Система секреции типа VI
Система секреции типа VI ( T6SS ) представляет собой молекулярную машину, используемую широким спектром видов грамотрицательных бактерий для транспортировки эффекторов изнутри ( цитоплазмы или цитозоля ) бактериальной клетки через клеточную оболочку в соседнюю клетку-мишень. Хотя часто сообщается, что T6SS был обнаружен в 2006 году исследователями, изучающими возбудителя холеры , Vibrio cholerae , первое исследование, демонстрирующее, что гены T6SS кодируют аппарат экспорта белка, было фактически опубликовано в 2004 году в исследовании секреции белка рыбным патогеном. Эдвардсиелла тарда . [ 1 ] [ 2 ] [ 3 ]
С тех пор было подсчитано, что по крайней мере четверть всех патогенных и непатогенных протеобактериальных геномов кодируют T6SS, включая патогены животных, растений и человека, а также почвенные, экологические или морские бактерии. [ 4 ] [ 5 ] [ 6 ] [ 7 ] Гены, кодирующие T6SS, обычно обнаруживаются в хромосомах, но также могут содержаться в мобильных генетических элементах и плазмидах, опосредующих их перенос и увеличение генетического разнообразия. [ 8 ] [ 9 ] Хотя большинство ранних исследований секреции типа VI были сосредоточены на ее роли в патогенезе высших организмов, теперь известно, что она действует преимущественно в рамках межбактериального антагонизма. [ 3 ]
Структура и механизм
[ редактировать ]
Считается, что T6SS напоминает перевернутый фаг, простирающийся наружу от поверхности бактериальной клетки. Он состоит из 14 белков, которые собираются в три подкомплекса: фаговый хвостоподобный канальец, структура, подобная базовой пластинке фага, и комплекс, охватывающий мембрану, охватывающий клеточную оболочку. Эти три подкомплекса работают вместе, транспортируя белки через оболочку бактериальной клетки в клетку-мишень посредством сократительного механизма. [ 10 ]

Фаг, похожий на хвост
[ редактировать ]Компонент T6SS, похожий на фаговый хвост, представляет собой динамическую трубчатую структуру, которая подвергается циклам сборки и разборки. Его длина может достигать 600 нм, и на электронных микрофотографиях было показано, что он простирается по всей бактериальной цитоплазме. [ 11 ] Канальцы состоят из повторяющихся единиц белков TssA и TssB (VipA/VipB), расположенных в виде оболочки вокруг трубки, построенной из сложенных друг на друга гексамерных колец белка, корегулируемого гемолизином (Hcp). [ 12 ] [ 13 ] На кончике трубки Hcp находится тример белка VgrG, похожего на шип фага, который, в свою очередь, увенчан заостренным белком, содержащим домен PAAR. [ 14 ] Считается, что сокращение оболочки выталкивает трубку Hcp, VgrG и связанные с ним субстраты за пределы бактериальной клетки, где спайк VgrG/PAAR облегчает проникновение через мембрану соседней клетки. Структура канальцев разрушается под действием белка ClpV, расщепляющего АТФ, который находится в основании канальцев. [ 13 ]
Опорная плита
[ редактировать ]Похожий на фаговый хвост каналец T6SS собирается по структуре, аналогичной базовым пластинкам бактериофага. Он состоит из белков TssE, TssF, TssG и TssK. Базовая пластинка и комплекс, похожий на фаговый хвост, взаимодействуют в бактериальной цитоплазме, а затем рекрутируются в клеточную оболочку с помощью мембранного комплекса. [ 10 ]
Мембрана
[ редактировать ]Мембранный комплекс T6SS отвечает за закрепление аппарата на клеточной мембране и обеспечивает канал, по которому субстраты продвигаются за счет сокращения канальцев, похожих на хвост фага. [ 10 ] Этот большой (1,7 мД) комплекс образуется из 10 взаимодействующих звеньев гетеротримера, содержащего TssJ, TssM и TssL. Считается, что он простирается от внутренней мембраны к внешней мембране оболочки грамотрицательных бактериальных клеток, образуя канал, который открывается и закрывается с помощью уникального механизма, подобного радужной оболочке. [ 15 ]
Распознавание субстрата
