Новая Зеландия Меньшая коротковолочная летучая мышь
| Меньшая коротковолодовая летучая мышь | |
|---|---|

| |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Млекопитающие |
| Заказ: | Chiroptera |
| Семья: | Mystacinidae |
| Род: | Мистацина |
| Разновидность: | М. Туберкулата
|
| Биномиальное название | |
| Mystacina Tuberculata Грей , 1843
| |
| Подвид | |
|
М. Т. Tuberculata | |
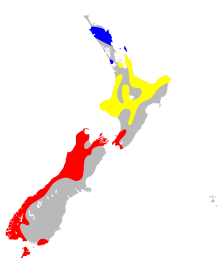
| |
| Ключ:
Диапазон М. Т. Аморика
Диапазон М. Т. Rhyacobia
Диапазон М. Т. Tuberculata
| |

Новая Зеландия Меньшая коротковолодовая летучая мышь ( Mystacina Tuberculata ) представляет собой всеядное всеядное млекопитающее для островов Новой Зеландии. Это один из двух существующих и трех общих наземных видов млекопитающих, уникальных для Новой Зеландии. Его ближайший родственник, новозеландская более широкая коротководья летучая мышь ( M. robusta ), в последний раз видели в 1965 году и предположительно вымерли из-за интенсивного хищничества от судовых крыс, представленных в последние несколько веков. Эти летучие мыши также обычно называют Пекапекой , их название на маори . Меньшие коротковольности летучих мышей имеют уникальные адаптации, которые отличают их от летучих мышей, найденных в других частях мира. Например, они полностью способны двигаться по земле, чтобы найти еду, и мужчины поют, чтобы привлечь партнеров, по очереди. Меньшие коротковольности летучих мышей являются уязвимым видом, поэтому проводится обширные работы по сохранению и исследования для предотвращения вымирания.
Таксономия и эволюция
[ редактировать ]
Таксономическая классификация
[ редактировать ]Mystacina Tuberculata являются членами подготовки Yangochiroptera и недавно были помещены в суперсемейство Noctillionoidea . Они являются членами семейства Mystacinidae , которая содержит единственный род Mystacina . В Мистацине находятся два близкородственных вида, М. Туберкулата и М. Робуста . Поскольку вполне вероятно, что М. Робуста вымер, М. Туберкулата является единственным подтвержденным существующим видом в роде и семействе. Размещение этого вида в таксономиях и филогенетических деревьях ранее было горячо обсуждаемой темой. Ток, стоящий внутри суперсемейства Noctillionoidea, является результатом секвенирования и анализа ДНК. [ 2 ] В противоположность этому некоторые ученые считают, что физические сходства указывают на то, что Mystacinidae попадает под суперсемейство Vespertilionoidea . [ 3 ] Филогенетическое положение этого вида имело множество других предполагаемых положений, в том числе в Phyllostomidae (летучие мыши с листьями) или Emballonuridae (летучих мышей с сак-крылатом и хвостами). [ 4 ] В то время как этот вид считается подвидом, некоторые источники называют вид как один единицу. Департамент сохранения Новой Зеландии признает три подвида. Они делятся по региону, каждый из которых живет; Northern ( Mystacina Tuberculata Aupourica ), Central ( Mystacina Tuberculata Rhyacobi ) и Southern ( Mystacina Tuberculata Tuberculata ). [ 5 ]
Эволюционная история
[ редактировать ]Было высказано предположение, что Mystacinidae возникла на австралийском континенте [ 6 ] и что суперсемейство Noctillionoidea возникло на Гондване . [ 7 ] Когда Гондвана разделилась на более мелкие континенты, члены суперсемейства стали географически изолированными. Эта изоляция привела к миллионам лет уникальной эволюционной истории для каждой семьи. Было подсчитано, что Mystacinidae впервые стала отдельной семьей между 68 и 35 миллионами лет назад. [ 8 ] Двадцать миллионов лет назад из семьи появились два рода; Мистацина и Икаропс . Окаменелости икаропов были обнаружены по всей Австралии, что указывает на то, что теперь, когда-то были хорошо рассеяны виды. [ 8 ] Неизвестно, когда виды в этих родах ( I. Aenae , I. Breviceps и I. Paradox ) вымерли.
