Трофическая когерентность
Трофическая когерентность — свойство ориентированных графов (или направленных сетей ). [1] В его основе лежит концепция трофических уровней, используемая главным образом в экологии . [2] но который может быть определен для направленных сетей в целом и обеспечивает определенную иерархическую структуру среди узлов. Трофическая когерентность — это тенденция узлов попадать на четко определенные трофические уровни. Это связано с несколькими структурными и динамическими свойствами направленных сетей, включая преобладание циклов. [3] и сетевые мотивы , [4] экологическая стабильность , [1] интервал, [5] и распространение таких процессов, как эпидемии и нейронные лавины . [6]
Определение
[ редактировать ]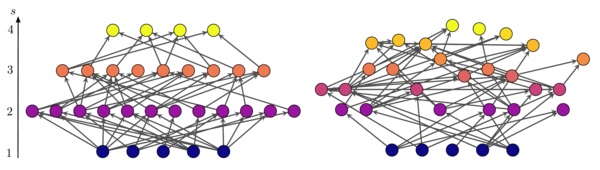
Рассмотрим направленную сеть, определяемую формулой тот матрица смежности .Каждый узел можно присвоить трофический уровень в соответствии с





где является в степени и узлах с (базальные узлы) имеют по соглашению.С каждым ребром связано трофическое различие , определяемое как .Трофическая связность сети является мерой того, насколько четко распределены трофические расстояния. , находится вокруг своего среднего значения, которое всегда .Это можно отразить с помощью параметра некогерентности. , равно стандартному отклонению :








где количество ребер в сети. [1]

На рисунке показаны две сети, различающиеся трофической связностью. Положение узлов на вертикальной оси соответствует их трофическому уровню. В сети слева узлы попадают в отдельные (целые) трофические уровни, поэтому сеть максимально связна. . На рисунке справа многие узлы имеют дробные трофические уровни, а сеть более бессвязна. . [6]


Трофическая когерентность в природе
[ редактировать ]Степень, в которой эмпирические сети трофически связны (или некогерентны), можно исследовать путем сравнения с нулевой моделью.Это обеспечивается базальным ансамблем , состоящим из сетей, в которых все небазальные узлы имеют одинаковую долю базальных узлов.для соседей. [3] Ожидаемые значения в этом ансамбле сходятся к значениям широко используемого конфигурационного ансамбля. [7] в пределе , (с и количество узлов и ребер), и может быть показано численно как хорошее приближение для конечных случайных сетей. Базальное ожидание ансамбля для параметра некогерентности равно





где — количество ребер, соединенных с базальными узлами. [3] Соотношение измеренные в эмпирических сетях, показывают, являются ли они более или менее когерентными, чем случайное ожидание.Например, Джонсон и Джонс [3] обнаружить в наборе сетей, что пищевые сети в значительной степени согласованы метаболические сети существенно некогерентны , а сети регуляции генов близки к случайному ожиданию .





Трофические уровни и функция узла
[ редактировать ]Пока еще мало понимания механизмов, которые могут привести к тому, что определенные виды сетей станут значительно более уязвимыми. когерентный или некогерентный. [3] Однако в системах, которые демонстрируют корреляцию между трофическим уровнем и другими особенностями узлов,процессы, которые способствовали созданию ребер между узлами с определенными характеристиками, могли вызвать когерентность или некогерентность.В случае пищевых сетей хищники, как правило, специализируются на потреблении добычи с определенными биологическими свойствами (такими как размер, скорость или поведение). которые коррелируют с их рационом и, следовательно, с трофическим уровнем. Это было предложено как причина согласованности пищевой сети. [1] Однако модели пищевой сети, основанные на оси ниш, не воспроизводят реалистичную трофическую когерентность. [1] что может означать, что либо этого объяснения недостаточно, либо несколько нишевых аспектов . необходимо учитывать [8]

Связь между трофическим уровнем и функцией узла можно увидеть в сетях, отличных от пищевых сетей. На рисунке показана сеть смежности слов Зеленые яйца и ветчина» . , взятая из книги доктора Сьюза « [3] Высота узлов представляет их трофические уровни (в соответствии с направлением края, которое противоположно направлению, указанному стрелками, которые указывают порядок, в котором слова объединяются в предложения). Синтаксическая функция слов также отображается цветом узла. Существует четкая связь между синтаксической функцией и трофическим уровнем: средний трофический уровень нарицательных существительных (синий) , тогда как у глаголов (красный) .Этот пример иллюстрирует, как трофическая когерентность или некогерентность могут возникать из функции узла, а также то, что трофическая структура сетей обеспечивает средства идентификации функции узла в определенных системах.


Создание трофически когерентных сетей
[ редактировать ]Существуют различные способы создания направленных сетей с заданной трофической когерентностью, все они основаны на постепенном введении новых ребер. к системе таким образом, что вероятность принятия каждого нового ребра-кандидата зависит от ожидаемой трофической разницы, которую оно будет иметь.
Модель предпочтительной добычи представляет собой развивающуюся сетевую модель, аналогичную модели предпочтительной привязанности Барабаси-Альберта , но вдохновленную экосистемой, которая растет за счет иммиграции новых видов. [1] Один начинается с базальных узлов и приступает к введению новых узлов в общей сложности .Каждый новый узел назначается первый сосед (вид-жертва в контексте пищевой сети) и новый край расположен от к . Новому узлу присваивается временный трофический уровень. .Затем еще новые соседи выбраны для из числа входящих в сеть по их трофическим уровням. В частности, для нового кандидата в соседнем , вероятность быть выбранным является функцией . Джонсон и др. [1] использовать







где является параметром, регулирующим трофическую когерентность: для генерируются максимально когерентные сети, и монотонно возрастает с для .Выбор является произвольным. Одна из возможностей состоит в том, чтобы установлен на ,где это количество узлов, уже находящихся в сети, когда прибывает, и это случайная величина, полученная из бета-распределения с параметрами и








( желаемое количество ребер).Таким образом, обобщенная каскадная модель [9] [10] восстанавливается в пределах , а распределения степеней такие же, как в нишевой модели [11] и обобщенная нишевая модель . [10] Этот алгоритм, как описано выше, генерирует сети без циклов (за исключением самоциклов, если новый узел сам считается одним из своих кандидатов-соседей ). Чтобы циклы любой длины были возможны, можно рассмотреть новые ребра-кандидаты, в которых новый узел является внутренним соседом, а также теми, в которых он был бы внешним соседом. Вероятность принятия этих ребер, , тогда зависит от .




