Новозеландская малая короткохвостая летучая мышь
| Малая короткохвостая летучая мышь | |
|---|---|

| |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Рукокрылые |
| Семья: | Мистациниды |
| Род: | Мистацин |
| Разновидность: | М. туберкулата
|
| Биномиальное имя | |
| Мистацина туберкулатная Грей , 1843 г.
| |
| Подвиды | |
|
М. т. туберкулёзный | |
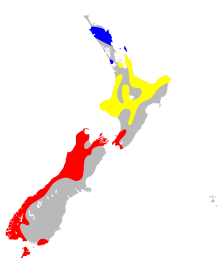
| |
| Ключ:
Диапазон М.т. опурика
Диапазон М.т. рякобия
Диапазон М.т. туберкулёзный
| |

Новозеландская малая короткохвостая летучая мышь ( Mystacina Tuberculata ) — мелкое всеядное млекопитающее, эндемичное для островов Новой Зеландии. Это один из двух существующих и трех видов наземных млекопитающих, уникальных для Новой Зеландии. Его ближайший родственник, новозеландская большая короткохвостая летучая мышь ( M.robusta ), в последний раз была замечена в 1965 году и считается вымершей из-за интенсивного нападения корабельных крыс, завезенных в последние несколько столетий. Этих летучих мышей также часто называют пекапека , их на языке маори название . Малые короткохвостые летучие мыши имеют уникальные приспособления, которые отличают их от летучих мышей, обитающих в других частях мира. Например, они вполне способны передвигаться по земле в поисках пищи, а самцы поют, чтобы привлечь партнеров, делая это по очереди. Малые короткохвостые летучие мыши являются уязвимым видом, поэтому проводятся обширные природоохранные работы и исследования, чтобы предотвратить их вымирание.
Таксономия и эволюция
[ редактировать ]
Таксономическая классификация
[ редактировать ]Mystacina Tuberculata являются членами подотряда Yangochiroptera и недавно были отнесены к надсемейству Noctillionoidea . Они являются членами семейства Mystacinidae , которое содержит единственный род Mystacina . В состав Mystacina входят два близкородственных вида: M. Tuberculata и M. Robusta . Поскольку вполне вероятно, что M.robusta вымерла, M. Tuberculata является единственным подтвержденным существующим видом в этом роде и семействе. Расположение этого вида в таксономиях и филогенетических деревьях ранее было предметом горячих дискуссий. Нынешнее положение надсемейства Noctillionoidea является результатом секвенирования и анализа ДНК. [ 2 ] Вопреки этому, некоторые ученые полагают, что физическое сходство указывает на то, что Mystacinidae относятся к надсемейству Vespertilionoidea . [ 3 ] Филогенетическое положение этого вида было выдвинуто множеством других предположений, в том числе в пределах Phyllostomidae (летучие мыши-листоносы) или Emballonuridae (летучие мыши с мешкокрылыми и влагалищными хвостами). [ 4 ] Хотя считается, что у этого вида есть подвиды, в некоторых источниках этот вид упоминается как одно целое. признает Департамент охраны природы Новой Зеландии три подвида. Они разделены по регионам, в которых каждый из них обитает; северная ( Mystacina tuberculata aupourica ), центральная ( Mystacina Tuberculata rhyacobi ) и южная ( Mystacina Tuberculata Tuberculata ). [ 5 ]
Эволюционная история
[ редактировать ]Было высказано предположение, что Mystacinidae произошли с австралийского континента. [ 6 ] и что надсемейство Noctillionoidea возникло на Гондване . [ 7 ] Когда Гондвана распалась на более мелкие континенты, члены суперсемьи оказались географически изолированы. Эта изоляция привела к миллионам лет уникальной эволюционной истории каждой семьи. Было подсчитано, что Mystacinidae впервые выделились в отдельное семейство между 68 и 35 миллионами лет назад. [ 8 ] Двадцать миллионов лет назад из этого семейства выделились два рода; Мистацина и Икаропс . Окаменелости икаропов были обнаружены по всей Австралии, что указывает на то, что ныне вымерший вид когда-то был широко распространен там. [ 8 ] Неизвестно, когда виды этого рода ( I. aenae , I. brviceps и I. paradox ) вымерли.