Мистацина мигрировала из Австралии в Новую Зеландию, поселившись как первые млекопитающие на этих островах. Основываясь на ископаемых доказательствах, этот род был определенно в Новой Зеландии во время миоцена . [ 9 ] Поэтому мы знаем, что Мистацина стала явным родом около 20 миллионов лет назад и отправилась из Австралии в Новую Зеландию где -то от 5,3 до 20 миллионов лет назад. [ Цитация необходима ] Это предполагает, что Icarops не мигрировал в Новую Зеландию, и что событие колонизации произошло после того, как роды стали различными. Неясно, когда М. Туберкулата и М. Робуста стали отдельными видами.
Распределение и диапазон
[ редактировать ]Меньшие коротковостые летучие мыши были обнаружены в нескольких популяциях, рассеянных по всей стране. На обоих главных островах есть популяции, а также различные окружающие острова. В настоящее время известно несколько ключевых групп населения. есть один На острове Литтл Барьер , один возле вершины Северного острова, семь в центральном районе Северного острова и один на юго-восточной стороне Северного острова. На материке Южного острова есть как минимум два; и один на острове трески дальше на юг. [ 10 ] Летучие мыши могут жить на широком диапазоне высот, от уровня моря до высокого уровня среди покрытых лесом гор. [ 6 ] М. Туберкулата предпочитает жить в нетронутых лесных условиях. Тем не менее, они сообщались в различных местах обитания, таких как сельскохозяйственные угодья и кустарники. [ 11 ]
Анатомия и физиология
[ редактировать ]Меньшие летучие мыши с короткими хвостами выглядят похожими на их родственники Microbat. Они маленькие по размеру и обычно весят от 12 до 15 граммов (0,42 и 0,53 унции) после полного выращивания. [ 12 ] У них относительно большие уши по сравнению с размерами головы и тела, которые сталкиваются вперед, чтобы помочь с эхолокацией. Меньшие коротковольности летучих мышей также имеют относительно большие ноздри, которые сталкиваются наружу, что позволяет этим животным использовать обоняние для руководства своими движениями. Их глаза маленькие, а их зрение относительно слабым, оправдывая их зависимость от звука и обонятельных чувств. [ Цитация необходима ] Эти особенности распространены во всем Chiroptera.
Их крылья относительно меньше, чем у других видов летучих мышей. Из -за этого они снизили ловкость и не могут летать так быстро. [ 13 ] Несмотря на это, они по -прежнему очень способны к полету; Просто меньше, чем другие виды летучих мышей. Каждая летучая мышь покрыта слоем толстого меха, который колеблется от светло -коричневого до темно -коричневого. Подставка, как правило, более светлый цвет, чем остальная часть тела. На заднем конце их тела короткие, но сильные задницы, полезные в их наземной локомоции. Относительно короткие хвосты - это то, что дает этому виду свое общее название. В верхнем центре каждой передней основы большой палец простирается наружу. Это, как и их пальцы, оснащены гвоздями, называемыми базальными когтями. Наличие различных когтей дополнительно позволяет двигаться вдоль земли и полезны при лазании по деревьям. [ 12 ] Базальные когти не видны у других видов хироптеранов.
Торпор
[ редактировать ]Одной из ключевых адаптаций, которые имеют меньшие коротковолодовые летучих мышей, является вход в состояние оцепенения. Торпор допускает продолжение выживания в периоды физиологического стресса, например, когда температура воздуха падает зимой. В этом состоянии летучие мыши неактивны, и только важные функции организма работают для снижения расходов на энергию. Входя в Torpor замедляет скорость метаболизма организма, но он отличается от зимней спячки, поскольку периоды намного короче и повторяются.
