Трогоцитоз
Трогоцитоз ( греч . trogo ; грызть ) — это когда клетка поедает другую клетку. [1] Это процесс, при котором лимфоциты ( В- , Т- и NK- клетки ), конъюгированные с антигенпрезентирующими клетками, извлекают поверхностные молекулы из этих клеток и экспрессируют их на своей собственной поверхности. [2] Молекулярная реорганизация, происходящая на границе раздела лимфоцита и антигенпредставляющей клетки во время конъюгации, также называется « иммунологическим синапсом ».
Открытие
[ редактировать ]Первые признаки существования этого процесса датируются концом 70-х годов, когда несколько исследовательских групп сообщили о присутствии неожиданных молекул, таких как молекулы главного комплекса гистосовместимости (MHC), на Т-клетках. Представление о том, что фрагменты мембраны, а не изолированные молекулы, могут захватываться Т-клетками на антигенпрезентирующих клетках, было предложено захватом молекул MHC, слитых с зеленым флуоресцентным белком (GFP) в их внутриклеточной части. [3] Демонстрация участия фрагментов мембраны в этом процессе переноса произошла, когда было обнаружено, что флуоресцентные зонды, встроенные в плазматическую мембрану антигенпрезентирующей клетки, а также молекулы, не относящиеся к MHC, захватываются Т-клетками вместе с антигеном. [4] [5]
Типы ячеек
[ редактировать ]Трогоцитоз первоначально был зарегистрирован в Т-, В- и NK-клетках как in vivo , так и in vitro . На Т- и В-клетках трогоцитоз запускается, когда рецептор Т-клеток (TCR) на Т-клетках или рецептор В-клеток (BCR) на В-клетках взаимодействует с антигеном, распознаваемым на антигенпрезентирующих клетках. Как и в лимфоцитах, трогоцитоз возникает при ПМН (полиморфно-ядерных лейкоцитах , также известных как гранулоциты) и связан с эффективной ADCC (антитело-зависимой клеточно-опосредованной цитотоксичностью).
Было показано, что для инициации ADCC in vitro PMN должны прикрепляться к своим клеткам-мишеням и образовывать плотные соединения с опсонизированными антителами опухолевыми клетками. Эта кластеризация клеток предшествует взаимному мембранному обмену липидов между эффекторной клеткой и клеткой-мишенью во время ADCC и не происходит в отсутствие опсонизирующих антител. [6] Трогоцитоз также встречается в моноцитах и дендритных клетках. За пределами иммунной системы аналогичный перенос фрагментов мембран был зарегистрирован между сперматозоидами и ооцитами, и считается, что этот процесс способствует слиянию гамет. [7]
В последнее время этот термин стали приписывать макрофагам, таким как резидентная микроглия ЦНС, которые способны частично удалять небольшие части нейрональных аксонов во время постнатального развития. [8]
Механизм действия
[ редактировать ]Трогоцитоз включает перенос фрагментов плазматической мембраны из представляющей клетки в лимфоцит. Трогоцитоз специфически запускается передачей сигналов антигенного рецептора на Т- и В-клетках, рецептором, ингибирующим и активирующим киллер, на NK-клетках, а также различными рецепторами на других клетках, включая рецептор Fc и рецептор класса А-поглотителя. Вполне вероятно, что трогоцитоз не включает захват везикул, таких как экзосомы, секретируемых антигенпрезентирующими клетками. Скорее, молекулы могут перемещаться от антигенпрезентирующих клеток к лимфоцитам, переносясь мембранными нанотрубками, или фрагменты мембраны могут быть разорваны Т-клетками из-за физических сил, необходимых для образования и деформации иммунологических синапсов. В зависимости от двух типов клеток, участвующих в конъюгатах, трогоцитоз может быть однонаправленным или двунаправленным. Белков, переносимых посредством трогоцитоза, много, и в основном они включают белки, встроенные в плазматическую мембрану или тесно связанные с ней (белки, охватывающие липидный бислой или встроенные во внеклеточные или внутриклеточные листки). Например, недавно было показано, что человеческие лимфоциты приобретают внутренней мембраны белок H-Ras , G-белок, жизненно важный для общих функций лимфоцитов и важный участник рака человека , из клеток, которые они сканируют. [9] Перенос зависел от клеточного контакта и происходил в контексте образования клеточного конъюгата. Более того, приобретение онкогенного H-RasG12V NK- и Т-лимфоцитами имело важные биологические функции при принятии лимфоцитов: перенесенный H-RasG12V индуцировал ERK фосфорилирование , увеличивал интерферона-γ и фактора некроза опухоли-α секрецию , усиливал пролиферацию лимфоцитов и усиленное уничтожение клеток-мишеней, опосредованное NK.