Мистацины мигрировали из Австралии в Новую Зеландию, поселившись на этих островах как первые млекопитающие. Судя по ископаемым свидетельствам, этот род определенно обитал в Новой Зеландии в миоцене . [ 9 ] Таким образом, мы знаем, что Mystacina выделилась в отдельный род около 20 миллионов лет назад и путешествовала из Австралии в Новую Зеландию примерно 5,3–20 миллионов лет назад. [ нужна ссылка ] Это предполагает, что Icarops не мигрировал в Новую Зеландию и что событие колонизации произошло после того, как роды стали различаться. Неясно, когда M. Tuberculata и M. Robusta стали отдельными видами.
Распространение и ареал
[ редактировать ]Малые короткохвостые летучие мыши были обнаружены в нескольких популяциях, разбросанных по всей стране. Население проживает на обоих главных островах, а также на различных прилегающих островах. В настоящее время известно несколько ключевых групп населения. Один из них находится на острове Литтл-Барьер (Хаутуру), один — на вершине Северного острова, семь — в центральной части Северного острова и один — на юго-восточной стороне Северного острова. На материковой части Южного острова их как минимум два; и один на острове Кодфиш южнее. [ 10 ] Летучие мыши могут жить на самых разных высотах: от уровня моря до высоких гор, покрытых лесом. [ 6 ] M. Tuberculata предпочитают жить в нетронутых лесах. Однако сообщалось о них в различных местах обитания, таких как сельскохозяйственные угодья и кустарники. [ 11 ]
Анатомия и физиология
[ редактировать ]Маленькие короткохвостые летучие мыши внешне похожи на своих родственников-микробатов. Они небольшого размера и обычно весят от 12 до 15 граммов (0,42–0,53 унции), когда вырастают. [ 12 ] У них относительно большие уши по сравнению с размерами головы и тела, которые обращены вперед, чтобы помочь в эхолокации. Малые короткохвостые летучие мыши также имеют относительно большие ноздри, обращенные наружу, что позволяет этим животным использовать обоняние для управления своими движениями. Их глаза маленькие, а зрение относительно слабое, что оправдывает их зависимость от звуков и обоняния. [ нужна ссылка ] Эти особенности характерны для всех рукокрылых.
Их крылья относительно меньше, чем у других видов летучих мышей. Из-за этого у них снижается ловкость, и они не могут летать так быстро. [ 13 ] Несмотря на это, они все еще способны летать; чуть меньше, чем у других видов летучих мышей. Каждая летучая мышь покрыта слоем густого меха, цвет которого варьируется от светло-коричневого до темно-коричневого. Брюшко обычно более светлого цвета, чем остальная часть тела. На заднем конце их тела расположены короткие, но сильные задние ноги, полезные при передвижении по земле. Относительно короткие короткие хвосты дали этому виду общее название. В верхней центральной части каждой передней конечности большой палец вытянут наружу. Они, как и пальцы ног, оснащены ногтями, называемыми базальными когтями. Наличие отчетливых когтей позволяет передвигаться по земле и полезно при лазании по деревьям. [ 12 ] Базальные когти не наблюдаются у других видов рукокрылых.
оцепенение
[ редактировать ]Одна из ключевых адаптаций мелких короткохвостых летучих мышей — вхождение в состояние оцепенения. Оцепенение позволяет продолжать выживание в периоды физиологического стресса, например, когда температура воздуха падает зимой. В этом состоянии летучие мыши неактивны, и работают только основные функции организма, позволяющие снизить расход энергии. Вхождение в оцепенение замедляет скорость обмена веществ в организме, но оно отличается от спячки, поскольку периоды намного короче и повторяются.