Продолжительность тумана обычно составляет от нескольких до нескольких часов за раз, и летучие мыши будут входить в это состояние несколько раз в течение сезона. Torpor используется в течение года, но, в частности, зимой, где они также каждый раз вступают в это состояние. [ 14 ] [ 15 ]
Пищу и диета
[ редактировать ]Предпочтительные продукты
[ редактировать ]
Новая Зеландия Меньшие коротковолодовые летучие мыши-вселенные. Их диета состоит из различных продуктов питания, обычно включающих насекомых, других мелких беспозвоночных, фруктов и деталей цветов. [ 16 ] В целом, они предпочитают потреблять насекомых, когда это возможно. В одном исследовании обнаружили следы мотыльков, пауков и гуков Huhu, присутствующих в их гуано . [ 17 ] Другое исследование показало, что эти летучие мыши способны потреблять половину массы тела в мотынах. [ 12 ] Средняя масса тела от 12 до 15 граммов (от 0,42 до 0,53 унции) приравнивается до от 6 до 7,5 грамма (от 0,21 до 0,26 унции) моли, употребляемых в день. Летучие мыши включают нектар и пыльцу, последний из которых, как полагают, обеспечивает им белок. [ 12 ] Тем не менее, были высказывались предположения и дебаты о том, употребляют ли летучие мыши намеренно пыльцу или пыльца, обнаруженная в их гуано, была фактически проглочена их добычей. [ 17 ] Нектар потребляется, поскольку сахар обеспечивает хороший, несложный источник энергии. Меньшие летучие мыши-важные виды опылителей в Новой Зеландии благодаря их потреблению цветов (либо при проглатывании, либо во втором, через их добычу). Исследователи обнаружили, что эти летучие мыши будут нести большие количества пыльцы от нескольких избранных видов растений, таких как Microspermum Collospermum и Dactylanthus taylorii (древесная роза). [ 18 ] Обеспечение сохранения меньших летучих мышей с короткой хвостами также может обеспечить сохранение видов растений, которые они опыляют.
Методы пищи
[ редактировать ]Одной из определяющих особенностей новозеландских летучих мышей является их способность к добыче как в воздухе, так и на земле. Их способность к добыче на местах необычна для видов летучих мышей. Тем не менее, раннее географическое разделение Новой Зеландии около 80 миллионов лет назад [ 19 ] Зналось, острова развивались свободными от хищников. Без хищников меньшая летучая мышь развивалась, чтобы выжить как в воздушных, так и в наземных местах обитания. Как и другие летучие мыши, в воздухе они ловят пищу, используя эхолокацию для обнаружения местоположения добычи. Меньшие коротковостые летучие мыши на земле, перемещаясь на их конечностях, поставляя добычу, используя обоняние. [ 16 ] У этого вида есть несколько адаптаций, которые помогают в поисках пищи на земле. Например, когда на земле люди будут складываться в свои крылья, чтобы использовать их в качестве передних конечностей - все четыре конечности в четвероногих локомоции. Кроме того, у них есть когти, специализирующиеся на том, чтобы помочь им копаться среди слоев мусора и грязи для добычи. [ 12 ] Во время движения на земле летучие мыши ходят так же, как и другие четвероногие животные. [ 20 ] Новая Зеландия Меньшие коротковолодовые летучие мыши имеют повышенное обоняние по сравнению с другими летучими мышами, как видно из их относительно больших и круглых ноздрей. [ 6 ] Хотя, как и другие летучие мыши, они не являются слепыми, они не сильно зависят от визуальной охоты, и такие сигналы, как аромат и звук, гораздо более критичны. Формация на земле может использоваться для выкопания насекомых и пауков, собирать цветы или упавшие фрукты, а также для сбора других стационарных форм пищи. Формация в воздухе часто ассоциируется с охотой на летающих насекомых, такими как мотыльки. Во время сильного дождя кормление редко, а во время более легкого дождя кормовая активность немного увеличивается. [ 21 ]