Физиологические последствия
[ редактировать ]Трогоцитоз может иметь физиологические последствия двумя способами: либо потому, что клетки-реципиенты приобретают и используют молекулы, которые они обычно не экспрессируют, либо потому, что клетки-доноры лишаются молекул, что может изменить их взаимодействие с клеточными партнерами. Приобретенные молекулы, такие как регуляторные молекулы с внеклеточными или внутриклеточными компонентами, могут изменять активность лимфоцитов и направлять некоторые функции лимфоцитов, такие как миграция к адекватным поврежденным тканям . Такие полученные фрагменты плазматической мембраны также могут способствовать способности к пролиферации , поскольку липиды являются высокоэнергетическими компонентами , требующими установления. Трогоцитоз, возможно, впервые появился у очень примитивных организмов, питающихся другими клетками.Большинство биологических функций, выявленных при трогоцитозе, наблюдаются у лимфоцитов и дендритных клеток. Основные выводы в этом направлении таковы:
- цитотоксические Т-лимфоциты, захватившие комплексы антигенный пептид-MHC, могут быть уничтожены CTL, специфичными для этого антигена (процесс, называемый братоубийством ).
- Т-хелперы, захватившие комплексы антигенный пептид-MHC, участвуют в цикле отрицательной регуляторной обратной связи, приводящем к их инактивации. [10]
- дендритные клетки, лишенные комплексов антигенный пептид-MHC Т-клетками посредством трогоцитоза, способствуют созреванию аффинности ответа Т-клеток путем отбора Т-клеток с высоким сродством [11]
- понижающая модуляция костимулирующих молекул на дендритных клетках, опосредованная Т-клетками, приводит к регуляции ответа Т-клеток [12]
- перенос антигена между дендритными клетками путем трогоцитоза способствует реактивации Т-клеток памяти за счет наивных Т-клеток. [13]
- перенос антигена между дендритными клетками путем трогоцитоза способствует отторжению аллотрансплантата. [14]
Приложения
[ редактировать ]В серотерапии
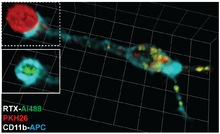
[ редактировать ]Терапевтические антитела можно использовать для лечения рака. Примером может служить ритуксимаб , терапевтическое антитело, используемое для лечения хронического лимфоцитарного лейкоза , распознающее молекулу CD20, экспрессируемую опухолевыми клетками, и приводит к их элиминации. [15] Однако использование слишком большого количества антител частично приводит к удалению комплексов ритуксимаб-CD20 с поверхности опухолевых клеток моноцитами посредством трогоцитоза. Этот эффект приводит к выходу опухолевых клеток за счет антигенной модуляции . Снижение дозы терапевтических антител для ограничения степени трогоцитоза может улучшить их терапевтическую эффективность. [16]
Эпратузумаб (Mab CD22) действует с помощью трогоцитоза для переноса CD22 и других B-клеточных белков из B-клеток в эффекторные клетки. [17]
В качестве инструментов иммуномониторинга
[ редактировать ]Анализы TRAP (протокол анализа TRogocytosis) позволяют идентифицировать, характеризовать и очищать Т- и B-клетки, распознающие их специфический антиген, на основе их способности извлекать молекулы (в данном случае флуоресцентные зонды) из плазматической мембраны антигенпрезентирующих клеток. [18] Эти анализы требуют такого оборудования, как проточный цитометр, но в остальном они очень дешевы, просты в выполнении, быстры (можно выполнить в течение 3 часов) и применимы к любой популяции Т- или В-клеток. Анализы TRAP успешно используются для обнаружения реакции Т-клеток на вирусные инфекции. [19] рак, [20] аутоиммунные заболевания [21] и вакцины. [22]
См. также
[ редактировать ]Процесс трогоцитоза считается отличным от несвязанных процессов, известных как фагоцитоз и парацитофагия, но похожим на них .
