Агент переноса генов
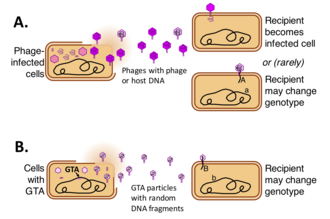
Агенты переноса генов ( GTA ) представляют собой ДНК-содержащие вирусоподобные частицы, которые продуцируются некоторыми бактериями и архей и опосредуют горизонтальный перенос генов . Различные типы GTA произошли независимо от вирусов нескольких бактериальных и архейных линий. Эти клетки производят частицы GTA, содержащие короткие сегменты ДНК, присутствующие в клетке. После того как частицы высвобождаются из клетки-продуцента, они могут прикрепляться к родственным клеткам и инъецировать свою ДНК в цитоплазму. Затем ДНК может стать частью генома клеток-реципиентов. [1] [2] [3] [4]
GTA классифицируются как вириформы в таксономии ICTV . Среди GTA, упомянутых в статье, RcGTA и DsGTA теперь входят в семейство Rhodogtaviriformidae , BaGTA — в Bartogtaviriformidae , а VSH-1 — в Brachygtaviriformidae . [5] Dd1 и VTA пока не имеют классификации.
Открытие агентов переноса генов
[ редактировать ]
Первая система GTA была открыта в 1974 году, когда смешанные культуры штаммов Rhodobacter capsulatus дали высокую частоту клеток с новыми комбинациями генов. [6] Ответственный фактор отличался от известных механизмов переноса генов тем, что не зависел от контакта с клетками, был нечувствителен к дезоксирибонуклеазе и не был связан с продукцией фага. Из-за своей предполагаемой функции его назвали агентом переноса генов (GTA, теперь RcGTA). Совсем недавно были открыты другие системы агентов переноса генов путем инкубации фильтрованной (бесклеточной) культуральной среды с генетически отличным штаммом. [3]
Гены GTA и эволюция
[ редактировать ]
Гены, определяющие GTA, происходят из ДНК бактериофага (фага), которая интегрировалась в хромосому хозяина. Такие профаги часто приобретают мутации, которые делают их дефектными и неспособными производить фаговые частицы. Многие бактериальные геномы содержат один или несколько дефектных профагов, подвергшихся более или менее обширным мутациям и делециям. Агенты переноса генов, как и дефектные профаги, возникают в результате мутации профагов, но они сохраняют функциональные гены головного и хвостового компонентов фаговой частицы (структурные гены) и гены упаковки ДНК. Гены фага, определяющие его регуляцию и репликацию ДНК, обычно удалены, а экспрессия кластера структурных генов находится под контролем клеточных регуляторных систем. Дополнительные гены, которые способствуют производству или поглощению GTA, обычно присутствуют в других местах хромосом. Некоторые из них выполняют регуляторные функции, а другие непосредственно способствуют производству GTA ( например, гены лизиса, полученные из фагов) или поглощению и рекомбинации ( например, гены фагового лизиса). например, производство капсул на клеточной поверхности и транспортных белков ДНК). Эти гены, ассоциированные с GTA, часто находятся под скоординированной регуляцией с основным кластером генов GTA. [7] Белки клеточного лизиса фагового происхождения (холин и эндолизин) затем ослабляют клеточную стенку и мембрану, позволяя клетке лопнуть и высвободить частицы GTA. Количество частиц GTA, производимых каждой клеткой, неизвестно.
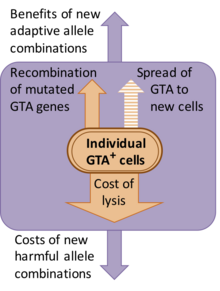
Некоторые системы GTA, по-видимому, являются недавними дополнениями к геномам своих хозяев, но другие сохраняются на протяжении многих миллионов лет. Там, где были проведены исследования дивергенции последовательностей (анализ dN/dS), они указывают на то, что функции белка сохраняются в результате естественного отбора (т.е. дефектные версии устраняются). [8] [9]
Однако природа этого отбора не ясна. Хотя первооткрыватели GTA предполагали, что перенос генов является функцией частиц, предполагаемые выгоды от переноса генов обходятся населению существенными издержками. Большая часть этих затрат возникает из-за того, что клетки, продуцирующие GTA, должны лизироваться (раскрываться), чтобы высвободить частицы GTA, но существуют также генетические затраты, связанные с созданием новых комбинаций генов, поскольку большинство новых комбинаций обычно менее подходят, чем исходная комбинация. [10] Одно из альтернативных объяснений состоит в том, что гены GTA сохраняются, поскольку GTA являются генетическими паразитами, которые инфекционно распространяются на новые клетки. Однако это исключено, поскольку частицы GTA обычно слишком малы, чтобы содержать гены, которые их кодируют. Например, основной кластер RcGTA (см. ниже) имеет длину 14 т.п.н., но частицы RcGTA могут содержать только 4–5 т.п.н. ДНК.