[ редактировать ]В отличие от субстратов других систем секреции (таких как общий секреторный путь или системы секреции III и IV), субстраты T6SS, как известно, не обладают какими-либо универсально идентифицирующими признаками. Вместо этого они распознаются и выбираются для секреции через один из двух структурных компонентов аппарата. Один класс субстратов связывается внутри поры гексамера белка, ко-регулируемого гемолизином (Hcp). [ 16 ] Поскольку субстраты нестабильны в отсутствие этого взаимодействия, считается, что комплексы субстрат-Hcp секретируются вместе, а не Hcp служит пассивным канальцем, через который проходят субстраты. Представители второго класса субстратов нацелены на секрецию посредством взаимодействия с шипоподобным белком VgrG, похожим на хвост фага. Эти субстраты часто представляют собой модульные белки, такие как токсины Rhs , которые обладают доменом PAAR для взаимодействия с VgrG на одном конце. [ 5 ] Есть также случаи, когда VgrG и субстрат являются частями одного и того же белка. [ нужна ссылка ]
Антиэукариотический
[ редактировать ]Хотя предковая функция T6SS, по-видимому, направлена на бактерии, было идентифицировано несколько систем, которые эволюционировали для нацеливания на эукариотические клетки. В целом, эти системы, нацеленные на эукариоты, участвуют в возникновении заболеваний. Например, внутриклеточному патогену Francesella tularensis требуется активность T6SS, чтобы выйти из фагосом и реплицироваться в цитоплазме макрофагов. [ 17 ] Механизм, с помощью которого секретируемые белки способствуют вирулентности F. tularensis , до сих пор неизвестен. T6SS холерного вибриона играет двойную роль: он способен воздействовать как на бактериальные, так и на эукариотические клетки. [ 18 ] По крайней мере, один субстрат, который он секретирует, специализируется на нацеливании на эукариотические клетки и действует путем перекрестного связывания белка цитоскелета актина. [ 19 ] Burkholderia pseudomallei и Edwardsiella tarda — два других организма, обладающих T6SS, который, по-видимому, предназначен для нацеливания на эукариоты. T6SS патогена растений Xanthomonas citri защищает его от хищной амебы Dictyostelium discoideum . [ 20 ]
Антибактериальный
[ редактировать ]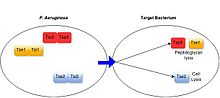
Было показано, что широкий спектр грамотрицательных бактерий обладает антибактериальным ССТ6, включая условно-патогенные микроорганизмы, такие как Pseudomonas aeruginosa , [ 21 ] облигатные виды-комменсалы, обитающие в кишечнике человека ( Bacteroides spp.), [ 22 ] и ассоциированные с растениями бактерии, такие как Agrobacterium tumefaciens . [ 23 ] Эти системы оказывают антибактериальную активность за счет функции секретируемых ими субстратов. [ 3 ] Все охарактеризованные белки T6SS, нацеленные на бактерии, действуют как токсины, либо убивая, либо предотвращая рост клеток-мишеней. Механизмы токсичности по отношению к клеткам-мишеням, проявляемые субстратами T6SS, разнообразны, но обычно включают воздействие на высококонсервативные бактериальные структуры, включая деградацию клеточной стенки за счет активности амидазы или гликогидролазы, разрушение клеточных мембран за счет активности липазы или образования пор, расщепление ДНК. и деградация основного метаболита НАД+. [ 3 ] [ 24 ] T6SS-положительные виды бактерий предотвращают опосредованную T6SS интоксикацию собственных и родственных клеток, вырабатывая иммунные белки, специфичные для каждого секретируемого токсина. Белки иммунитета функционируют путем связывания с белками-токсинами, часто в их активном центре, тем самым блокируя их активность. [ 25 ] [ 3 ]
Регулирование
[ редактировать ]Система GacS/RSM
[ редактировать ]Некоторые исследования были посвящены регуляции T6SS с помощью двухкомпонентных систем . У P. aeruginosa было обнаружено, что двухкомпонентная система GacS/Rsm участвует в регуляции системы секреции типа VI. Эта система регулирует экспрессию малых регуляторных молекул РНК Rsm , а также участвует в формировании биопленок . При стимуляции пути GacS/Rsm увеличение количества молекул Rsm приводит к ингибированию мРНК-связывающего белка RsmA. RsmA представляет собой ингибитор трансляции, который связывается с последовательностями вблизи сайта связывания рибосомы для экспрессии гена T6SS. Этот уровень регуляции также наблюдался у P. fluorescens и P. syringae . [ 26 ] [ 27 ]
