Tarsophlebiidae
| Tarsophlebiidae Временный диапазон: Верхняя юра - нижний меловой
| |
|---|---|

| |
| Tarsophlebia Eximia , Верхняя Юра, Солнхофен Платтенкалк | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Членистоногие |
| Сорт: | Инсекта |
| Заказ: | Одоната |
| Подотряд: | † tarsophlebioptera |
| Семья: | † Tarsophlebiidae Handlirsch , 1906 |
| Тип рода | |
| Тарсофлебия Хаген , 1866
| |
| Роды | |
| |
Tarsophlebiidae - это вымерший семейство среднего размера ископаемых одонатов из юрской и нижней меловой периода Евразии верхней . членом класса Damel-Dragonfly («анизозигоптер») в группе STEM Анизоптера Они являются либо самым базальным , либо родственной группой всех недавних одонатов. Они характеризуются базально открытой дискоидальной клеткой в обеих парах крыльев, очень длинных ног, мужского Cerci в форме весла и гипертрофированным яйцом у женщин. [ 1 ]
Описание
[ редактировать ]Взрослый
[ редактировать ]Тело
[ редактировать ]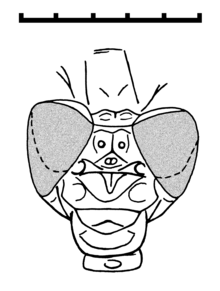

Голова похожа на головку недавних Gomphidae с двумя большими и глобулярными составными глазами , которые отчетливо разделены, но ближе друг к другу, чем у Damselflies ( Zygoptera ). Есть также два головных шва. [ 2 ]
Птероторакса, кажется, даже сильнее искажен, чем в Самих. Ноги чрезвычайно длинные с короткими и сильными шипами, и с очень удлиненной тарзи. Существует три сегмента Tarsal ( трэзомеры ), из которых самый базальный в два раза больше, чем у других. Пара когти для лапка не хватает вентро-апикального крючка современных одонатов.
Крылья и Венеция крыла
[ редактировать ]Самая большая вида Turanophlebia Sinica достигла размаха крыльев в размере около 10,3 сантиметров (4,1 дюйма), в то время как минорный минор Tarsophlebia tarsophlebia достиг лишь 5,5 сантиметра (2,2 дюйма). [ 1 ]

Венеция крыла характеризуется следующими функциями:
крылья гиалиновые, стройные и не преследуемые; Дискоидальная клетка в основном открыта в обеих парах крыльев, так что Arculus неполный; Внешний уход в дискоидальной клетке очень острая; Большая и острая субдейско -клетка в задних крылах; первичные антеннодальные скобки Ax1 и Ax2 сильнее, чем вторичные антеннодальные перекрестки; узд в дистальном положении при 44-47% длины крыла; узкий с терминальным изломом CP и сильной узловой бороздой; Птеростигма удлиняется (охватывающая несколько клеток) и с наклонной кремной веной; Одна Лестина наклонная вена, присутствующая между RP2 и IR2; На всех крыльях есть пары вторичных продольных вогнутых интеркалярных вен перед передней и задней частью выпуклых вен CUA, MA и IR2, и пристально параллельно им (постеро-интеркаляры всегда длиннее, чем связанные с ними антеро-интеркальные); задних крыльев без вены Cuab; Подобный кроссвеину остатки чашки вены изогнуты и, скорее, похож на ветвь АА. [ 1 ] [ 2 ]
Сексуальный диморфизм
[ редактировать ]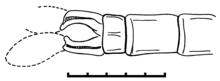
Мужчины отличаются, похожим на весло cerci, [ 1 ] В то время как женщины отличаются очень длинными и худыми, гипертрофированным яйцом, который выступает далеко за пределы живота. [ 2 ]