Продолжительность оцепенения обычно составляет от нескольких до нескольких часов, и летучие мыши впадают в это состояние несколько раз в течение сезона. Торпор используется в течение всего года, но особенно зимой, когда они также каждый раз входят в это состояние на более длительное время. [ 14 ] [ 15 ]
Собирательство и диета
[ редактировать ]Предпочитаемые продукты
[ редактировать ]
Новозеландские короткохвостые летучие мыши всеядны. Их диета состоит из различных продуктов питания, обычно включая насекомых, других мелких беспозвоночных, фрукты и части цветов. [ 16 ] Обычно они предпочитают есть насекомых, когда это возможно. следы моли, пауков и жуков хуху Одно исследование обнаружило в их гуано . [ 17 ] Другое исследование показало, что эти летучие мыши способны съедать половину своего веса в моли. [ 12 ] Средняя масса тела от 12 до 15 граммов (от 0,42 до 0,53 унции) соответствует 6–7,5 граммам (от 0,21 до 0,26 унции) моли, съеденной в день. Летучие мыши потребляют нектар и пыльцу, последняя из которых, как полагают, обеспечивает их белком. [ 12 ] Тем не менее, были предположения и споры о том, потребляют ли летучие мыши пыльцу намеренно или пыльца, обнаруженная в их гуано, действительно была проглочена их добычей. [ 17 ] Нектар потребляется, поскольку сахара являются хорошим и несложным источником энергии. Малые короткохвостые летучие мыши являются важным видом опылителей в Новой Зеландии за счет потребления цветов (либо путем проглатывания, либо вторично через добычу). Исследователи обнаружили, что эти летучие мыши переносят большое количество пыльцы некоторых избранных видов растений, таких как Collospermum microspermum и Dactylanthus taylorii (деревянная роза). [ 18 ] Обеспечение сохранения малых короткохвостых летучих мышей может также гарантировать сохранение видов растений, которые они опыляют.
Методы добывания пищи
[ редактировать ]Одной из определяющих особенностей новозеландских короткохвостых летучих мышей является их способность добывать пищу как в воздухе, так и на земле. Их способность добывать пищу на земле необычна для летучих мышей. Однако раннее географическое разделение Новой Зеландии около 80 миллионов лет назад [ 19 ] означало, что острова освободились от хищников. Без хищников малая короткохвостая летучая мышь эволюционировала, чтобы выжить как в воздушной, так и в наземной среде обитания. Как и другие летучие мыши, в воздухе они ловят пищу, используя эхолокацию для определения местонахождения добычи. Маленькие короткохвостые летучие мыши добывают пищу на земле, передвигаясь на конечностях и выискивая добычу с помощью обоняния. [ 16 ] У этого вида есть несколько приспособлений, которые помогают им искать пищу на земле. Например, находясь на земле, особи складывают крылья, чтобы использовать их в качестве передних конечностей — все четыре конечности передвигаются на четвероногих. Кроме того, у них есть когти, которые помогают им копаться в опавших листьях и слоях земли в поисках добычи. [ 12 ] Передвигаясь по земле, летучие мыши ходят так же, как и другие четвероногие животные. [ 20 ] Новозеландские малые короткохвостые летучие мыши обладают повышенным обонянием по сравнению с другими летучими мышами, о чем свидетельствуют их относительно большие и округлые ноздри. [ 6 ] Хотя, как и другие летучие мыши, они не слепы, они не сильно полагаются на визуальную охоту, причем гораздо более важными являются такие сигналы, как запах и звук. Собирание пищи на земле можно использовать для выкапывания насекомых и пауков, сбора цветов или опавших фруктов, а также для сбора других стационарных форм пищи. Поиск пищи в воздухе часто связан с охотой на летающих насекомых, например, мотыльков. Во время сильного дождя добыча пищи происходит редко, а во время слабого дождя активность кормодобывания немного увеличивается. [ 21 ]

Эхолокация