Определение кворума
[ редактировать ]Существуют различные примеры, в которых определение кворума регулирует T6SS. В исследованиях T6SS Vibrio cholerae было обнаружено, что серотип O37 имеет высокую экспрессию гена vas . С другой стороны, серотипы O139 и O1 демонстрируют обратное: заметно низкую экспрессию гена vas . Было высказано предположение, что различия в выражении объясняются различиями в уровнях чувства кворума. У Vibrio cholerae сигналы аутоиндуктора-1 (AI-1) обнаруживаются LuxQ, сенсорной киназой . LuxQ активирует LuxU, который затем действует на LuxO, ДНК-связывающий белок, который подавляет экспрессию гена HapR . Считается, что делеции HapR LuxO приводят к сильной индукции экспрессии гена vas и, следовательно, экспрессии T6SS, демонстрируя, что T6SS регулируется в той или иной форме посредством определения кворума. [ 28 ] Однако штаммы O1 с делециями LuxO по-прежнему имели относительно спокойный T6SS по сравнению со штаммом O37, что позволяет предположить, что здесь также задействованы дополнительные факторы. [ 29 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Рао П.С., Ямада Ю., Тан Ю.П., Люнг К.Ю. (2004). «Использование протеомики для выявления новых детерминант вирулентности, необходимых для патогенеза Edwardsiella tarda» . Мол Микробиол . 53 (2): 573–86. дои : 10.1111/j.1365-2958.2004.04123.x . ПМИД 15228535 . S2CID 25901071 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Худ Р.Д., Петерсон С.Б., Мугас Дж.Д. (март 2017 г.). «От поиска к золоту: обнаружение того, что секреция типа VI нацелена на бактерии» . Клетка-хозяин и микроб . 21 (3): 286–289. дои : 10.1016/j.chom.2017.02.001 . ПМК 6404758 . ПМИД 28279332 .
- ^ Jump up to: а б с д и Рассел А.Б., Петерсон С.Б., Мугас Дж.Д. (февраль 2014 г.). «Эффекторы системы секреции типа VI: целевые яды» . Обзоры природы. Микробиология . 12 (2): 137–48. дои : 10.1038/nrmicro3185 . ПМК 4256078 . ПМИД 24384601 .
- ^ Бойер, Фредерик; Фишант, Гвеннаэль; Берто, Жереми; Ванденбрук, Ив; Аттри, Ина (декабрь 2009 г.). «Раскрытие системы секреции бактерий типа VI с помощью полногеномного анализа in silico: что можно узнать из доступных микробных геномных ресурсов?» . БМК Геномика . 10 (1): 104. дои : 10.1186/1471-2164-10-104 . ISSN 1471-2164 . ПМК 2660368 . ПМИД 19284603 .
- ^ Jump up to: а б Шнайдер М.М., Бут С.А., Хо Б.Т., Баслер М., Мекаланос Дж.Дж., Лейман П.Г. (август 2013 г.). «Белки с повторами PAAR обостряют и разнообразят спайки системы секреции типа VI» . Природа . 500 (7462): 350–353. Бибкод : 2013Natur.500..350S . дои : 10.1038/nature12453 . ПМЦ 3792578 . ПМИД 23925114 .
- ^ Бингл, Льюис Э.Х.; Бейли, Кристофер М; Паллен, Марк Дж (февраль 2008 г.). «Секреция VI типа: руководство для начинающих» . Современное мнение в микробиологии . 11 (1): 3–8. дои : 10.1016/j.mib.2008.01.006 . ПМИД 18289922 .
- ^ Монхарас Ферия, Хулия; Вальвано, Мигель А. (19 октября 2020 г.). «Обзор антиэукариотических эффекторов T6SS» . Границы клеточной и инфекционной микробиологии . 10 : 584751. дои : 10.3389/fcimb.2020.584751 . ISSN 2235-2988 . ПМЦ 7641602 . ПМИД 33194822 .
- ^ Робинсон, Лука; Лиау, Джени; Омоле, Захра; Ся, Донг; ван Влит, Арну Х.М.; Корчионивоски, Николае; Хачани, Абдеррахман; Гундогду, Озан (29 июня 2021 г.). «Биоинформатический анализ системы секреции Campylobacter jejuni типа VI и прогнозирование эффекторов» . Границы микробиологии . 12 : 1751. doi : 10.3389/fmicb.2021.694824 . ISSN 1664-302X . ПМЦ 8285248 . ПМИД 34276628 .
- ^ Филлу, Ален; Хачани, Абдеррахман; Блевес, София (июнь 2008 г.). «Бактериальная машина секреции типа VI: еще один игрок в транспорте белков через мембраны» . Микробиология . 154 (6): 1570–1583. дои : 10.1099/mic.0.2008/016840-0 . hdl : 10044/1/40505 . ПМИД 18524912 .