Самские вторичные гениталии были уникального примитивного типа, с небольшой везикул сперматозоидов на Sternite 3, две пары маленьких пластинчатых гамали на Sternite 2 и очень короткая средняя лигула на стерните. Очевидно, ни одна из этих структур не является гипертрофией как сперма, интраминационный орган (функциональный половой член). В каждом из трех подчиненных недавних одонатов другая часть этого аппарата увеличивается и развивается как индуцитационный орган и устройство для удаления иностранной спермы (конкуренция сперматозоидов): в Zygoptera это лигула, средний процесс Sternite 2; У Epiophlebiidae это боковая пара задних хамули на сегменте 2; и в анизоптере это непарная пузырька сперматозоидов на передней части строита 3. Исключительно хорошо сохранившийся мужской образец Намуротипа Сиппели показал, что в протодонате меганазоптере полностью отсутствовал вторичный генитальный аппарат на брюшных сегментах 2 и 3 и все еще имел первичные гениталии на 9-м сегменте, которые сильно напоминают те из серебряных рыб без крыла, которые не сопоставляются, но депозит наружных. сперматофоры . Только на основе такого спаривающего поведения есть эволюция вторичного совокупного совокупления вообще. Первым шагом, вероятно, было привязанность сперматофора к базальным стернитам мужского живота вместо отложения на субстрат. Женщина теперь должна была получить сперматофор от мужского вентера. Это создало адаптивное давление для создания структур хранения для структур сперматофора и прикрепления для женских гениталий. Tarsophlebiidae, вероятно, представляет это состояние эволюции. Развитие жидких сперматозоидов и интраминализационных органов для совокупления, по -видимому, развивалось три раза параллельно в трех существующих подчиненных. [ 1 ] [ 3 ]
Боковые ушные изделия по бокам базального живота у мужчин -тарсофлебии были описаны Nel et al. (1993). [ 2 ] Тем не менее, Бехли (1996) показал, что эти предполагаемые ученые -мужчины были основаны на неверном толковании постериоров Hamuli, [ 4 ] который был подтвержден Fleck et al. (2004). [ 1 ]
Fleck et al. (2004) продемонстрировали, что мужчина Tarsophlebiidae действительно обладает уникальным типом анальных придаток. Описание калоптериговоподобных придатков Tarsophlebia Eximia , по-видимому, с двумя парами Classers, [ 2 ] [ 4 ] был основан на неверных толкованиях из -за артефактов сохранения. Cerci очень длинные, с двойным стволом базальным черешкам и дистальным расширением. Сломанные двойные черешки двух Cerci обычно неверно истолкованы как две пары Classers, в то время как дистальные пластины упускаются из виду или рассматривались как артефакты. [ 1 ] Действительно, Tarsophlebia не обладает ни зигоптероидами, ни подобными анизоптероидам. Там нет видимых парапрокций (как в Zygoptera) и epiproct (как у Epiophlebiidae и Anisoptera). Если эти структуры вторично уменьшены или отсутствуют, в основном отсутствуют, не ясны.
Личинка
[ редактировать ]Никаких ископаемых личинок этого вымершего семейства еще не обнаружено.
Классификация
[ редактировать ]Семья была создана Хандлиршем (1906) на основе типа рода Tarsophlebia Hagen , 1866. [ 5 ]
Tarsophlebiopsis mayi был признан действительным большинством авторов до самого последнего пересмотра семьи Fleck et al. (2004), [ 6 ] который продемонстрировал, что голотип этого таксона, скорее всего, является лишь аберрантным образцом тарсофлебии . [ 1 ] [ 2 ]
Семейство содержит 2 описанных рода с полностью 9 действительными видами: [ 7 ]
- Гонка † Тарсофлебия Хаген, 1866 [ 8 ] (Тип рода; синоним: Tarsophlebiopsis tillyard, 1923) [ 9 ]
Диагноз: кубито-анальные районы передних крыльев и задних крыльев с четырьмя рядами клеток или меньше; 16 или менее постснодальные кроссвейны; Менее десяти вторичных антенодальных кроссвейнов в Хиндвинге; и IR1 относительно короткий.

