CLE пептид
Пептиды CLE ( CLAVATA3/Embryo Surrounding Region-Related ) представляют собой группу пептидов, обнаруженных в растениях, которые участвуют в передаче сигналов в клетках . Производство контролируется генами CLE. При связывании с пептидным рецептором CLE в другой клетке происходит цепная реакция событий, которая может привести к различным физиологическим процессам и процессам развития. Этот сигнальный путь сохраняется у разнообразных наземных растений . [1]
Фон
[ редактировать ]И растения, и животные используют небольшие полипептиды для передачи сигналов при межклеточной коммуникации. Передача сигналов, связанных с CLAVATA3/окружающей областью эмбриона, также известная как растительный пептидный гормон, важна для передачи сигналов от клетки к клетке, а также для связи на большие расстояния. Эти два действия особенно важны для растительных клеток, поскольку они неподвижны и должны осуществлять клеточное расширение. Было обнаружено, что в многоклеточных организмах межклеточная связь имеет решающее значение для многих процессов роста, происходящих внутри организма. Полипептиды из 12 или 13 аминокислот представляют собой зрелые формы белков CLE, которые происходят из консервативных доменов CLE. [2] [3] [4] Выявляется все больше и больше генов CLE, и в этой области проводится все больше исследований. Гены CLE обнаружены не только у семенных растений, но также у ликофитов , мохообразных и зеленых водорослей . [5] [6]
Гены
[ редактировать ]Большинство исследований, посвященных передаче сигналов пептида CLE, проводились с Arabidopsis , поскольку этот геном содержит 32 члена семейства генов CLE. CLV3, принадлежащий к семейству генов CLE, обнаружен в одной или нескольких тканях Arabidopsis . Все 32 члена семейства CLE имеют две общие характеристики, которые включают: кодирование небольшого белка с предполагаемым сигналом секреции на их N-концах и наличие консервативного мотива CLE на их С-концах или рядом с ними. [7] 32 члена семейства генов CLE произошли в результате мутаций исходного гена.
Структуры
[ редактировать ]Пептиды CLE кодируются генами CLE. Эти пептиды различаются по структуре, причем каждая пептидная структура выполняет в растении разную функцию. Установлено, что минимальная длина функционирующих CLE-пептидов составляет 12 аминокислот с несколькими критическими остатками. [8] В растении обнаружены две разные пептидные структуры: А-типа и В-типа. Когда секретируются гормоны А-типа, растение замедляет скорость роста корней, тогда как секреция пептидов В-типа влияет на рост сосудов растения. [9] Секреция пептидов А-типа ускоряет развитие сосудов растения, опосредованное пептидами В-типа. Это говорит о том, что эти два типа пептидов работают вместе, регулируя рост растения. Специфические пептиды: [8]
Пептиды А-типа
- КЛЭ 1/3/4
- КЛЭ 2
- КЛЭ 5/6
- КЛЕ 7
- КЛЕ 8
- КЛЕ 9
- КЛЭ 10
- КЛЕ 11
- КЛЕ 12
- КЛЕ 13
- КЛЕ 14
- КЛЕ 16
- КЛЕ 17
- КЛЕ 18
- КЛЕ 19
- КЛЕ 20
- КЛЕ 21
- КЛЕ 22
- КЛЕ 25
- КЛЕ 26
- КЛЕ 27
- КЛЕ 40
- КЛЕ 45
Пептиды B-типа
- КЛЕ 41/44/ТДИФ
- КЛЕ 42
- КЛЕ 43
- КЛЕ 46
Передача сигналов в апикальной меристеме побега
[ редактировать ]
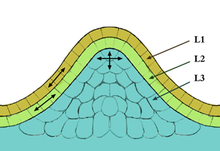