Эхолокация
[ редактировать ]Как и другие микробаты, эти летучие мыши используют эхолокацию во время полета. [ 22 ] Меньшие короткохвостые летучие мыши испускают звуки в диапазоне 20-40 кГц . Каждый вызов длится от семи до девяти миллисекунд. [ 23 ] Эхолокация не работает так же эффективно во время питания на земле, потому что эхо может быть нарушено лесным полом и всем, что занимает это пространство. Еще одна причина, по которой эхолокация не работает при поиске пищи на местах, заключается в том, что отрывистые отрывистые спины могут прервать новые эхо, отправляемые, вызвав путаницу для животного и предотвращая правильную работу системы. Эта проблема не возникает, когда летучие мыши находятся в воздухе, так как звук имеет больше расстояния для перемещения, а меньше объектов может нарушить сигнал или вызвать путаницу. [ Цитация необходима ]
Жизненный цикл и социальные системы
[ редактировать ]Социальные системы и спаривание
[ редактировать ]Меньшие коротковолодовые летучие мыши формируют общества деления . Это означает, что популяции будут собираться в определенное время, отдельное и рекордное позже. Где эти животные группируются вместе, известны как педали. Для разграбления они предпочитают находить деревья с большими внутренними полостями, где они создают свой «дом». Сайт обезживания будет содержать несколько деревьев в одном и том же окружении. Количество людей, присутствующих на месте, сильно различается, при этом некоторые колонии состоят из сотен или тысяч отдельных летучих мышей. [ 24 ]
Мужские летучие мыши пытаются привлечь партнера в течение весенних и летних месяцев до начала спаривания, что обычно происходит осенью (март -май в Новой Зеландии). Как и многие другие виды, мужчины привлекают товарищей ночью, используя голосовые звонки. Мужчины будут петь от одного из деревьев, разбитых в колонии в течение нескольких минут. Несколько мужчин будут агрегировать в этом же числе, по очереди петь. [ 24 ] Такое поведение классифицируется как система размножения лека . В этом типе спаривания происходит половой отбор, поскольку мужчины сигнализируют потенциальным женщинам в надежде, что они будут впечатлены и спариваются с мужчинами. Разведение Lek - это то, как мужские летучие мыши «доказывают свою ценность». Следующие события спаривания являются немогамными, и, как наблюдалось мужчины, спариваются с несколькими разными женщинами. [ 25 ] В то время как системы размножения лека наблюдаются по всему животному царству, они редки в летучих мышах и наблюдаются только у одного вида.
Ранняя жизнь и жизненный цикл
[ редактировать ]
Меньшие короткохвостые щенки летучих мышей становятся активными в течение дня после рождения. В течение первых двух дней они безволосые, с слаборазвитыми зубами, крыльями и ушами. [ 12 ] [ 26 ] Щенки способны летать в течение примерно четырех недель. Через шесть недель после роста они покидают беременных, в которых они выросли. Через несколько месяцев они достигли размера своего взрослого тела. [ 6 ] Щенки родились летом (декабрь -февраль в Новой Зеландии). В течение этого летнего периода родов рассеянные люди будут перегруппироваться, образуя большую колонию. Щенки сначала будут кормить молоко, произведенное их матерями. Каждая мать родила одного щенка за сезон. Результатом этого являются более высокие материнские инвестиции в соответствии с потомством, однако система немоногамного спаривания приводит к низким отцовским инвестициям. Щенки поднимаются в специальных разделах сайта Riooning в колонии, посвященного материнской деятельности. Другие летучие мыши будут сгруппировать в этих материнских номерах и помогут в воспитании щенков. [ Цитация необходима ]