Ссылки
[ редактировать ]- ^ Танцуй, Эмбер (2019). «Основная концепция: клетки поедают друг друга посредством недооцененного процесса трогоцитоза» . Труды Национальной академии наук . 116 (36): 17608–17610. дои : 10.1073/pnas.1912252116 . ПМК 6731757 . ПМИД 31481628 .
- ^ Жоли, Этьен; Худрисье, Денис (сентябрь 2003 г.). «Что такое трогоцитоз и какова его цель?» . Природная иммунология . 4 (9): 815. дои : 10.1038/ni0903-815 . ПМИД 12942076 .
- ^ Хуанг, Дж.; Ян, Ю; Сепульведа, Х; Ши, Вт; Хван, я; Петерсон, Пенсильвания; Джексон, MR; Срент, Дж; Цай, З. (29 октября 1999 г.). «TCR-опосредованная интернализация комплексов пептид-MHC, приобретенных Т-клетками». Наука . 286 (5441): 952–954. дои : 10.1126/science.286.5441.952 . ПМИД 10542149 .
- ^ Патель, Дхавал М.; Арнольд, Паула Ю.; Уайт, Грегори А.; Нарделла, Джон П.; Мэнни, Марк Д. (15 ноября 1999 г.). «Комплексы MHC/пептид класса II высвобождаются из APC и приобретаются Т-клеточными ответчиками во время специфического распознавания антигена» . Журнал иммунологии . 163 (10): 5201–5210. дои : 10.4049/jimmunol.163.10.5201 . ПМИД 10553040 . S2CID 38736928 .
- ^ Худрисье, Денис; Рионд, Джоэль; Мазаргиль, Оноре; Гайрин, Жан Эдуард; Жоли, Этьен (15 марта 2001 г.). «Авангард: CTL быстро захватывают фрагменты мембраны из клеток-мишеней в зависимости от передачи сигналов TCR» . Журнал иммунологии . 166 (6): 3645–3649. дои : 10.4049/jimmunol.166.6.3645 . ПМИД 11238601 .
- ^ Хорнер, Хайке; Фрэнк, Кэрола; Дечант, Клаудия; Репп, Роланд; Гленни, Мартин; Херрманн, Мартин; Штокмейер, Бернхард (1 июля 2007 г.). «Образование интимных клеточных конъюгатов и обмен мембранных липидов предшествуют индукции апоптоза в клетках-мишенях во время антителозависимой гранулоцитарно-опосредованной цитотоксичности» . Журнал иммунологии . 179 (1): 337–345. дои : 10.4049/jimmunol.179.1.337 . ПМИД 17579054 .
- ^ Барро-Ланге, Вирджиния; Науд-Барриант, Натали; Бомзель, Морган; Вольф, Жан-Филипп; Зийят, Ахмед (15 июня 2007 г.). «Перенос фрагментов мембран ооцита в оплодотворяющие сперматозоиды» . Журнал ФАСЭБ . 21 (13): 3446–3449. дои : 10.1096/fj.06-8035hyp . ПМИД 17575263 . S2CID 35630271 .