Большинство бактерий не были проверены на наличие GTA, и многие другие системы GTA могут ожидать открытия. Хотя исследования ДНК генов, связанных с GTA, обнаружили гомологи во многих геномах, интерпретация затруднена из-за сложности различения генов, кодирующих GTA, от обычных профаговых генов. [8] [9]
Производство GTA
[ редактировать ]
В лабораторных культурах продукция GTA обычно максимизируется за счет определенных условий роста, которые индуцируют транскрипцию генов GTA; большинство GTA не индуцируются методами лечения, повреждающими ДНК, которые индуцируют многие профаги. Даже в условиях максимальной стимуляции только небольшая часть культуры продуцирует GTA, обычно менее 1%. [11] [12]
Этапы производства GTA основаны на этапах фаговой инфекции. Структурные гены сначала транскрибируются и транслируются, а белки собираются в пустые головки и неприкрепленные хвосты. Затем машина упаковки ДНК упаковывает ДНК в каждую головку, разрезая ДНК, когда головка заполнена, прикрепляя хвост к головке, а затем перемещая вновь созданный конец ДНК на новую пустую головку. В отличие от генов профагов, гены, кодирующие GTA, не вырезаются из генома и не реплицируются для упаковки в частицы GTA. Два наиболее изученных GTA (RcGTA и BaGTA) случайным образом упаковывают всю ДНК в клетке без чрезмерного представительства генов, кодирующих GTA. [11] [13] Количество частиц GTA, производимых каждой клеткой, неизвестно.
GTA-опосредованная трансдукция
[ редактировать ]
Приведет ли высвобождение частиц GTA к переносу ДНК в новые геномы, зависит от нескольких факторов. Во-первых, частицы должны выжить в окружающей среде – об этом мало что известно, хотя сообщается, что частицы весьма нестабильны в лабораторных условиях. [14] Во-вторых, частицы должны встретиться и прикрепиться к подходящим клеткам-реципиентам, обычно представителям того же или близкородственного вида. Как и фаги, GTA прикрепляются к определенным белковым или углеводным структурам на поверхности клетки-реципиента перед инъекцией своей ДНК. В отличие от фага, хорошо изученные GTA, по-видимому, вводят свою ДНК только через первую из двух мембран, окружающих цитоплазму реципиента, и используют другую систему, компетентностную , а не фаговую, для транспортировки одной цепи двойной ДНК. ДНК проходит через внутреннюю мембрану в цитоплазму. [15] [16]
Если механизм рекомбинационной репарации клетки обнаруживает хромосомную последовательность, очень похожую на поступающую ДНК, он заменяет первую на вторую путем гомологичной рекомбинации, опосредованной клеточным белком RecA . Если последовательности не идентичны, это приведет к образованию клетки с новой генетической комбинацией. Однако, если поступающая ДНК не тесно связана с последовательностями ДНК в клетке, она будет деградировать, и клетка будет повторно использовать свои нуклеотиды для репликации ДНК.
Конкретные системы GTA
[ редактировать ]RcGTA/Rhodobactegtaviriform ( Rhodobacter capsulatus )
[ редактировать ]
GTA, продуцируемая альфапротеобактерией Rhodobacter capsulatus , получившая название R. capsulatus GTA (RcGTA), в настоящее время является наиболее изученной GTA. Когда лабораторные культуры R. capsulatus вступают в стационарную фазу, часть бактериальной популяции индуцирует выработку RcGTA, и частицы впоследствии высвобождаются из клеток посредством лизиса клеток . [12] Большинство структурных генов RcGTA кодируются генетическим кластером размером ~ 15 т.п.н. на бактериальной хромосоме. Однако другие гены, необходимые для функции RcGTA, например гены, необходимые для лизиса клеток, расположены отдельно. [2] [17] Частицы RcGTA содержат фрагменты ДНК размером 4,5 т.п.н. с равномерным представлением всей хромосомы, за исключением двукратного провала в месте кластера генов RcGTA.
Регуляция продукции и трансдукции GTA лучше всего изучена у R. capsulatus , где система кворума и CtrA-фосфореле контролирует экспрессию не только основного кластера генов RcGTA, но также системы лизиса клеток холина/эндолизина, шипов головки частиц. , белок прикрепления (возможно, хвостовые волокна), а также гены капсулы и процессинга ДНК, необходимые для функции реципиента RcGTA. Неохарактеризованный стохастический процесс дополнительно ограничивает экспрессию кластера генов только 0,1-3% клеток.
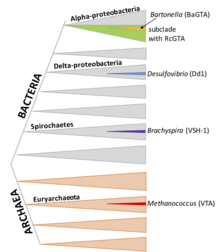
RcGTA-подобные кластеры обнаруживаются в большом субкладе альфапротеобактерий, хотя гены также часто теряются в результате делеции. Недавно было продемонстрировано, что несколько представителей отряда Rhodobacterales производят функциональные частицы, подобные RcGTA. Группы генов, гомологичных RcGTA, присутствуют в хромосомах различных типов альфапротеобактерий. [8]
DsGTA/Диногтавириформ ( Dinoroseobacter shibae )
[ редактировать ]D. shibae , как и R. capsulatus , является членом отряда Rhodobacterales, и его GTA имеет общего предка и многие особенности с RcGTA, включая организацию генов, упаковку коротких фрагментов ДНК (4,2 т.п.н.) и регуляцию посредством определения кворума и Фосфорреле CtrA. [18] Однако его механизм упаковки ДНК обладает гораздо большей специфичностью: резкие пики и спады покрытия позволяют предположить, что он может преимущественно инициировать упаковку в определенных участках генома. ДНК основного кластера генов DsGTA упакована очень плохо.
BaGTA/Bartonegtaviriform ( вид Bartonella )
[ редактировать ]Виды Bartonella являются представителями Alphaproteobacteria, таких как R. capsulatus и D. shibae , но BaGTA не связана с RcGTA и DsGTA. [19] Частицы BaGTA крупнее, чем RcGTA, и содержат фрагменты ДНК размером 14 т.п.н. Хотя эта емкость в принципе могла бы позволить BaGTA упаковать и передать свой кластер GTA размером 14 КБ, измерения покрытия ДНК показывают снижение покрытия кластера. Считается, что соседняя область с высоким покрытием обусловлена локальной репликацией ДНК. [13]
ВШ-1 (Brachyspira hyodysenteriae)
[ редактировать ]Brachyspira — род спирохет; Было показано, что несколько видов несут гомологичные кластеры генов GTA. Частицы содержат фрагменты ДНК длиной 7,5 т.п.н. Производство VSH-1 стимулируется агентом, повреждающим ДНК, митомицином С и некоторыми антибиотиками. Это также связано с обнаруживаемым лизисом клеток, что указывает на то, что значительная часть культуры может продуцировать VSH-1. [20]
Dd1 (десульфовибриондесульфуриканы)
[ редактировать ]D. desulfuricans — почвенная бактерия из семейства дельтапротеобактерий; Dd1 упаковывает фрагменты ДНК размером 13,6 т.п.н. Неясно, какие гены кодируют эту GTA: в бактериальном геноме есть одна область размером 17,8 т.п.н. с фагоподобными структурными генами, но их связь с продукцией GTA еще экспериментально не доказана. [21]
ВТА (Methanococcus voltae)
[ редактировать ]M. voltae — архей; Известно, что его GTA переносит фрагменты ДНК размером 4,4 т.п.н., но иным образом не охарактеризован, [22] хотя было высказано предположение, что дефектный провирус, родственный Methanococcus вирусам типа «голова-хвост» ( Caudoviricetes ) в геноме A3 M. voltae, представляет локус GTA. [23] Возможная терминаза terL ( D7DSG2 ) была снова идентифицирована в 2019 году. [24]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Ланг А.С., Вестбай АБ, Битти Дж.Т. (сентябрь 2017 г.). «Распространение, эволюция и роль агентов переноса генов в прокариотическом генетическом обмене». Ежегодный обзор вирусологии . 4 (1): 87–104. doi : 10.1146/annurev-virology-101416-041624 . ПМИД 28784044 .
- ^ Jump up to: а б Ланг А.С., Жаксыбаева О, Битти Дж.Т. (июнь 2012 г.). «Агенты переноса генов: фагоподобные элементы генетического обмена» . Обзоры природы. Микробиология . 10 (7): 472–82. дои : 10.1038/nrmicro2802 . ПМЦ 3626599 . ПМИД 22683880 .
- ^ Jump up to: а б Стэнтон ТБ (апрель 2007 г.). «Профагоподобные агенты переноса генов - новые механизмы обмена генами для видов Methanococcus, Desulfovibrio, Brachyspira и Rhodobacter». Анаэроб . 13 (2): 43–9. дои : 10.1016/j.anaerobe.2007.03.004 . ПМИД 17513139 .
- ^ Грюль член парламента, