Ромбическая губа
Ромбическая губа представляет собой заднюю часть развивающегося метэнцефалона , которую можно временно распознать внутри эмбриона позвоночного. Он простирается назад от крыши четвертого желудочка до дорсальных нейроэпителиальных клеток. [1] Ромбическую губу можно разделить на восемь структурных единиц на основе ромбомеров 1–8 (r1–r8), которые можно распознать на ранних стадиях развития заднего мозга. [2] Производя гранулярные клетки и пять ядер ствола мозга, ромбическая губа играет важную роль в развитии сложной нервной системы мозжечка. [3]

История
[ редактировать ]В ходе исследований человеческих эмбрионов, проведенных в конце 1890-х годов, швейцарский анатом Вильгельм Хис идентифицировал часть нейроэпителия заднего мозга, которая отличалась от остальной части нейроэпителия заднего мозга по своей морфологии, устойчивому делению хромосом на поздних стадиях эмбриогенеза и расположению потоков нейронов. через периферию заднего мозга. Он назвал эту зону «раутенлиппе», или ромбическая губа, потому что казалось, что она исходит из ромбовидного отверстия четвертого желудочка, очень напоминая форму рта. Он предположил, что ромбическая губа содержит прецеребеллярные предшественники, которые мигрируют вентрально, чтобы заселить ядра моста и оливы, но доступные в то время методологии ограничивали количество доказательств, которые он мог собрать. Первое реальное доказательство того, что прецеребеллярные нейроны имеют дорсальное происхождение, было получено в 1990-х годах с помощью химер кур-перепелов — метода, при котором части нейроэпителия заднего мозга перепела прививаются куриным эмбрионам in ovo. . Отслеживание вентральной миграции дочерних клеток перепела подтвердило теорию Хиса. [4]
Разработка
[ редактировать ]| Метэнцефалон | |
|---|---|
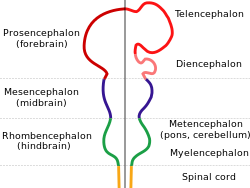 Схема, изображающая основные подразделения эмбрионального мозга позвоночных. Эти области позже дифференцируются в переднего мозга , среднего мозга и заднего мозга . структуры | |
 | |
| Идентификаторы | |
| НейроЛекс ID | бирнлекс_965 |
| Анатомические термины нейроанатомии | |
Происхождение мозжечковой территории
[ редактировать ]- Мозжечок районе , или «маленький мозг», представляет собой часть мозга, прикрепленную к стволу мозга в моста . Помимо других функций, он важен для баланса, координации движений и поддержания мышечного тонуса. [5] Внешний слой мозжечка, кора, состоит из трех слоев, содержащих два класса нейронов. Один из этих классов был идентифицирован как гранулярные клетки , которые производятся ромбической губой. [6]
- Гены FGF8 и EN1 играют важную роль в формировании паттерна областей среднего и заднего мозга. [7] В частности, исследования показали, что разные дозы FGF8, по-видимому, определяют тектум или мозжечок: области, подвергшиеся воздействию высоких доз FGF8, приобретают мозжечковые характеристики, в то время как области, подвергшиеся воздействию слабых доз, становятся тектумом. [8]
Клеточный вклад ромбической губы
[ редактировать ]- Нервная спецификация мозжечка начинается с симметричного выступа в четвертом желудочке . [7] Примерно с девятого по одиннадцатый день эмбрионального развития клетки возникают из двух областей: ромбической губы и желудочковой зоны. Ромбическая губа производит нейроны глутаматергических гранул (которые развиваются из r1) и пять ядер ствола мозга (считается, что они развиваются из r6-r8). Одно ядро ствола мозга, нижнее оливковое ядро, выбрасывает лазящие волокна для иннервации клеток Пуркинье. Остальные четыре ядра (наружное клиновидное ядро, латеральное ретикулярное ядро, мостовое ядро и таламическое ретикулярное ядро) проецируют мшистые волокна для иннервации гранулярных нейронов. [2] [4] [7] Тем временем клетки желудочковой зоны эволюционируют в ГАМКергические клетки Пуркинье (другой тип нейронов мозжечка) и глубокие ядра мозжечка . [8]
Ромбическое развитие губ
[ редактировать ]- Расположенная между четвертым желудочком и верхней пластинкой, ромбическая губа простирается от r1 до r8 и может быть разделена на верхнюю, или мозжечковую, и нижнюю, или заднюю части мозга, поскольку ствол мозга изгибается во время более позднего развития. [2] [7] Экспрессия Math1 , транскрипционного фактора с базовой структурой спираль-петля-спираль , управляет зародышевым эпителием ромбической губы и экспрессируется в областях среднего и заднего мозга уже на 9.5 день эмбриона (E9.5). [7] [9] Было показано, что у мышей с нулевым значением Math 1 отсутствует несколько производных ромбических губ, включая гранулярные нейроны мозжечка и понтинное ядро прецеребеллярной системы. [7] Ромбическая губа организована в соответствии с временной картой судеб, в которой предшественники, экспрессирующие Math1 , которые сначала эмигрируют из зародышевого слоя (до E12.5), становятся глубокими ядерными нейронами, тогда как те, которые выходят позже (E13), становятся гранулярными клетками. [8] Эксперименты с химерами кур и перепелов показали, что r1 является наиболее вероятным источником предшественников гранулярных клеток. Внутри ромбической губы предшественники гранулярных клеток делятся и приобретают униполярную морфологию с единственным отростком, направленным к месту назначения. [7]
- По мере продолжения развития клетки-предшественники гранул, которые возникли в RL, генерируют внешний слой гранул (EGL). Это движение происходит между E15 и P15 (эмбриональная стадия 15 и постнатальная стадия 15). ЭГЛ представляет собой вторичный герминативный эпителий, охватывающий весь мозжечок. [8] Этот период является важным этапом развития мозжечка. Экспансия клеток-предшественников EGL создает большую популяцию нейронов, численность которой превышает численность клеток Пуркинье в 250:1 во взрослой коре мозжечка. [6] Нейроны-гранулы EGL экспрессируют гены, которые играют специфическую и решающую роль в пролиферации клеток. Эти гены называются Math1, RU49/Zipro1 и Zic1. [7] Как упоминалось ранее, Math1 выражается на ранних стадиях развития RL. Эксперименты по нокауту с участием RU49/Zipro1 приводят к очень небольшим изменениям в структуре мозга, но сверхэкспрессия этих генов приводит к резкому увеличению количества гранулярных клеток и пролиферации внешнего EGL. [7] Этот результат позволяет предположить, что ген RU49/Zipro1 играет достаточную роль в пролиферации клеток.
