Диапазон фракционирования
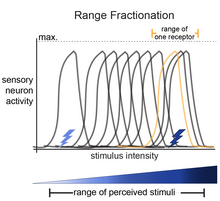
Фракционирование диапазона — это термин, используемый в биологии для описания способа, с помощью которого группа сенсорных нейронов способна кодировать стимул различной величины. Органы чувств обычно состоят из множества сенсорных рецепторов, измеряющих одно и то же свойство. Эти сенсорные рецепторы демонстрируют ограниченную степень точности из-за верхнего предела скорости стрельбы . Если рецепторы наделить четко выраженными передаточными функциями таким образом, что точки наибольшей чувствительности разбросаны по оси измеряемого качества, можно повысить точность работы органа чувств в целом.
В основе идеи фракционирования диапазона лежит то, что каждый стимул (например, прикосновение) имеет диапазон интенсивности, который можно ощутить (от легкого прикосновения до глубокого/сильного прикосновения). Чтобы организм мог ощущать диапазон интенсивности стимула, сенсорные нейроны настроены на части всего диапазона. В совокупности характер активности сенсорных нейронов определяет, как организм может идентифицировать конкретные параметры стимула. Это было показано для проприоцептивных нейронов ноги саранчи . [1] [2] проприоцептивные нейроны палочника, [3] Нейроны органа Джонстона у дрозофилы. [4] и в слухочувствительных нейронах сверчков. [5] [6] [7]
Фракция диапазона аналогична теории меченых линий в том, что они обе описывают феномен, посредством которого сенсорные нейроны разделяют задачу кодирования диапазона интенсивности стимула. Однако разница заключается в нижестоящих синаптических партнерах. Теория меченых линий описывает постсинаптически полностью разделенные каналы . Напротив, сенсорные нейроны, которые используют фракционирование диапазона, имеют общих синаптических партнеров, и именно их коллективная активность информативна для типа стимула.
Ссылки
[ редактировать ]- ^ Ашервуд П.Н., Рунион Х.И., Кэмпбелл Дж.И. (1968). «Строение и физиология хордотонального органа ноги саранчи». Журнал экспериментальной биологии . 48 (2): 305–323. дои : 10.1242/jeb.48.2.305 .
- ^ Мэтисон Т. (1992). «Диапазонное фракционирование в заднегрудном хордотональном органе бедренной кости саранчи». Журнал сравнительной физиологии А. 170 (4): 509–520. дои : 10.1007/BF00191466 . S2CID 26197182 .
- ^ Хофманн Т., Кох Ю.Т., Басслер У. (1 января 1985 г.). «Физиология бедренного хордотонального органа палочника Cuniculina Impigra » . Журнал экспериментальной биологии . 114 (1): 207–223. дои : 10.1242/jeb.114.1.207 . ISSN 1477-9145 .
- ^ Пателла П., Уилсон Р.И. (апрель 2018 г.). «Функциональные карты механосенсорных особенностей мозга дрозофилы» . Современная биология . 28 (8): 1189–1203.e5. дои : 10.1016/j.cub.2018.02.074 . ПМЦ 5952606 . ПМИД 29657118 .
- ^ Олдфилд Б.П. (1983). «Центральные проекции первичных слуховых волокон у Tettigoniidae (Orthoptera: Ensifera)». Журнал сравнительной физиологии А. 151 (3): 389–395. дои : 10.1007/bf00623914 . ISSN 0340-7594 . S2CID 11430039 .
- ^ Симодзава Т., Кану М. (1984). «Разновидности нитевидных волосков: фракционирование диапазона сенсорными афферентами и зернистыми интернейронами сверчка». Журнал сравнительной физиологии А. 155 (4): 485–493. дои : 10.1007/bf00611913 . ISSN 0340-7594 . S2CID 42806812 .
- ^ Олдфилд Б.П., Кляйндиенст Х.У., Хубер Ф. (октябрь 1986 г.). «Физиология и тонотопическая организация слуховых рецепторов сверчка Gryllus bimaculatus DeGeer». Журнал сравнительной физиологии А. 159 (4): 457–464. дои : 10.1007/bf00604165 . ПМИД 3783498 . S2CID 27321719 .
Эта по биологии статья незавершена . Вы можете помочь Википедии, расширив ее . |