- Tarsophlebia minor Fleck et al., 2004 [ 1 ] - (Тип Местности: Эйхштатт, Бавария, Германия; тип горизонта: Верхний юрский период, Malm Zeta 2b, нижний титониан / 150 млн лет, зона Hybonotum, Solnhofener Plattenkalk; Голотип: № 55 в MCZ; размер: задницу 26 мм)
- Род Turanophlebia Pritykina, 1968 [ 11 ]
Диагноз: крылья с более плотным веновлением; наличие более 25 постнадальных перекрестных веков (11-16 в Tarsophlebia ); шесть или более рядов клеток между CuA и задним краем задних крыльев (менее пяти рядов в Tarsophlebia ); Более 10 вторичных антеннодальных перекрестных веков в задних крылах (менее 10 в тарсофлебии ); IR1 дольше, чем в Tarsophlebia ; Наличие длинных вторичных продольных, а не зигзагообразных вен в области между IR2 и RP2.- Turanophlebia anglican fleck et al., 2004 [ 1 ] - (Тип местности: Clockhouse (Butterley) Brickworks, Англия; тип горизонт: нижний меловый, верхний гаутеривиан / 130 млн лет, Нижняя глина Уилда ; голотип: № Bmb 018531 в музее Бут в Брайтоне / Великобритания; размер: задницы 39,7 мм)
- Turanophlebia liaoningensis fang & Zheng, 2022 [ 12 ] - (Тип местности: деревня Хуанбанжигу, Западный Ляонинг, Китай; тип горизонт: Нижний мелочный, формация Йикса )
- Turanophlebia Martynovi Pritykina, 1968 [ 11 ] - (тип видов; тип местности: Каратау-Михайаловка, Чимкент-регион, южный Казахстан; тип горизонт: Верхняя юрская игра, Калловиан-Киммериджс или Оксфордский- Киммериджиан / 163 мю, формация Карабастау ; Голотип: нет. ;
- Turanophlebia Mongolica Fleck et al., 2004 [ 1 ] -(Тип Местности: Бон-Тсагаан, Баянхонгор Аймак, Центральная Монголия; тип горизонт: нижний меловой, хаутерива-бремиан / 129 мю, формация Дзун-Бейна; Голотип: №. Пять 3559/69 в палеонтологическом институте. мм)
- Turanophlebia шее (Martynov, 1927) [ 13 ] - (Тип местности: Каратау-Карабастау, Чимкент регион, южный Казахстан; тип горизонт: Верхний юрский период, Калловиан-Киммеридж или Оксфордский- Киммемеридж / 163 Мия, Формирование Карабастау ; Голотип: №. Пин 2452/3 в палонтологическом институте Moscow; задним ходом 39,0 мм ;
- Туранофлебия? Sibirica Pritykina, 1977 [ 14 ] , Сибири / Трансбайкалия, Россия; тип горизонт: Нижний меховой, неокомиан-беррийский / 145-140 мя -(Тип местности: Баисса, река Витим, Бельят Республика ; институте Москва 1258 в / палеонтологическом 41,0 мм)
- Turanophlebia Sinica Huang & In, 2009 [ 15 ] -(Тип местности: Западный Ляонинг, PR China; тип горизонт: нижний меловые, Барремано-Аптян / 125-121 Mya, Yixian Formation ; голотип: № NIGP 148201 в Институте геологии и палеонтологии Нанкин. мм)
- Turanophlebia vitimensis fleck et al., 2004 [ 1 ] -(Тип местности: правая сторона реки Вим вниз по течению Рамыворской деревни, район Эрой, Бурьят Республика, Сибири / Трансайкалия, Россия; тип горизонт: Нижний меховой, неокоманский / бремианский / 145-120mya или Барреманоаптян / 125 мю, формация Zaza ; Голотип: нет.
