s0027-5107(00)00144-5 . ПМЦ 2929247 . ПМИД 11166030 .
- ^ Самородницкий, Дэн (5 июня 2024 г.). «Большая часть жизни на Земле находится в состоянии покоя после нажатия на «аварийный тормоз» » . Журнал Кванта . Проверено 12 июня 2024 г.
- ^ Новик А (1955). «Рост бактерий». Ежегодный обзор микробиологии . 9 : 97–110. дои : 10.1146/annurev.mi.09.100155.000525 . ПМИД 13259461 .
- ^ Анагностопулос С., Спизизен Дж. (1961). «Требования к трансформации в Bacillus Subtilis» . Дж. Бактериол . 81 (5): 741–6. дои : 10.1128/JB.81.5.741-746.1961 . ПМК 279084 . ПМИД 16561900 .
- ^ Штайнхаус Э.А., Биркеланд Дж.М. (сентябрь 1939 г.). «Исследования жизни и смерти бактерий: I. Фаза старения стареющих культур и вероятные задействованные механизмы» . Журнал бактериологии . 38 (3): 249–261. дои : 10.1128/jb.38.3.249-261.1939 . ПМЦ 374517 . ПМИД 16560248 .
- ^ Финкель С.Е., Колтер Р. (март 1999 г.). «Эволюция микробного разнообразия при длительном голодании» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (7): 4023–4027. Бибкод : 1999PNAS...96.4023F . дои : 10.1073/pnas.96.7.4023 . ПМК 22413 . ПМИД 10097156 .
- ^ Финкель С.Е. (февраль 2006 г.). «Долгосрочное выживание во время стационарной фазы: эволюция и фенотип GASP». Обзоры природы. Микробиология . 4 (2): 113–120. дои : 10.1038/nrmicro1340 . ПМИД 16415927 . S2CID 3337959 .
- ^ Мойер К.Л., Морита Р.Ю. (16 апреля 2007 г.). «Психрофилы и психротрофы». ЭЛС . дои : 10.1002/9780470015902.a0000402.pub2 . ISBN 9780470016176 .
- ^ «Мезофил» . Биологический онлайн-словарь . Проверено 6 февраля 2018 г.
- ^ Бергей Д.Х. (июль 1919 г.). «Термофильные бактерии» . Журнал бактериологии . 4 (4): 301–306. дои : 10.1128/jb.4.4.301-306.1919 . ПМЦ 378811 . ПМИД 16558843 .
- ^ Бламир Дж. «Влияние pH на скорость роста» . Бруклинский колледж . Проверено 8 октября 2016 г.
Внешние ссылки
[ редактировать ]- Исследование экспоненциального роста популяций бактерий.
- Научная помощь: ресурс для средней школы по бактериальному росту (GCSE, Alevel).
- Микробный рост, BioMineWiki. Архивировано 20 сентября 2011 г. в Wayback Machine.
- Из демонстрационного проекта Wolfram — требуется проигрыватель CDF (бесплатно) :
- Окончательное количество бактериальных клеток
- Моделирование записей подсчета микроорганизмов с помощью расширенной модели решения Ферми
- Зарождающиеся процессы роста с конкурирующими механизмами
- Модифицированный логистический изотермический коэффициент роста микроорганизмов
- Обобщенный логистический (Verhulst) изотермический рост микробов
- Рост микробной популяции, смертность и переходы между ними
- Диауктический рост бактерий на двух субстратах
Эта статья включает материал из статьи, опубликованной 26 апреля 2003 г. на Nupedia ; автор Нагина Пармар; рассмотрен и одобрен группой биологии; редактор Гайта Ланглуа; ведущий рецензент Гайта Ланглуа; ведущие редакторы, Рут Ифчер. и Ян Хогл.