[ редактировать ]Как и другие летучие мыши, эти летучие мыши используют эхолокацию во время полета. [ 22 ] Малые короткохвостые летучие мыши издают звуки в диапазоне 20-40 кГц . Каждый звонок длится от семи до девяти миллисекунд. [ 23 ] Эхолокация не работает так эффективно во время поиска пищи на земле, потому что эхо может быть нарушено лесной подстилкой и всем, что занимает это пространство. Другая причина, по которой эхолокация не работает при поиске пищи на земле, заключается в том, что отраженное эхо может прервать отправку новых эхо, вызывая замешательство у животного и мешая системе работать правильно. Эта проблема не возникает, когда летучие мыши находятся в воздухе, поскольку звук проходит большее расстояние, а меньшее количество объектов может нарушить сигнал или вызвать путаницу. [ нужна ссылка ]
Жизненный цикл и социальные системы
[ редактировать ]Социальные системы и брачное поведение
[ редактировать ]Малые короткохвостые летучие мыши образуют общества деления-слияния . Это означает, что популяции будут собираться в определенное время, разделяться и вновь собираться позже. Места, где эти животные собираются вместе, называются насестами. Для ночлега они предпочитают находить деревья с большими внутренними полостями, где создают свой «дом». Место ночлега может содержать несколько деревьев в пределах одной и той же территории. Количество особей, присутствующих на местах ночлега, сильно различается: некоторые колонии состоят из сотен или тысяч отдельных летучих мышей. [ 24 ]
Самцы летучих мышей пытаются привлечь самку в весенние и летние месяцы до начала спаривания, что обычно происходит осенью (март – май в Новой Зеландии). Как и многие другие виды, самцы привлекают самок ночью с помощью голосовых криков. Самцы будут петь на одном из деревьев в колонии в течение нескольких минут. На одном и том же насесте собираются несколько самцов, по очереди поющих. [ 24 ] Такое поведение классифицируется как система размножения лек . При этом типе спаривания происходит половой отбор, когда самцы подают сигналы потенциальным самкам в надежде, что они произведут впечатление и спарятся с самцом. Разведение леков – это то, как летучие мыши-самцы «доказывают свою ценность». Последующие события спаривания немоногамны, и было замечено, что самцы спариваются с несколькими разными самками. [ 25 ] Хотя системы размножения леков наблюдаются во всем животном мире, они редки у летучих мышей и наблюдались только у одного другого вида.
Ранняя жизнь и жизненный цикл
[ редактировать ]
Детеныши малых короткохвостых летучих мышей становятся активными в течение дня после рождения. Первые два дня они безволосые, с недоразвитыми зубами, крыльями и ушами. [ 12 ] [ 26 ] Щенки способны летать примерно через четыре недели. Через шесть недель после рождения они покидают родильные дома, в которых выросли. Через несколько месяцев они достигают размеров взрослого тела. [ 6 ] Щенки рождаются летом (декабрь – февраль в Новой Зеландии). Во время летнего периода родов рассредоточенные особи перегруппируются, образуя большую колонию. Щенки сначала питаются молоком, произведенным их матерями. Каждая мать рожает одного детеныша за сезон. Результатом этого являются более высокие материнские инвестиции на одного потомка, однако немоногамная система спаривания приводит к низким отцовским инвестициям. Щенки выращиваются в специальных участках места ночлега колонии, предназначенных для материнской деятельности. Другие самки летучих мышей группируются у этих материнских насестов и помогают в выращивании детенышей. [ нужна ссылка ]
Сведений о средней продолжительности жизни малых короткохвостых летучих мышей нет. Было замечено, что другие летучие мыши подотряда Yangochiroptera живут более тридцати лет, что неожиданно для их небольших размеров тела. [ 27 ] Весной/летом летучие мыши-самцы привлекают самку. Эти пары спариваются осенью. Осенью и зимой летучие мыши разбиваются на более мелкие колонии, пока самки беременны. Затем, когда снова наступает лето, летучие мыши объединяются в большую колонию, и рождаются детеныши. К осени щенки достигли физической зрелости. Неясно, в каком возрасте они достигают половой зрелости. [ нужна ссылка ]