- ^ Jump up to: а б с Чианфанелли Ф.Р., Монлезун Л., Култерст С.Дж. (январь 2016 г.). «Цель, зарядка, огонь: система секреции типа VI, бактериальное нанооружие». Тенденции в микробиологии . 24 (1): 51–62. дои : 10.1016/j.tim.2015.10.005 . ПМИД 26549582 .
- ^ Баслер М., Пилхофер М., Хендерсон Г.П., Дженсен Г.Дж., Мекаланос Дж.Дж. (февраль 2012 г.). «Секреция типа VI требует динамической сократительной структуры, напоминающей хвост фага» . Природа . 483 (7388): 182–6. Бибкод : 2012Natur.483..182B . дои : 10.1038/nature10846 . ПМЦ 3527127 . ПМИД 22367545 .
- ^ Брюне Ю.Р., Энен Ж., Селия Х., Каскальес Э. (март 2014 г.). «Секреция типа VI и хвостовые трубки бактериофагов имеют общий путь сборки» . Отчеты ЭМБО . 15 (3): 315–21. дои : 10.1002/эмбр.201337936 . ПМЦ 3989698 . ПМИД 24488256 .
- ^ Jump up to: а б Бонеманн Г., Петросюк А., Диманд А., Зентграф Х., Могк А. (февраль 2009 г.). «Ремоделирование канальцев VipA/VipB с помощью ClpV-опосредованной нити имеет решающее значение для секреции белка типа VI» . Журнал ЭМБО . 28 (4): 315–25. дои : 10.1038/emboj.2008.269 . ПМК 2646146 . ПМИД 19131969 .
- ^ Бонеманн Г., Петросюк А., Диманд А., Зентграф Х., Могк А. (февраль 2009 г.). «Ремоделирование канальцев VipA/VipB с помощью ClpV-опосредованной нити имеет решающее значение для секреции белка типа VI» . Журнал ЭМБО . 28 (4): 315–25. дои : 10.1038/emboj.2008.269 . ПМК 2646146 . ПМИД 19131969 .
- ^ Дюран Э., Нгуен В.С., Зуед А., Логгер Л., Пеау-Арноде Г., Аштген М.С., Спинелли С., Демитер А., Бардьо Б., Дюжанкур А., Руссель А., Камбийо С., Каскалес Е., Фронзес Р. (июль 2015 г.). «Биогенез и структура основного комплекса секреторной мембраны VI типа» . Природа . 523 (7562): 555–60. Бибкод : 2015Natur.523..555D . дои : 10.1038/nature14667 . ПМИД 26200339 . S2CID 205244835 .
- ^ Сильверман Дж.М., Агнелло Д.М., Чжэн Х., Эндрюс Б.Т., Ли М., Каталано С.Э., Гонен Т., Мугус Дж.Д. (сентябрь 2013 г.). «Корегулируемый гемолизином белок является экспортируемым рецептором и шапероном субстратов секреции типа VI» . Молекулярная клетка . 51 (5): 584–93. doi : 10.1016/j.molcel.2013.07.025 . ПМЦ 3844553 . ПМИД 23954347 .
- ^ Nano FE, Шмерк С (июнь 2007 г.). «Остров патогенности Франциселлы». Анналы Нью-Йоркской академии наук . 1105 (1): 122–37. Бибкод : 2007NYASA1105..122N . дои : 10.1196/анналы.1409.000 . ПМИД 17395722 . S2CID 35958075 .
- ^ Донг Т.Г., Хо Б.Т., Йодер-Хаймс Д.Р., Мекаланос Дж.Дж. (февраль 2013 г.). «Идентификация T6SS-зависимых эффекторных и иммунных белков с помощью Tn-seq в холерном вибрионе » . Труды Национальной академии наук Соединенных Штатов Америки . 110 (7): 2623–8. Бибкод : 2013PNAS..110.2623D . дои : 10.1073/pnas.1222783110 . ПМЦ 3574944 . ПМИД 23362380 . ,
- ^ Пукацки С., Ма А.Т., Ревель А.Т., Стертевант Д., Мекаланос Дж.Дж. (сентябрь 2007 г.). «Система секреции типа VI перемещает белок, похожий на шип фага, в клетки-мишени, где он перекрестно связывает актин» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (39): 15508–13. Бибкод : 2007PNAS..10415508P . дои : 10.1073/pnas.0706532104 . ПМК 2000545 . ПМИД 17873062 . ,
- ^ Байер-Сантос Э., Лима Л.Д., Чесети Л.М., Ратагами С.Ю., де Сантана Э.С., да Силва А.М., Фара К.С., Альварес-Мартинес CE (апрель 2018 г.). « Xanthomonas citri T6SS опосредует устойчивость к хищничеству Dictyostelium и регулируется σ-фактором ECF и родственной киназой Ser/Thr». Экологическая микробиология . 20 (4): 1562–1575. Бибкод : 2018EnvMi..20.1562B . дои : 10.1111/1462-2920.14085 . ПМИД 29488354 . S2CID 3579518 .