- ^ Вайнхард, Летиция; ди Бартоломеи, Джулия; Боласко, Джулия; Мачадо, Педро; Шибер, Николь Л.; Нениските, Урте; Эксига, Мелани; Вадисюте, Огюст; Раджоли, Анджело; Шертель, Андреас; Шваб, Янник; Гросс, Корнелиус Т. (26 марта 2018 г.). «Микроглия реконструирует синапсы путем пресинаптического трогоцитоза и индукции филоподий головки позвоночника» . Природные коммуникации . 9 (1): 1228. Бибкод : 2018NatCo...9.1228W . дои : 10.1038/s41467-018-03566-5 . ПМЦ 5964317 . ПМИД 29581545 .
- ^ Рехави, Одед; Гольдштейн, Итамар; Верницкий, Хелли; Ротблат, Барак; Клоог, Йоэль; Канеллопулос, Жан (21 ноября 2007 г.). «Межклеточный перенос онкогенного H-Ras в иммунологическом синапсе» . ПЛОС ОДИН . 2 (11): е1204. Бибкод : 2007PLoSO...2.1204R . дои : 10.1371/journal.pone.0001204 . ПМК 2065899 . ПМИД 18030338 .
- ^ Хелфт, Джули; Жаке, Александра; Джонкер, Натали Т.; Гранжан, Изабель; Дороти, Гийом; Киссенпфенниг, Адриан; Малиссен, Бернар; Матцингер, Полли; Ланц, Оливье (15 августа 2008 г.). «Антигенспецифические взаимодействия ТТ регулируют экспансию Т-клеток CD4» . Кровь . 112 (4): 1249–1258. дои : 10.1182/blood-2007-09-114389 . ПМК 2515122 . ПМИД 18539897 .
- ^ Кедл, Росс М.; Шефер, Брайан С.; Капплер, Джон В.; Маррак, Филиппа (3 декабря 2001 г.). «Т-клетки подавляют комплексы пептид-MHC на APC in vivo». Природная иммунология . 3 (1): 27–32. дои : 10.1038/ni742 . ПМИД 11731800 . S2CID 20735730 .
- ^ Куреши, Омар С.; Чжэн, Юн; Накамура, Кёко; Аттридж, Кесли; Манзотти, Клэр; Шмидт, Эмили М.; Бейкер, Дженнифер; Джеффри, Луиза Э.; Каур, Сатдип; Бриггс, Зоя; Хоу, Ти З.; Футтер, Клэр Э.; Андерсон, Грэм; Уокер, Люси СК; Сансом, Дэвид М. (29 апреля 2011 г.). «Транс-эндоцитоз CD80 и CD86: молекулярная основа клеточной внешней функции CTLA-4» . Наука . 332 (6029): 600–603. Бибкод : 2011Sci...332..600Q . дои : 10.1126/science.1202947 . ПМК 3198051 . ПМИД 21474713 .
- ^ Оаким, Линда М.; Беван, Майкл Дж. (31 марта 2011 г.). «Перекрестные дендритные клетки стимулируют активацию CD8+ Т-клеток памяти после вирусной инфекции» . Природа . 471 (7340): 629–632. Бибкод : 2011Natur.471..629W . дои : 10.1038/nature09863 . ПМЦ 3423191 . ПМИД 21455179 .
- ^ Эррера, Оскель Баррозу; Гольшаян, Дела; Тибботт, Ребекка; Очоа, Франсиско Сальсидо; Джеймс, Марта Дж.; Марелли-Берг, Федерика М.; Лехлер, Роберт И. (15 октября 2004 г.). «Новый путь презентации аллоантигена дендритными клетками» . Журнал иммунологии . 173 (8): 4828–4837. дои : 10.4049/jimmunol.173.8.4828 . ПМИД 15470023 .