Обобщенная модель преференциального хищничества [6] аналогичен описанному выше, но имеет определенные преимущества. В частности, он более поддается аналитическому анализу, и можно создавать сети с точным числом ребер. .Сеть начинается с базальные узлы, а затем еще новые узлы добавляются следующим образом. Когда каждый из них входит в систему, ему случайным образом назначается один сосед из числа тех, кто уже там. Тогда каждый узел имеет целочисленный временный трофический уровень. . Остальные ребра вводятся следующим образом. Каждая пара узлов имеет две связанные временные трофические дистанции, и . Каждое из этих ребер-кандидатов принимается с вероятностью, зависящей от этого временного расстояния. Клейз и Джонсон [6] использовать







поскольку они находят распределение трофических расстояний в нескольких видах сетей примерно нормальным , и этот выбор приводит к диапазону параметра в котором . После введения всех ребер необходимо пересчитать трофические уровни всех узлов, поскольку они будут отличаться от первоначально назначенных временных, если только .Как и в модели преимущественного хищничества, средний параметр некогерентности полученных сетей является монотонно возрастающей функцией для . На рисунке выше показаны две сети с разной трофической когерентностью, созданные с помощью этого алгоритма.



Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Джонсон С., Домингес-Гарсиа В., Донетти Л., Муньос М.А. (2014). «Трофическая согласованность определяет стабильность пищевой сети» . Proc Natl Acad Sci США . 111 (50): 17923–17928. arXiv : 1404.7728 . Бибкод : 2014PNAS..11117923J . дои : 10.1073/pnas.1409077111 . ПМЦ 4273378 . ПМИД 25468963 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Левин С. (1980). «Некоторые показатели трофической структуры, применимые к сложным пищевым сетям». J Теория Биол . 83 (2): 195–207. дои : 10.1016/0022-5193(80)90288-X .
- ^ Jump up to: а б с д и ж Джонсон С. и Джонс Н.С. (2017). «Без петель в сетях связано с трофической когерентностью» . Proc Natl Acad Sci США . 114 (22): 5618–5623. arXiv : 1505.07332 . дои : 10.1073/pnas.1613786114 . ПМЦ 5465891 . ПМИД 28512222 .
- ^ Клэйз Дж. и Джонсон С. (2017). «Происхождение семейств мотивов в пищевых сетях» . Научные отчеты . 7 (1): 16197. arXiv : 1609.04318 . Бибкод : 2017НатСР...716197К . дои : 10.1038/s41598-017-15496-1 . ПМК 5700930 . ПМИД 29170384 .
- ^ Домингес-Гарсиа В., Джонсон С., Муньос М.А. (2016). «Интервальность и когерентность в сложных сетях». Хаос . 26 (6): 065308. arXiv : 1603.03767 . Бибкод : 2016Хаос..26f5308D . дои : 10.1063/1.4953163 . ПМИД 27368797 . S2CID 16081869 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с д Клэйз Дж. и Джонсон С. (2016). «От нейронов к эпидемиям: как трофическая когерентность влияет на процессы распространения». Хаос . 26 (6): 065310. arXiv : 1603.00670 . Бибкод : 2016Хаос..26f5310K . дои : 10.1063/1.4953160 . ПМИД 27368799 . S2CID 205214650 .
- ^ Ньюман, МЭД (2003). «Структура и функции сложных сетей». Обзор СИАМ . 45 (2): 167–256. arXiv : cond-mat/0303516 . Бибкод : 2003SIAMR..45..167N . дои : 10.1137/S003614450342480 . S2CID 221278130 .
- ^ Россберг А.Г., Брэннстрем А., Дикманн У. (2010). «Структура пищевой сети в пространствах трофических ниш низкой и высокой размерности» . Интерфейс JR Soc . 7 (53): 1735–1743. дои : 10.1098/rsif.2010.0111 . ПМЦ 2988264 . ПМИД 20462875 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Коэн Дж. Э. и Ньюман К. М. (1985). «Стохастическая теория общественных пищевых сетей I. Модели и агрегированные данные». Учеб. Р. Сок. Б. 224 (1237): 421–448. Бибкод : 1985РСПСБ.224..421С . дои : 10.1098/rspb.1985.0042 . S2CID 52993453 .
- ^ Jump up to: а б Стоуффер Д.Б., Камачо Дж., Амарал ЛАН (2006). «Надежный показатель интервальности пищевой сети» . Proc Natl Acad Sci США . 103 (50): 19015–19020. Бибкод : 2006PNAS..10319015S . дои : 10.1073/pnas.0603844103 . ПМЦ 1748169 . ПМИД 17146055 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Уильямс Р.Дж. и Мартинес Н.Д. (2000). «Простые правила порождают сложные пищевые сети». Природа . 404 (6774): 180–183. Бибкод : 2000Natur.404..180W . дои : 10.1038/35004572 . ПМИД 10724169 . S2CID 205004984 .