Нет записей о средней продолжительности жизни меньших летучих мышей. Было отмечено, что другие летучие мыши в подрядке Янгочироптера живут в течение тридцати лет, что неожиданно для их маленьких размеров тела. [ 27 ] Весной/летом летучие мыши мужчин привлекают партнера. Эти пары будут спариваться осенью. Осенью и зимой летучие мыши распределятся в более мелкие колонические группы, в то время как летучие мыши беременны. Затем, когда он снова становится летом, летучие мыши перегруппируются в большую колонию, а щенки рождаются. Примерно в осени щенки достигли физической зрелости. Неясно, в каком возрасте они достигают сексуальной зрелости. [ Цитация необходима ]
Человеческое воздействие и сохранение
[ редактировать ]Угрозы
[ редактировать ]Новой Зе В настоящее время МСКН . в [ 28 ] Внедрение хищников млекопитающих оказало огромное влияние на количество видов. После колонизации Новой Зеландии и введения трех ключевых видов крыс эндемичные популяции летучих мышей испытывали уменьшение количества. Полинезийские крысы ( R. esculans ) были одним из главных хищников летучих мышей до 19 -го века, когда введение черной крысы ( R. rattus ) и коричневой крысы ( R. norvegicus ) приводило к более тяжелой хищнике. [ 29 ] хищники были колонизировали хищные птицы в Новой Зеландии. До того, как хищные хищные птицы, такие как смеящаяся сова , [ 11 ] Другим распространенным хищником этих летучих мышей являются общие домашние кошки, которые поймают летучих мышей, когда они вылетают из своих мест. Разделки летучих мышей очень уязвимы, так как они содержат более потенциальную добычу в одной области. Если бы эти ноги были обнаружены хищником, они могли бы испытать события массовой смертности; В одном инциденте 102 меньших коротковолодоровных смертей летучих мышей были вызваны одним кошкой дома на центральном северном острове. [ 30 ]
Другой причиной их снижения является повышенное использование земли для антропогенной деятельности, таких как лесозаготовка и сельское хозяйство. Эти действия часто могут привести к негативным последствиям для местной экосистемы, таких как фрагментация леса . [ 31 ] Для местных животных в Новой Зеландии фрагментация представляет такие угрозы, как снижение генетического разнообразия. Тем не менее, возможно, что меньшие летучих мышей не будут влиять на фрагментацию леса, так как, как было замечено, демонстрирует поведенческую пластичность посредством способности менять/занимать различные места обитания. [ 25 ]
Текущие планы сохранения
[ редактировать ]Нынешнее состояние сохранения для меньших летучих мышей управляется в Департаменте сохранения Новой Зеландии (DOC). План восстановления с конца 1990-х годов включает в себя многосторонний подход различных действий, которые необходимо предпринять для предотвращения дальнейшей угрозы этого вида. Некоторые из подробных действий включают создание новых групп летучих мышей на различных островах без хищников вокруг Новой Зеландии, осматривая материковую часть для выявления областей, которые требуют дополнительных усилий по сохранению, и потенциально создать пленную популяцию. [ 32 ]
Ссылки
[ редактировать ]- ^ О'Доннелл, С. (2021). " Mystacina tuberculata " . МСОП красный список угрожаемых видов . 2021 : E.T14261A22070543. doi : 10.2305/iucn.uk.2021-2.rlts.t14261a22070543.en . Получено 22 ноября 2021 года .
- ^ Кеннеди, Мартин; Патерсон, Адриан М.; Моралес, Хуан Карлос; Парсонс, Стюарт; Уиннингтон, Эндрю П.; Спенсер, Хэмиш Г. (ноябрь 1999 г.). «Долго и не хватает этого: длина ветвления и проблема размещения новозеландской короткоххватанной летучей мыши Mystacina». Молекулярная филогенетика и эволюция . 13 (2): 405–416. doi : 10.1006/mpev.1999.0660 . ISSN 1055-7903 . PMID 10603267 .