Все остальные описанные таксоны являются синонимами типового вида Tarsophlebia Eximia . [ 1 ] [ 2 ]
Филогения
[ редактировать ]Монофилия Tarsophlebiidae решительно подтверждается следующим набором производных символов ( аутапоморфий ):
задних крыльев с гипертрофированными субдейно -клетками, которая развивается как «псевдоидальная клетка»; слияние вен MAB+MP+CUA для значительного расстояния до разделения MP и CUA в Hindwing; Вена АА сильно изогнулась при внедрении чашки; Чрезвычайно острые дистальные углы переднего крыла дискоидальной и субдоискоидальной клетки. Характеристики тела «отчетливо длительные ноги, с очень длинными лапши» и «мужчина cerci с дистальными экспансиями, похожими на весло», известны из одного вида рода Tarsophlebia ( T. Eximia ) и туранофлебии ( T. vitimensis ) соответственно и, следовательно, принадлежали к общему плану основания всех Tarsophlebiidae. Чрезвычайно продолжительный самка яйца может стать еще одной синапоморфией для семьи, но он известен только из T. eximia и из одного образца T. minor в музее Солнхофена. [ 1 ]
Bechly (1996) и Nel et al. (2004: Приложение 2) предложил длинный список предполагаемых синапоморфий , которые демонстрируют более тесную связь Tarsophlebiidae с Crown Group Odonata:
наличие костющего треугольника как широкое и сильное склеротизация базальной коренной края; дистальная дискоидальная вена mAb (= дистальная сторона дискоидальной клетки) и субдейскоидальная вена (происхождение CuA на MP) выровнены и применяются дорсально сильной склеротизацией, так что эта структура, по -видимому, пересекает вену MP и вогнутую склад Эта вена (образование «дисколальной скобки» Sensu Carle 1982); Эта дисковая скобка выровнена с Arculus в плане основного плана (сохраняется только в некоторых таксонах с открытой дискоидальной клеткой и в передних крыстях эпиофлебии); Средняя часть (первая вилка RP и основание IR2) сдвинута в основном, причем RP3/4 обычно возникает базал субнода (отменен в какой -то Coenagrionoidea) и RP2, возникающих вблизи субнода (в плане первого плана); более производный тип узлов, с изгибом в SCP; Наклонная базальная скобка (все еще присутствующая в протанизоптере и протозигоптере) преобразуется в поперечный Ax0 «базальный кронштейн», который выглядит как первичный антенодальный кроссвейн; Наличие двух сильных первичных антенодальных перекрестных вин Ax1 и AX2 (также присутствующих в некоторых протозигоптерах, например, PIN -код 1/276 из верхнего пермца России); Птеростигма отчетливо закреплена наклонным постбнодальным перекрестком под базальным краем птеростигмы; Наличие сериотированной наклонной вены «О» между RP2 и IR2 (второе, отсутствующее в некоторых таксонах); В медианном пространстве (= базальное пространство) выпуклый остаток СМИ («Редиментальный Cua» Sensu Fraser 1957) подавляется, поскольку он сливается с кубитальным стеблем с общим медио-кубит-стволом (предполагаемое присутствие этого оставить в голотипе Tarsophlebiopsis - это тератология или артефакт сохранения), сходящийся к некоторым протанизоптерам, триадофлебиоморфу и протозигоптеру.
Бехли (1996) предположил, что несколько уникальных симплсуоморфных особенностей всех Tarsophlebiidae указывают на то, что эта семья представляет родственную группу всех недавних одонат. [ 4 ] Эти особенности являются базально открытой дискоидальной клеткой в заднем крыле (вместо закрытого), которая подразумевает неполный Arculus, (тем временем оспариваемое) присутствие четырех тарсомеров равной длины (вместо трех) и очень примитивного состояния мужского вторичного Генитальный аппарат (а именно лигула -аромата; Поэтому Бехли рассматривал сходство Tarsophlebiidae и Epiprocta, упомянутых Nel et al. (1993), а именно, менее разделенные и относительно большие глаза, наличие двух головных швов и маленьких колючек (также присутствующих в Meganisoptera [ 3 ] ), как симплсуоморфии.
Однако, основываясь на кладистическом исследовании 14 символов Fleck et al. (2004) снова предположили, что Tarsophlebiidae, скорее, может быть родственной группой кладской Epiprocta, которая включает Epiophlebiidae и Anisoptera. Тем не менее, этот результат обладает очень низкой статистической поддержкой и может также быть артефактом компьютерного алгоритма экологически числа, потому что ни один из 14 символов не представляет однозначную синапоморфию для Tarsophlebiidae и Epiprocta. [ 1 ]
Huang & Nel (2009) представила убедительные доказательства от новой ископаемой Tarsophlebiidae из Китая о том, что число Tarsomeres составляет всего три, как и в современных одонатах, но первая Tarsomere примерно в два раза больше, чем у других. [ 15 ] Бехли (1996) уже обсуждал эту возможность и предположил, что такой длинный базальный тарзомер, по сравнению с коротким базальным тарзомерой в протодонатах и всех недавних одонатах, все еще может быть плезиоморфным состоянием, которое могло вызвать слияние двух базальных тарзомеров. [ 4 ] Huang & Nel (2004) утверждают, что нет никаких доказательств такого слияния в окаменелостях и что удлинение первого сегмента также может быть полученной особенностью (Autapomorphy) Tarsophlebiidae.