Меристематические клетки дают начало различным органам растения и поддерживают его рост. Существует два типа меристематических тканей: 1) апикальная меристема 2) латеральная меристема. Апикальная меристема бывает двух типов; апикальная меристема побега (SAM) дает начало таким органам, как листья и цветы, а апикальная меристема корня (RAM) обеспечивает меристематические клетки для будущего роста корня. Клетки SAM и RAM быстро делятся и считаются неопределенными, поскольку не обладают каким-либо определенным конечным статусом. В этом смысле меристематические клетки часто сравнивают со стволовыми клетками животных, которые имеют аналогичное поведение и функции. В растениях клетки SAM играют важную роль в общем росте и развитии, это связано с тем, что все клетки, составляющие основные части растения, происходят из апикальной меристемы побега (SAM). В SAM можно обнаружить три различные важные области: центральную зону, периферическую зону и меристему ребра. Каждая из этих областей играет важную роль в производстве новых стволовых клеток в SAM. Все SAM обычно имеют куполообразную форму и имеют слоистые структуры, называемые оболочкой и телом. CLV3 играет важную роль в регуляции производства стволовых клеток в регионе центральной зоны (SAM), это также верно для клеток, стимулирующих ген WUSCHEL (WUS). Комбинация этих двух генов регулирует производство стволовых клеток с помощью WUS, отрицательно или положительно регулируя производство стволовых клеток, контролируя ген CLV3; [10] [11]
Гены других растений
[ редактировать ]Гены CLE были обнаружены у многочисленных однодольных , двудольных и даже мхов . Исследования даже показали, что некоторые растения, такие как рис, содержат домен multi-CLE. [5] [7] Различные CLE-подобные гены также были обнаружены в геномах фитопаразитических нематод, таких как свекловичные, соевые и картофельные нематоды . [12] [5] [13]
Ссылки
[ редактировать ]- ^ Бецуяку С., Сава С., Ямада М. (2011). «Функция пептидов CLE в развитии растений и взаимодействиях растений и микробов» . Книга «Арабидопсис» . 9 : e0149. дои : 10.1199/tab.0149 . ПМК 3268505 . ПМИД 22303273 .
- ^ Кондо Т., Сава С., Киносита А., Мизуно С., Какимото Т., Фукуда Х., Сакагами Ю. (2006). «Растительный пептид, кодируемый CLV3, идентифицированный с помощью анализа MALDI-TOF MS in situ». Наука . 313 (5788): 845–48. Бибкод : 2006Sci...313..845K . дои : 10.1126/science.1128439 . JSTOR 3846935 . ПМИД 16902141 . S2CID 31119711 .
- ^ Ито Ю, Наканомио И, Мотосе Х, Ивамото К, Сава С, Домаэ Н, Фукуда Х (август 2006 г.). «Пептиды Dodeca-CLE как супрессоры дифференцировки стволовых клеток растений». Наука . 313 (5788): 842–5. Бибкод : 2006Sci...313..842I . дои : 10.1126/science.1128436 . ПМИД 16902140 . S2CID 11502739 .
- ^ Охяма К., Шинохара Х., Огава-Охниши М., Мацубаяси Ю. (август 2009 г.). «Гликопептид, регулирующий судьбу стволовых клеток Arabidopsis thaliana». Нат. хим. Биол . 5 (8): 578–80. дои : 10.1038/nchembio.182 . ПМИД 19525968 .
- ^ Перейти обратно: а б с Олкерс К., Гоффард Н., Вейлер Г.Ф., Грессхофф П.М., Матезиус Ю., Фрики Т. (январь 2008 г.). «Биоинформатический анализ семейства сигнальных пептидов CLE» . BMC Завод Биол . 8 :1. дои : 10.1186/1471-2229-8-1 . ПМК 2254619 . ПМИД 18171480 .
- ^ Мива Х, Тамаки Т, Фукуда Х, Сава С (июнь 2009 г.). «Эволюция передачи сигналов CLE: происхождение разнообразия рецепторов CLV1 и SOL2/CRN» . Поведение сигнала объекта . 4 (6): 477–81. дои : 10.4161/psb.4.6.8391 . ПМЦ 2688290 . ПМИД 19816140 .
- ^ Перейти обратно: а б Кок Дж. М., Маккормик С. (июль 2001 г.). «Большое семейство генов, имеющих гомологию с CLAVATA3» . Физиол растений . 126 (3): 939–42. дои : 10.1104/стр.126.3.939 . ПМК 1540125 . ПМИД 11457943 .
- ^ Перейти обратно: а б Ямагучи Ю.Л., Исида Т., Сава С. (2016). «Пептиды CLE и их сигнальные пути в развитии растений» . Дж. Эксп. Бот . 67 (16): 4813–26. дои : 10.1093/jxb/erw208 . ПМИД 27229733 .