- ^ Бом, Пол В.; Кеннеди, Адам Д.; Уильямс, Майкл Э.; Линдорфер, Маргарет А.; Тейлор, Рональд П. (15 февраля 2006 г.). «Реакция бритья: комплексы ритуксимаб/CD20 удаляются из клеток мантийной лимфомы и клеток хронического лимфоцитарного лейкоза моноцитами THP-1» . Журнал иммунологии . 176 (4): 2600–2609. дои : 10.4049/jimmunol.176.4.2600 . ПМИД 16456022 .
- ^ Уильямс, Майкл Э.; Денсмор, Джон Дж.; Павлючкович, Эндрю В.; Бом, Пол В.; Кеннеди, Адам Д.; Линдорфер, Маргарет А.; Хэмил, Сьюзен Х.; Эгглтон, Джейн С.; Тейлор, Рональд П. (15 ноября 2006 г.). «Низкие дозы ритуксимаба трижды в неделю уменьшают потерю CD20 при бритье и способствуют более эффективному нацеливанию при хроническом лимфоцитарном лейкозе» . Журнал иммунологии . 177 (10): 7435–7443. дои : 10.4049/jimmunol.177.10.7435 . ПМИД 17082663 .
- ^ «Эпратузумаб» . Иммуномедики .
- ^ Добеф, Сандрин; Пуо, Анн-Лора; Жоли, Этьен; Худрисье, Денис (29 декабря 2006 г.). «Простой метод, основанный на трогоцитозе, для обнаружения, количественного определения, характеристики и очистки антигенспецифических живых лимфоцитов с помощью проточной цитометрии путем захвата ими фрагментов мембран из антигенпрезентирующих клеток». Протоколы природы . 1 (6): 2536–2542. дои : 10.1038/нпрот.2006.400 . ПМИД 17406507 . S2CID 25090649 .
- ^ Бидлинг, Кэрол; Слифка, Марк К. (1 октября 2006 г.). «Количественная оценка жизнеспособных вирусспецифичных Т-клеток без предварительного знания точной эпитопной специфичности». Природная медицина . 12 (10): 1208–1212. дои : 10.1038/nm1413 . ПМИД 17013384 . S2CID 9102979 .
- ^ Махленкин, Артур; Узана, Ронни; Франкенбург, Шошана; Айзенберг, Галит; Айзенбах, Леа; Питцовски, Джейкоб; Городецкий, Рафаэль; Ниссан, Авирам; Перец, Тамар; Лотем, Михал (15 марта 2008 г.). «Захват мембран опухолевых клеток путем трогоцитоза облегчает обнаружение и выделение опухолеспецифичных функциональных ЦТЛ» . Исследования рака . 68 (6): 2006–2013. дои : 10.1158/0008-5472.CAN-07-3119 . ПМИД 18339883 .
- ^ Бахбуи, Бучаиб; Петтре, Сеголен; Бертло, Лорелин; Гарсия, Александра; Элонг Нгоно, Энни; Дегок, Николя; Мишель, Лаура; Вертлевски, Сандрин; Лефрер, Фабьен; Мейниэль, Клэр; Делькруа, Катрин; Бруар, Софи; Лаплауд, Давид-Аксель; Суиллу, Жан-Поль (июнь 2010 г.). «Распознавание Т-клетками аутоантигенпредставляющих клеток с помощью анализа переноса белка выявляет высокую частоту антимиелиновых Т-клеток при рассеянном склерозе» . Мозг . 133 (6): 1622–1636. дои : 10.1093/brain/awq074 . ПМИД 20435630 .
- ^ Добеф, Сандрин; Превиль, Ксавье; Момот, Мари; Миссери, Иоланда; Жоли, Этьен; Худрисье, Денис (сентябрь 2009 г.). «Улучшение схем введения вакцин на основе CyaA с использованием анализов TRAP для обнаружения антигенспецифических CD8 + Т-клеток непосредственно ex vivo». Вакцина . 27 (41): 5565–5573. doi : 10.1016/j.vaccine.2009.07.035 . ПМИД 19647811 .