- ^ Джонс -младший, Дж. Нокс; Картер, Дилфорд С. (1976). «Аннотированный контрольный список с ключами к подсемействам и родам. Биология летучих мышей семейства Нового Света Phyllostomidae, часть I.» Lubbock: Музей Texas Tech. Университет 10 : 7–38. doi : 10.5962/bhl.title.142603 . S2CID 83452376 .
- ^ Кирш, Джон Ау; Хатчон, Джеймс М.; Бирнс, Динна Г.П.; Ллойд, Брайан Д. (1998). «Сродство и историческая зоогеография новозеландской коротковолокнистой летучей мыши Mystacina Tuberculata Grey 1843, выведенная из сравнений с гибридизацией ДНК» . Журнал эволюции млекопитающих . 5 (1): 33–64. doi : 10.1023/a: 1020519019830 . S2CID 23809083 .
- ^ «Леткохволостная летучая мышь» . www.doc.govt.nz. Получено 2021-12-25 .
- ^ Jump up to: а беременный в дюймовый Картер, Джеральд Г.; Riskin, Daniel K. (2006-05-24). "Mystacina tuberculata" . Виды млекопитающих (790): 1–8. doi : 10.1644/790.1 . ISSN 0076-3519 . S2CID 198122536 .
- ^ Рука, Сюзанна Дж.; Бек, Робин доктор медицины; Арчер, Майкл; Симмонс, Нэнси Б.; Ганнелл, Грегг Ф.; Скофилд, Р. Пол; Теннисон, Алан Д.Д.; De Pietri, Vanesa L.; Солсбери, Стивен У.; Worthy, Trevor H. (2018-01-10). «Новая, большая вселяящая летучая мышь (Noctilionoidea: Mystacinidae) выявляет потерянное морфологическое и экологическое разнообразие со времен миоцена в Новой Зеландии» . Научные отчеты . 8 (1): 235. BIBCODE : 2018NATSR ... 8..235H . doi : 10.1038/s41598-017-18403-w . ISSN 2045-2322 . PMC 5762892 . PMID 29321543 .
- ^ Jump up to: а беременный Рука, Сюзанна; Арчер, Майкл; Godthelp, Henk (2005-05-01). «Австралийские олиго-миоценовые мистациниды (Microchiroptera): верхняя зубная зуба, новые таксоны и дивергенция видов новозеландских видов» . Geobios . 38 (3): 339–352. Bibcode : 2005geobi..38..339h . doi : 10.1016/j.geobios.2003.11.005 . ISSN 0016-6995 .
- ^ Tennyson, Alan JD (2010). «Происхождение и история наземных позвоночных Новой Зеландии». Новозеландский журнал экологии . 34 (1): 6–27. ISSN 0110-6465 . JSTOR 24060711 .
- ^ Ллойд, Брайан Д. (2003). «Демографическая история новозеландской короткохвостовой летучей мыши Mystacina Tuberculata, выведенной из модифицированных последовательностей контрольной области» . Молекулярная экология . 12 (7): 1895–1911. Bibcode : 2003molec..12.1895L . doi : 10.1046/j.1365-294x.2003.01879.x . ISSN 1365-294X . PMID 12803640 . S2CID 33906414 .
- ^ Jump up to: а беременный Ллойд, Б.Д. (2001-03-01). «Достижения в Новой Зеландии Mammalogy 1990–2000 гг . Журнал Королевского общества Новой Зеландии . 31 (1): 59–81. Bibcode : 2001jrsnz..31 ... 59L . doi : 10.1080/03014223.2001.9517639 . ISSN 0303-6758 . S2CID 129564107 .
- ^ Jump up to: а беременный в дюймовый и фон Даниэль, MJ (1979-04-01). «Новозеландская коротководья летучая мышь, Mystacina Tuberculata; обзор настоящих знаний» . Новая Зеландия Журнал зоологии . 6 (2): 357–370. doi : 10.1080/03014223.1979.10428375 . ISSN 0301-4223 .