- ^ Уитфорд Р., Фернандес А., Де Гроодт Р., Ортега Э., Хилсон П. (ноябрь 2008 г.). «Растительные CLE-пептиды двух различных функциональных классов синергически индуцируют деление сосудистых клеток» . Учеб. Натл. акад. наук. США . 105 (47): 18625–30. Бибкод : 2008PNAS..10518625W . дои : 10.1073/pnas.0809395105 . ПМК 2587568 . ПМИД 19011104 .
- ^ Брэнд Ю, Флетчер Дж. К., Хобе М., Мейеровиц Э. М., Саймон Р. (июль 2000 г.). «Зависимость судьбы стволовых клеток Arabidopsis от петли обратной связи, регулируемой активностью CLV3». Наука . 289 (5479): 617–9. Бибкод : 2000Sci...289..617B . дои : 10.1126/science.289.5479.617 . ПМИД 10915624 .
- ^ Шуф Х., Ленхард М., Хакер А., Майер К.Ф., Юргенс Г., Ло Т. (март 2000 г.). «Популяция стволовых клеток меристем побегов Arabidopsis поддерживается регуляторной петлей между генами CLAVATA и WUSCHEL» . Клетка . 100 (6): 635–44. дои : 10.1016/s0092-8674(00)80700-x . ПМИД 10761929 .
- ^ Ван X, Митчам М.Г., Гао Б., Ли С., Диаб Х., Баум Т.Дж., Хасси Р.С., Дэвис Э.Л. (март 2005 г.). «Ген паразитизма растительно-паразитической нематоды с функцией, аналогичной CLAVATA3/ESR (CLE) Arabidopsis thaliana» . Мол. Растительный патол . 6 (2): 187–91. дои : 10.1111/j.1364-3703.2005.00270.x . ПМИД 20565649 .
- ^ Го Ю, Хан Л., Хаймс М., Денвер Р., Кларк С.Э. (сентябрь 2010 г.). «CLAVATA2 образует отдельный CLE-связывающий рецепторный комплекс, регулирующий спецификацию стволовых клеток Arabidopsis» . Плант Дж . 63 (6): 889–900. дои : 10.1111/j.1365-313X.2010.04295.x . ПМЦ 2974754 . ПМИД 20626648 .
Дальнейшее чтение
[ редактировать ]- Боммерт П., Дже Б.И., Гольдшмидт А., Джексон Д. (октябрь 2013 г.). «Ген Gα кукурузы COMPACT PLANT2 участвует в передаче сигналов CLAVATA, контролируя размер меристемы побега». Природа . 502 (7472): 555–8. Бибкод : 2013Natur.502..555B . дои : 10.1038/nature12583 . ПМИД 24025774 . S2CID 4407655 .
- Чу Х, Лян В, Ли Дж, Хун Ф, Ву Ю, Ван Л, Ван Дж, Ву П, Лю С, Чжан Ц, Сюй Дж, Чжан Д (декабрь 2013 г.). «Сигнальный модуль CLE-WOX регулирует поддержание корневой меристемы и развитие сосудистой ткани риса». Дж. Эксп. Бот . 64 (17): 5359–69. дои : 10.1093/jxb/ert301 . ПМИД 24043854 .
- Кларк С.Е., Уильямс Р.В., Мейеровиц Э.М. (май 1997 г.). «Ген CLAVATA1 кодирует предполагаемую рецепторную киназу, которая контролирует размер побегов и цветочной меристемы арабидопсиса» . Клетка . 89 (4): 575–85. дои : 10.1016/s0092-8674(00)80239-1 . ПМИД 9160749 .
- Джун Дж., Фиуме Э., Редер А.Х., Мэн Л., Шарма В.К., Осмонт К.С., Бейкер С., Ха СМ, Мейеровиц Э.М., Фельдман Л.Дж., Флетчер Дж.К. (декабрь 2010 г.). «Комплексный анализ экспрессии сигнальных генов полипептида CLE и активности сверхэкспрессии у арабидопсиса» . Физиол растений . 154 (4): 1721–36. дои : 10.1104/стр.110.163683 . ПМК 2996011 . ПМИД 20884811 .
- Ло Т., Майер К.Ф., Бергер Дж., Юргенс Г. (январь 1996 г.). «Ген WUSCHEL необходим для целостности побегов и цветочной меристемы арабидопсиса». Разработка . 122 (1): 87–96. дои : 10.1242/dev.122.1.87 . PMID 8565856 .