Воздействие человека и сохранение
[ редактировать ]Угрозы
[ редактировать ]новозеландских короткохвостых летучих мышей как уязвимых классифицирует В настоящее время МСОП , при этом количество особей со временем сокращается. [ 28 ] Появление хищников-млекопитающих оказало огромное влияние на численность этого вида. После колонизации Новой Зеландии и появления трех ключевых видов крыс численность эндемичных популяций летучих мышей сократилась. Полинезийские крысы ( R. exulans ) были одними из основных хищников летучих мышей вплоть до 19 века, когда появление черной крысы ( R. rattus ) и коричневой крысы ( R. norvegicus ) привело к усилению хищничества. [ 29 ] До того, как хищники-млекопитающие колонизировали Новую Зеландию, на малую короткохвостую летучую мышь охотились хищные птицы, такие как смеющаяся сова . [ 11 ] Еще одним распространенным хищником этих летучих мышей являются обычные домашние кошки, которые ловят летучих мышей, когда они вылетают из мест своего ночлега. Убежища летучих мышей очень уязвимы, поскольку в пределах одной области они содержат больше потенциальной добычи. Если бы эти насесты были найдены хищником, они могли бы столкнуться с массовой смертностью; В одном случае 102 меньших случая смерти короткохвостых летучих мышей были вызваны единственной домашней кошкой в центральной части Северного острова. [ 30 ]
Другой причиной их снижения является более широкое использование земель для антропогенной деятельности, такой как лесозаготовки и сельское хозяйство. Такая деятельность часто может привести к негативным последствиям для местной экосистемы, например, к фрагментации лесов . [ 31 ] Для местных животных Новой Зеландии фрагментация представляет собой угрозу, например, снижение генетического разнообразия. Однако возможно, что фрагментация леса не повлияет на мелких короткохвостых летучих мышей, поскольку было замечено, что этот вид демонстрирует поведенческую пластичность благодаря способности менять / занимать разные среды обитания. [ 25 ]
Текущие планы сохранения
[ редактировать ]Текущее состояние охраны мелких короткохвостых летучих мышей контролируется Департаментом охраны природы Новой Зеландии (DOC). План восстановления, действующий с конца 1990-х годов, включает в себя многосторонний подход, включающий различные действия, которые необходимо предпринять, чтобы предотвратить дальнейшее исчезновение этого вида. Некоторые из детальных действий включают создание новых популяций летучих мышей на различных свободных от хищников островах вокруг Новой Зеландии, обследование материка для выявления областей, требующих дополнительных усилий по сохранению, и потенциальное создание популяции в неволе. [ 32 ]
Ссылки
[ редактировать ]- ^ О'Доннелл, К. (2021). « Мистацина туберкулатная » . Красный список исчезающих видов МСОП . 2021 : e.T14261A22070543. doi : 10.2305/IUCN.UK.2021-2.RLTS.T14261A22070543.en . Проверено 22 ноября 2021 г.
- ^ Кеннеди, Мартин; Патерсон, Адриан М.; Моралес, Хуан Карлос; Парсонс, Стюарт; Виннингтон, Эндрю П.; Спенсер, Хэмиш Г. (ноябрь 1999 г.). «Длинное и короткое: длина ветвей и проблема размещения новозеландской короткохвостой летучей мыши Mystacina». Молекулярная филогенетика и эволюция . 13 (2): 405–416. дои : 10.1006/mpev.1999.0660 . ISSN 1055-7903 . ПМИД 10603267 .
- ^ Джонс-младший, Джей Нокс; Картер, Дилфорд С. (1976). «Аннотированный контрольный список с указанием подсемейств и родов. Биология летучих мышей нового мирового семейства Phyllostomidae, часть I». Лаббок: Музей Техасского технологического института. Университет . 10 :7–38. дои : 10.5962/BHL.TITLE.142603 . S2CID 83452376 .