- ^ Норберг, Улла М.; Рейнер, JMV; Lighthill, Майкл Джеймс (1987-09-16). «Экологическая морфология и полеты в летучих мышах (Mammalia; Chiroptera): адаптация крыла, эффективность полета, стратегия поиска пищи и эхолокация» . Философские транзакции Королевского общества Лондона. B, биологические науки . 316 (1179): 335–427. Bibcode : 1987rsptb.316..335n . doi : 10.1098/rstb.1987.0030 .
- ^ Czenze, Zenon J.; Бригам, Р. Марк; Хикки, Энтони -младший; Парсонс, Стюарт (январь 2017 г.). «Холодно и одиноко? Выбор и сезон влияют на узоры оцепенения в меньших летучих мышах» . Oecologia . 183 (1): 1–8. Bibcode : 2017oecol.183 .... 1c . doi : 10.1007/s00442-016-3707-1 . HDL : 10294/8901 . ISSN 0029-8549 . PMID 27561779 . S2CID 37006674 .
- ^ Geiser, F.; Бригам, Р.М. (2000-03-01). «Торпор, тепловая биология и энергетика в австралийских длинно ушах летучих мышей (Nyctophilus)». Журнал сравнительной физиологии б . 170 (2): 153–162. doi : 10.1007/s003600050270 . ISSN 1432-136x . PMID 10791575 . S2CID 5775476 .
- ^ Jump up to: а беременный Джонс, Гарет; Уэбб, Питер I.; Седжли, Джейн А.; О'Доннелл, Колин Ф.Дж. (2003-12-01). «Таинственная мистацина: как новозеландская короткоххватанная летучая мышь (мистацина туберкулата) находит добычу насекомых». Журнал экспериментальной биологии . 206 (23): 4209–4216. doi : 10.1242/jeb.00678 . ISSN 0022-0949 . PMID 14581591 . S2CID 36430937 .
- ^ Jump up to: а беременный Даниэль, MJ (1976-12-01). «Кормление у короткохвостого летучей мыши (Mystacina tuberculata) на фруктах и, возможно, нектаре». Новая Зеландия Журнал зоологии . 3 (4): 391–398. doi : 10.1080/03014223.1976.9517927 . ISSN 0301-4223 .
- ^ Каммингс, Г.; Андерсон, с.; Деннис, Т.; Toth, C.; Парсонс, С. (2014). «Конкуренция за опыление меньшей короткоххватываемой летучей мыши и ее влияние на фенологию цветения некоторых новозеландских эндемиков» . Журнал зоологии . 293 (4): 281–288. doi : 10.1111/jzo.12147 . ISSN 1469-7998 .
- ^ Голдберг, Джулия; Тревик, Стивен А; Патерсон, Адриан М. (2008-10-27). «Эволюция наземной фауны Новой Зеландии: обзор молекулярных доказательств» . Философские транзакции Королевского общества B: биологические науки . 363 (1508): 3319–3334. doi : 10.1098/rstb.2008.0114 . PMC 2607375 . PMID 18782728 .
- ^ Riskin, Daniel K.; Парсонс, Стюарт; Шутт, Уильям А. младший; Картер, Джеральд Г.; Hermanson, John W. (2006-05-01). «Земная локомоция новозеландской короткохвостовой летучей мыши Mystacina Tuberculata и обыкновенной вампиров Bat Desmodus rotundus» . Журнал экспериментальной биологии . 209 (9): 1725–1736. doi : 10.1242/jeb.02186 . ISSN 0022-0949 . PMID 16621953 . S2CID 18305396 .
- ^ Чензе, Зенон; Терли, Тертия (февраль 2021 г.). «Посещение цветов Dactylanthus в Новой Зеландии меньших летучих мышей, по-видимому, влияет на ежедневные осадки» . Новозеландский журнал экологии . doi : 10.20417/nzjecol.45.8 . ISSN 0110-6465 . S2CID 231983713 .