- В районе P15 пролиферация гранулярных клеток требует взаимодействия с клетками Пуркинье, типом нейронов мозжечка, характеризующимся большим и разветвленным дендритным ветвлением. [8] Эти клетки выпускают звуковой еж (Тсс); который представляет собой белок, который контролирует дальнейшую пролиферацию предшественников гранулярных клеток. [7] Это означает, что на стадии P15 пролиферация предшественников гранулярных клеток контролируется генами, которые они транскрибируют (RU49/Zipro1), а также продуктами соединительных клеток (Shh).
- Как только EGL полностью сформирован, предшественники гранулярных клеток мигрируют внутрь, образуя слой, называемый внутренним слоем гранул. Непосредственно перед миграцией клетки претерпевают ряд электрофизиологических изменений. Считается, что NMDA-рецепторы активируются для подготовки клетки к дальнейшей миграции. [7] Активация этих NMDA-рецепторов вызывает деполяризацию клеток. Чтобы сбалансировать эту деполяризацию, активируется канал GIRK2. Он участвует в гиперполяризации клетки после того, как рецепторы NMDA деполяризуют клетку. [7] Пока происходят эти изменения, клетки также направляются в этот новый слой радиальными глиальными клетками. Попав в IGL, они перестают делиться и больше не экспрессируют ген Math1.
- Заключительная стадия созревания гранулярных клеток происходит в IGL. На этой стадии гранулярные клетки экспрессируют другой набор зрелых рецепторов, включая рецепторы GC5 и ГАМК . Новые клетки также вступают в контакт с клетками-зернами. Мшистые волокна, которые в основном встречаются в прецеребеллярных ядрах, вступают в контакт с гранулярными клетками в IGL. Нейроны-гранулы также расширяются, чтобы вступить в контакт с клетками Гольджи . [7]
Ссылки
[ редактировать ]- ^ Гилторп Дж.Д., Папантониу Э.К., Чедоталь А., Ламсден А., Вингейт Р.Дж. (октябрь 2002 г.). «Миграция производных ромбических губ мозжечка» . Разработка . 129 (20): 4719–28. ПМИД 12361964 .
- ^ Jump up to: а б с Дун XP (август 2012 г.). «Происхождение нейронов лазающих волокон и определение ромбической губы». Международный журнал нейробиологии развития . 30 (5): 391–5. дои : 10.1016/j.ijdevneu.2012.02.002 . ПМИД 22406199 .
- ^ Вингейт Р.Дж. (февраль 2001 г.). «Ромбическая губа и раннее развитие мозжечка». Современное мнение в нейробиологии . 11 (1): 82–8. дои : 10.1016/S0959-4388(00)00177-X . ПМИД 11179876 .
- ^ Jump up to: а б Рэй Р.С., Дымецки С.М. (декабрь 2009 г.). «Rautenlippe Redux — к единому взгляду на прецеребеллярную ромбическую губу» . Современное мнение в области клеточной биологии . 21 (6): 741–7. дои : 10.1016/j.ceb.2009.10.003 . ПМК 3729404 . ПМИД 19883998 .
- ^ Род, Сили (2012). Анатомия и физиология Сили . Нью-Йорк, штат Нью-Йорк: МакГроу Хилл. п. 1152. ИСБН 978-0-07-352561-7 .
- ^ Jump up to: а б Хаттен М.Е., Хайнц Н. (1995). «Механизмы формирования и спецификации нейронных паттернов в развивающемся мозжечке». Ежегодный обзор неврологии . 18 : 385–408. дои : 10.1146/annurev.ne.18.030195.002125 . ПМИД 7605067 .
- ^ Jump up to: а б с д и ж г час я дж к л м Ван В.Ю., Зогби Х.Ю. (июль 2001 г.). «Генетическая регуляция развития мозжечка». Обзоры природы Неврология . 2 (7): 484–91. дои : 10.1038/35081558 . ПМИД 11433373 .
- ^ Jump up to: а б с д и Карлетти Б., Росси Ф. (февраль 2008 г.). «Нейрогенез в мозжечке». Нейробиолог . 14 (1): 91–100. дои : 10.1177/1073858407304629 . ПМИД 17911211 .
- ^ Роуэн, Элисон (2005). «Тайна математики» . Обзоры природы Неврология . 6 (12): 916. дои : 10.1038/nrn1813 .
Дальнейшее чтение
[ редактировать ]- Роуэн, Элисон (2006). «Развитие: картирование развития мозжечка» . Обзоры природы Неврология . 7 (8): 598. doi : 10.1038/nrn1982 .