Два альтернативных филогенетических положения Tarsophlebiidae
Гипотеза Fleck et al. (2004):
| Одоната |
| ||||||||||||||||||||||||
Гипотеза о Бехли (1996, 2007):
| Одонатоптера |
| ||||||||||||||||||
Биология
[ редактировать ]Ничего не известно об экологии и поведении Tarsophlebiidae, но, вероятно, это было очень похоже на недавние одонаты. С их очень длинным яйцом женщины, вероятно, вставили свои яйца в грязь в или рядом с водой, похожими на современные Cordulegastridae . Примечательно, что две другие группы мезозойских одонатов (а именно Steleopteridae и Aeschnidiidae ) имели одинаково удлиненный яйцеклетчик.
Географическое и геологическое распределение
[ редактировать ]Окаменевает отчетность этой семьи варьируется от Верхней Юры Казахстана и Германии до нижнего мела Англии, Трансбайкалии, Монголии и Китая. Там нет записей о Tarsophlebiidae из какого -либо ископаемого населенного пункта за пределами Евразиатского региона. [ 7 ]
История
[ редактировать ]Первыми окаменелостями Tarsophlebiid были образцы Tarsophlebia eximia из верхней юрской сольнхофен Платтенкалк из Германии, описанного как « гетерофлебия экзимия » и Euphaea longiventris от Hagen (1862). [ 1 ] Большие пересмотра этой семьи были предоставлены Nel et al. (1993) и Fleck et al. (2004). [ 1 ] [ 2 ]
Примечательные образцы
[ редактировать ]Помимо важных образцов типа, есть также несколько исключительно хорошо сохранившихся окаменелостей, которые способствуют нашим знаниям о морфологии Tarsophlebiidae. [ 7 ] К ним относятся следующие образцы: [ 1 ]
- Женский образец нет. SOS 3609 ( Tarsophlebia Eximia ) в Юра-Музее Эйхштатт показывает гипертрофированный яйцеклетчик
- Женский образец без номера сбора ( минор Tarsophlebia ) в музее Bürgermeister Müller в Солнхофене (BMMS) показывает гипертрофированный яйцеклетчик (см. Фото)
- мужской образец №. SOS 1720 ( Tarsophlebia eximia ) в Юра-Музее Эйхштатт является единственным известным образцом, который показывает под алкоголь
- мужской образец №. AS-VI-44B (голотип Tarsophlebia Eximia ) в BSPGM показывает кажущуюся сегментацию лапки на четыре сегмента на правых заджебках
- Образец №. JME 1960/66K в Юра-Музее Эйхштатт показывает очевидную сегментацию лапки на четыре сегмента на левой передней части
- мужской образец №. 6126 ( Tarsophlebia Eximia ) в Coll. Карпентер из MCZ показывает идеально сохранившуюся голову в дорсальном обзоре
- мужской образец №. 6222 ( Tarsophlebia Eximia ) в Coll. Карпентер MCZ показывает хорошо сохранившуюся Cerci
- образцы . Мужчина -
- Мужской образец NIGP 148201 (голотип Turanophlebia sinica ) имеет хорошо сохранившуюся голову и наиболее сохранившиеся ноги всех известных окаменелостей Tarsophlebiidae, которые показывают важную сегментацию лапки. В исходном описании (Huang & Nel, 2009: рис. 10) видимый швар между Clypeus и Labrum ошибочно помечен как «мандибулы». [ 15 ]
- Мужской образец Tarsophlebia Eximia в частной коллекции Spiegelberg (Гейдельберг, Германия) показывает идеально сохранившуюся Cerci (см. Фото)
Ссылки
[ редактировать ]- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий Jarzembowski E, Nel A (2004). (PDF) "(PDF )" (PDF) . Геодорсии . 26 (1): 33–60.
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час В A, Martinez-Delclos X. «Анизигоптер» окаменелости. Мартина Числа за пределами серии 3: 1–311.
- ^ Подпрыгнуть до: а беременный Бехли Г., Браукманн С., Зессин В., Грёнинг Е. (2001). «Новые результаты, касающиеся морфологии самых древних стрекоз (Insecta: Odonatoptera) из Намурии Хаген-Ворхалле (Германия)» (PDF) . J. Zool. Система Эвол. Резерв 39 (4): 209–226. doi : 10.1046/j.1439-0469.2001.00165.x .
- ^ Подпрыгнуть до: а беременный в дюймовый Бехли Г. (1996). «Морфологические исследования на крылатых велосипедах недавних стрекоз и их обычных представителей группы (Insecta; Pterygota; Odonata), с особым рассмотрением филогенетической системы и основным планом *odonata». Петалура . Специальный том 2: 1–402.
- ^ Handlirsch A (1906-1908). Сформировать ископаемые насекомые и филогения недавнего. Руководство для палеонтологов и зоологов (на немецком языке). Лейпциг: Энгельман В.П.
- ^ Fraser FC (1955). ( PDF ) (PDF) . Психика : 62 (3) (3): 134–1 doi : 10.1155/ 1955/7
- ^ Подпрыгнуть до: а беременный в [1] Вход базы данных палеобиологии для Tarsophlebiidae
- ^ Хаген Ха (1866). «Нейроптер литографических сланцев в Баварии. Pars I: Tarsophlebia, изофлебия, стенофлебия, анакс». Palaeontographica . 15 : 57–96.
- ^ Подпрыгнуть до: а беременный Tillyard RJ (1923). «Tarsophlebiopsis mayi n. Gen. Et n. Sp., Стрекоза, найденная в камере тела кораллианского аммонита». Геологический журнал . 60 (4): 146–152. Bibcode : 1923geom ... 60..146t . doi : 10.1017/s001675680008523x . S2CID 128609930 .
- ^ Хаген Ха (1862). «О нейропине из литографического сланца в Баварии». Palaeontographica . 10 : 96–145.
- ^ Подпрыгнуть до: а беременный Pritykina LN (1968). «Стрекози Каратау (Одоната)» [Dragonflies of Karatau (Odonata)]. В Ройдендорф Б.Б. (ред.). Yurskoy Nasekomiye karatau [ юрские насекомые Каратау ] (на русском языке). Москва: Академия наук СССР, раздел общей биологии, издательство "Наука". С. 26–54.
- ^ Клык, Руи; Чжэн, Даран (2022-11-23). «Новая дракоза Tarsophlebiid из нижнего мела Западного Ляонинг, северо -восточный Китай (Insecta: Odonatoptera, Panodonata)» . Мерашные исследования . 143 : 105424. DOI : 10.1016/j.cretres.2022.105424 . ISSN 0195-6671 . S2CID 253875737 .
- ^ Martynov AV (1927). "Jurassic fossil Insects from Turkestan. 7. Some Odonata, Neuroptera, Thysanoptera". Izvestiya Akademii Nauk SSSR . 21 (9–11): 757–768.
- ^ Pritykina LN (1977). "New dragonflies from Lower Cretaceous deposits of Transbaikalia and Mongolia". Trudy Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya . 4 : 81–96.
- ^ Подпрыгнуть до: а беременный в Huang Dy, Nel A (2009). «Первые китайские Tarsophlebiidae из нижней меловой образования Йикса, с морфологическими и филогенетическими последствиями (Odonatoptera: Panodonata)» (PDF) . Мерашные исследования . 30 (2): 429–433. doi : 10.1016/j.cretres.2008.08.005 .
Дальнейшее чтение
[ редактировать ]- Карпентер Ф.М. (1992). Superclass Hexapoda. Том 3 части R, членистоногие 4; Трактат о палеонтологии беспозвоночных . Геологическое общество Америки и Университет Канзаса. ISBN 978-0-8137-3019-6 .
- Фрейзер ФК (1957). «Реклассификация Odonata Odonata, основанную на некоторых новых интерпретациях Venation of the Dragonfly Wing». Справочник Королевского зоологического общества Нового Южного Уэльса . 12 : 1–133.
- Гримальди Д. , Энгель М.С. (2005). Эволюция насекомых . Издательство Кембриджского университета . ISBN 978-0-521-82149-0 .
- Rasnitsyn, AP , Quicke, DLJ (2002). История насекомых . Kluwer Academic Publishers . ISBN 978-1-4020-0026-3 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка )
Внешние ссылки
[ редактировать ]- Окаменелости Солнгофена (фотографии ископаемых Solnhofen Tarsophlebia )
- Проект Tree of Life (Page Odonata)
- Гиперболическое филогенетическое дерево Одонаты (Г. Бехли)
- Филогенетическая систематика Odonata - Tarsophlebiidae (G. Bechly, 2007)