- ^ Симмонс, Джеймс А.; Стейн, Роджер А. (1980). «Акустическая визуализация в сонаре летучей мыши: сигналы эхолокации и эволюция эхолокации». Журнал сравнительной физиологии а . 135 (1): 61–84. doi : 10.1007/bf00660182 . ISSN 0340-7594 . S2CID 20515827 .
- ^ Парсонс, Стюарт (январь 1998 г.). «Влияние ситуации записи на эхолокационные звонки новозеландской меньшей короткохвостовой летучей мыши (Mystacina tuberculatagray)». Новая Зеландия Журнал зоологии . 25 (2): 147–156. doi : 10.1080/03014223.1998.9518145 . ISSN 0301-4223 .
- ^ Jump up to: а беременный Кольер, К. (2020). Атрибуты социальной организации и поведения мужчин в меньшей коротковольности летучих мышей, Mystacina Tuberculata (докторская диссертация, Researchspace@ Auckland).
- ^ Jump up to: а беременный Тот, Кори А.; Деннис, Тодд Э.; Pattemore, David E.; Парсонс, Стюарт (2015). «Женщины как мобильные ресурсы: коммунальные педали способствуют принятию размножения лека в умеренной летучей мыши» . Поведенческая экология . 26 (4): 1156–1163. doi : 10.1093/beheco/arv070 . ISSN 1045-2249 .
- ^ Searchfield, D. (январь 2016 г.). «Первое размножение и воспитание рук новозеландского меньшего хвоста мистацины Tuberculata в зоопарке Окленда». Международный зоопарк . 50 (1): 165–173. doi : 10.1111/izy.12121 . ISSN 0074-9664 .
- ^ Дамманн, Филипп (октябрь 2017 г.). «Медленное старение у млекопитающих-бессмысленные из африканских ротов и летучих мышей». Семинары в биологии клеток и развития . 70 : 154–163. doi : 10.1016/j.semcdb.2017.07.006 . ISSN 1084-9521 . PMID 28698112 .
- ^ О'Доннелл, С. (2020-10-15). "Mystacina tuberculata" . Красный список IUCN из угрожаемых видов 2021 года. Doi : 10.2305/iucn.uk.2021-2.rlts.t14261a22070543.en . S2CID 239146418 .
{{cite journal}}: CITE Journal требует|journal=( помощь ) - ^ Король, Кэролин М.; Фостер, Стейси; Миллер, Стивен (2011-06-23). «Инвазивные европейские крысы в Британии и Новой Зеландии: те же виды, разные результаты». Журнал зоологии . 285 (3): 172–179. doi : 10.1111/j.1469-7998.2011.00827.x . ISSN 0952-8369 .
- ^ Scrimgeour, J; Бит, а; Swanney, M (сентябрь 2012 г.). «Хищничество кошек короткохвостых летучих мышей (мистацина Tuberculata Rhyocobia) в лесу Рангатауа, гора Руапеху, Центральный Северный остров, Новая Зеландия» . Новая Зеландия Журнал зоологии . 39 (3): 257–260. doi : 10.1080/03014223.2011.649770 . ISSN 0301-4223 . S2CID 84907680 .
- ^ Adepoju, Kayode Adewale; Салами, Айобами Т. (2017-10-25). «Геопространственная оценка фрагментации леса и ее последствия для экологических процессов в тропических лесах» . Журнал ландшафтной экологии . 10 (2): 19–34. doi : 10.1515/jlecol-2017-0002 . ISSN 1805-4196 . S2CID 89729777 .
- ^ Моллой, Дженис (1995). План восстановления летучей мыши (Пека Пека) (Мистацина, Чалинолобус) . MJ Даниэль, Новая Зеландия. Уничтоженное видовое отделение. Веллингтон, Новая Зеландия: Отделение видов, кафедра сохранения. ISBN 0-478-01570-4 Полем OCLC 154583004 .