- ^ Кирш, Джон AW; Хатчон, Джеймс М.; Бирнс, Дина, врач общей практики; Ллойд, Брайан Д. (1998). «Сродство и историческая зоогеография новозеландской короткохвостой летучей мыши, Mystacina Tuberculata Grey 1843, полученные на основе сравнения гибридизации ДНК» . Журнал эволюции млекопитающих . 5 (1): 33–64. дои : 10.1023/A:1020519019830 . S2CID 23809083 .
- ^ «Короткохвостая летучая мышь» . www.doc.govt.nz. Проверено 25 декабря 2021 г.
- ^ Перейти обратно: а б с д Картер, Джеральд Г.; Рискин, Дэниел К. (24 мая 2006 г.). «Мистацина туберкулатная» . Виды млекопитающих (790): 1–8. дои : 10.1644/790.1 . ISSN 0076-3519 . S2CID 198122536 .
- ^ Хэнд, Сюзанна Дж.; Бек, Робин, доктор медицины; Арчер, Майкл; Симмонс, Нэнси Б.; Ганнелл, Грегг Ф.; Скофилд, Р. Пол; Теннисон, Алан Джей Ди; Де Пьетри, Ванеса Л.; Солсбери, Стивен В.; Уорти, Тревор Х. (10 января 2018 г.). «Новая всеядная летучая мышь с крупным телом (Noctilionoidea: Mystacinidae) обнаруживает утраченное морфологическое и экологическое разнообразие со времен миоцена в Новой Зеландии» . Научные отчеты . 8 (1): 235. Бибкод : 2018НатСР...8..235H . doi : 10.1038/s41598-017-18403-w . ISSN 2045-2322 . ПМК 5762892 . ПМИД 29321543 .
- ^ Перейти обратно: а б Хэнд, Сюзанна; Арчер, Майкл; Годхелп, Хенк (1 мая 2005 г.). «Австралийские олиго-миоценовые мистациниды (Microchiroptera): верхний зубной ряд, новые таксоны и дивергенция новозеландских видов» . Геобиос . 38 (3): 339–352. Бибкод : 2005Geobi..38..339H . дои : 10.1016/j.geobios.2003.11.005 . ISSN 0016-6995 .
- ^ Теннисон, Алан Джей Ди (2010). «Происхождение и история наземных позвоночных Новой Зеландии». Новозеландский журнал экологии . 34 (1): 6–27. ISSN 0110-6465 . JSTOR 24060711 .
- ^ Ллойд, Брайан Д. (2003). «Демографическая история новозеландской короткохвостой летучей мыши Mystacina Tuberculata, выведенная на основе модифицированных последовательностей контрольных областей» . Молекулярная экология . 12 (7): 1895–1911. Бибкод : 2003MolEc..12.1895L . дои : 10.1046/j.1365-294X.2003.01879.x . ISSN 1365-294X . ПМИД 12803640 . S2CID 33906414 .
- ^ Перейти обратно: а б Ллойд, Б.Д. (1 марта 2001 г.). «Достижения новозеландской маммологии 1990–2000: короткохвостые летучие мыши» . Журнал Королевского общества Новой Зеландии . 31 (1): 59–81. Бибкод : 2001JRSNZ..31...59L . дои : 10.1080/03014223.2001.9517639 . ISSN 0303-6758 . S2CID 129564107 .
- ^ Перейти обратно: а б с д и ж Дэниел, MJ (1 апреля 1979 г.). «Новозеландская короткохвостая летучая мышь, Mystacina Tuberculata; обзор современных знаний» . Новозеландский журнал зоологии . 6 (2): 357–370. дои : 10.1080/03014223.1979.10428375 . ISSN 0301-4223 .
- ^ Норберг, Улла М.; Рейнер, JMV; Лайтхилл, Майкл Джеймс (16 сентября 1987 г.). «Экологическая морфология и полет летучих мышей (Mammalia; Chiroptera): адаптации крыльев, летные характеристики, стратегия кормления и эхолокация» . Философские труды Лондонского королевского общества. Б. Биологические науки . 316 (1179): 335–427. Бибкод : 1987РСТБ.316..335Н . дои : 10.1098/rstb.1987.0030 .
- ^ Чензе, Зенон Дж.; Бригам, Р. Марк; Хики, Энтони-младший; Парсонс, Стюарт (январь 2017 г.). «Холодно и одиноко? Выбор места и время года влияют на характер оцепенения у мелких короткохвостых летучих мышей» . Экология . 183 (1): 1–8. Бибкод : 2017Oecol.183....1C . дои : 10.1007/s00442-016-3707-1 . hdl : 10294/8901 . ISSN 0029-8549 . ПМИД 27561779 . S2CID 37006674 .
- ^ Гейзер, Ф.; Бригам, Р.М. (1 марта 2000 г.). «Топпор, тепловая биология и энергетика австралийских ушастых летучих мышей (Nyctophilus)». Журнал сравнительной физиологии Б. 170 (2): 153–162. дои : 10.1007/s003600050270 . ISSN 1432-136Х . ПМИД 10791575 . S2CID 5775476 .
- ^ Перейти обратно: а б Джонс, Гарет; Уэбб, Питер I.; Седжли, Джейн А.; О'Доннелл, Колин Ф.Дж. (1 декабря 2003 г.). «Таинственная Mystacina: как новозеландская короткохвостая летучая мышь (Mystacina Tuberculata) находит добычу насекомых». Журнал экспериментальной биологии . 206 (23): 4209–4216. дои : 10.1242/jeb.00678 . ISSN 0022-0949 . ПМИД 14581591 . S2CID 36430937 .
- ^ Перейти обратно: а б Дэниел, MJ (1 декабря 1976 г.). «Питание короткохвостой летучей мыши (Mystacina Tuberculata) фруктами и, возможно, нектаром». Новозеландский журнал зоологии . 3 (4): 391–398. дои : 10.1080/03014223.1976.9517927 . ISSN 0301-4223 .
- ^ Каммингс, Г.; Андерсон, С.; Деннис, Т.; Тот, К.; Парсонс, С. (2014). «Конкуренция за опыление со стороны малой короткохвостой летучей мыши и ее влияние на фенологию цветения некоторых эндемиков Новой Зеландии» . Журнал зоологии . 293 (4): 281–288. дои : 10.1111/jzo.12147 . ISSN 1469-7998 .
- ^ Гольдберг, Джулия; Тревик, Стивен А; Патерсон, Адриан М (27 октября 2008 г.). «Эволюция наземной фауны Новой Зеландии: обзор молекулярных данных» . Философские труды Королевского общества B: Биологические науки . 363 (1508): 3319–3334. дои : 10.1098/rstb.2008.0114 . ПМК 2607375 . ПМИД 18782728 .
- ^ Рискин, Дэниел К.; Парсонс, Стюарт; Шатт, Уильям А. младший; Картер, Джеральд Г.; Хермансон, Джон В. (1 мая 2006 г.). «Наземное передвижение новозеландской короткохвостой летучей мыши Mystacina Tuberculata и обыкновенной летучей мыши-вампира Desmodus rotundus» . Журнал экспериментальной биологии . 209 (9): 1725–1736. дои : 10.1242/jeb.02186 . ISSN 0022-0949 . ПМИД 16621953 . S2CID 18305396 .
- ^ Ценце, Зенон; Терли, Терция (февраль 2021 г.). «Посещение цветка дактилантуса новозеландскими короткохвостыми летучими мышами, по-видимому, зависит от ежедневных осадков» . Новозеландский журнал экологии . doi : 10.20417/nzjecol.45.8 . ISSN 0110-6465 . S2CID 231983713 .
- ^ Симмонс, Джеймс А.; Штейн, Роджер А. (1980). «Акустическая визуализация в гидролокаторе летучих мышей: сигналы эхолокации и эволюция эхолокации». Журнал сравнительной физиологии А. 135 (1): 61–84. дои : 10.1007/bf00660182 . ISSN 0340-7594 . S2CID 20515827 .
- ^ Парсонс, Стюарт (январь 1998 г.). «Влияние ситуации записи на эхолокационные крики новозеландской короткохвостой летучей мыши (Mystacina tuberculataGray)». Новозеландский журнал зоологии . 25 (2): 147–156. дои : 10.1080/03014223.1998.9518145 . ISSN 0301-4223 .
- ^ Перейти обратно: а б Кольер, К. (2020). Атрибуты социальной организации и певческое поведение самцов малой короткохвостой летучей мыши, Mystacina tuberculata (докторская диссертация, ResearchSpace @ Auckland).
- ^ Перейти обратно: а б Тот, Кори А.; Деннис, Тодд Э.; Паттемор, Дэвид Э.; Парсонс, Стюарт (2015). «Самки как мобильный ресурс: общинные насесты способствуют освоению размножения леков у летучих мышей умеренного пояса» . Поведенческая экология . 26 (4): 1156–1163. дои : 10.1093/beheco/arv070 . ISSN 1045-2249 .
- ^ Поискфилд, Д. (январь 2016 г.). «Первое разведение и ручное выращивание новозеландской короткохвостой летучей мыши Mystacina Tuberculata в Оклендском зоопарке». Международный зоопаркский ежегодник . 50 (1): 165–173. дои : 10.1111/izy.12121 . ISSN 0074-9664 .
- ^ Дамманн, Филип (октябрь 2017 г.). «Медленное старение млекопитающих. Уроки африканских землекопов и летучих мышей». Семинары по клеточной биологии и биологии развития . 70 : 154–163. дои : 10.1016/j.semcdb.2017.07.006 . ISSN 1084-9521 . ПМИД 28698112 .
- ^ О'Доннелл, К. (15 октября 2020 г.). «Мистацина туберкулатная» . Красный список исчезающих видов МСОП 2021. doi : 10.2305/iucn.uk.2021-2.rlts.t14261a22070543.en . S2CID 239146418 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Кинг, Кэролайн М.; Фостер, Стейси; Миллер, Стивен (23 июня 2011 г.). «Инвазивные европейские крысы в Великобритании и Новой Зеландии: один и тот же вид, разные результаты». Журнал зоологии . 285 (3): 172–179. дои : 10.1111/j.1469-7998.2011.00827.x . ISSN 0952-8369 .
- ^ Скримджер, Дж; Бит, А; Суонни, М. (сентябрь 2012 г.). «Кошачье хищничество короткохвостых летучих мышей (Mystacina tuberculata rhyocobia) в лесу Рангатауа, гора Руапеху, центральная часть Северного острова, Новая Зеландия» . Новозеландский журнал зоологии . 39 (3): 257–260. дои : 10.1080/03014223.2011.649770 . ISSN 0301-4223 . S2CID 84907680 .
- ^ Адеподжу, Кайоде Адевале; Салами, Аёбами Т. (25 октября 2017 г.). «Геопространственная оценка фрагментации лесов и ее последствий для экологических процессов в тропических лесах» . Журнал ландшафтной экологии . 10 (2): 19–34. doi : 10.1515/jlecol-2017-0002 . ISSN 1805-4196 . S2CID 89729777 .
- ^ Моллой, Дженис (1995). План восстановления летучих мышей (Peka peka) (Mystacina, Chalinolobus) . MJ Дэниел, Новая Зеландия. Отдел по исчезающим видам. Веллингтон, Новая Зеландия: Отдел по исчезающим видам, Департамент охраны природы. ISBN 0-478-01570-4 . OCLC 154583004 .