- ^ Худ Р.Д., Сингх П., Сюй Ф., Гювенер Т., Карл М.А., Тринидад Р.Р., Сильверман Дж.М., Олсон Б.Б., Хикс К.Г., Племел Р.Л., Ли М., Шварц С., Ван Вай, Мерц А.Дж., Гудлетт Д.Р., Мугас Дж.Д. (январь 2010 г.) ). «Система секреции типа VI Pseudomonas aeruginosa направляет токсин на бактерии» . Клетка-хозяин и микроб . 7 (1): 25–37. дои : 10.1016/j.chom.2009.12.007 . ПМЦ 2831478 . ПМИД 20114026 .
- ^ Рассел А.Б., Векслер А.Г., Хардинг Б.Н., Уитни Дж.С., Бон А.Дж., Гу Я.А., Тран БК, Барри Н.А., Чжэн Х., Петерсон С.Б., Чоу С., Гонен Т., Гудлетт Д.Р., Гудман А.Л., Мугас Дж.Д. (август 2014 г.). «Путь, связанный с секрецией типа VI, у Bacteroidetes опосредует межбактериальный антагонизм» . Клетка-хозяин и микроб . 16 (2): 227–236. дои : 10.1016/j.chom.2014.07.007 . ПМЦ 4136423 . ПМИД 25070807 .
- ^ Ма Л.С., Хачани А., Лин Дж.С., Филлу А., Лай Э.М. (июль 2014 г.). « Agrobacterium tumefaciens использует суперсемейство эффекторов ДНКазы секреции типа VI в качестве оружия для межбактериальной конкуренции на растениях» . Клетка-хозяин и микроб . 16 (1): 94–104. дои : 10.1016/j.chom.2014.06.002 . ПМК 4096383 . ПМИД 24981331 .
- ^ Уитни Дж.С., Квентин Д., Савай С., Леру М., Хардинг Б.Н., Ледвина Х.Э., Тран БК, Робинсон Х., Гу Ю.А., Гудлетт Д.Р., Раунсер С., Мугас Дж.Д. (октябрь 2015 г.). «Межбактериальный токсин NAD(P)(+)гликогидролазы требует фактора элонгации Tu для доставки в клетки-мишени» . Клетка . 163 (3): 607–19. дои : 10.1016/j.cell.2015.09.027 . ПМЦ 4624332 . ПМИД 26456113 .
- ^ Хо Б.Т., Донг Т.Г., Мекаланос Дж.Дж. (январь 2014 г.). «Вид на убийство: система секреции бактерий VI типа» . Клетка-хозяин и микроб . 15 (1): 9–21. дои : 10.1016/j.chom.2013.11.008 . ПМЦ 3936019 . ПМИД 24332978 .
- ^ Mougous JD, Cuff ME, Raunser S, Shen A, Zhou M, Gifford CA, Goodman AL, Joachimiak G, Ordoñez CL, Lory S, Walz T, Joachimiak A, Mekalanos JJ (июнь 2006 г.). «Локус вирулентности Pseudomonas aeruginosa кодирует аппарат секреции белка» . Наука . 312 (5779): 1526–30. Бибкод : 2006Sci...312.1526M . дои : 10.1126/science.1128393 . ПМК 2800167 . ПМИД 16763151 .
- ^ Records AR, Gross DC (июль 2010 г.). «Сенсорные киназы RetS и LadS регулируют секрецию и факторы вирулентности Pseudomonas syringae типа VI» . Журнал бактериологии . 192 (14): 3584–96. дои : 10.1128/JB.00114-10 . ПМК 2897360 . ПМИД 20472799 .
- ^ Records AR, Gross DC (июль 2010 г.). «Сенсорные киназы RetS и LadS регулируют секрецию и факторы вирулентности Pseudomonas syringae типа VI» . Журнал бактериологии . 192 (14): 3584–96. дои : 10.1128/JB.00114-10 . ПМК 2897360 . ПМИД 20472799 .
- ^ Сильверман Дж. М., Брюне Ю. Р., Каскальс Э., Мугус Дж. Д. (2012). «Строение и регуляция системы секреции VI типа» . Ежегодный обзор микробиологии . 66 : 453–72. doi : 10.1146/annurev-micro-121809-151619 . ПМК 3595004 . ПМИД 22746332 .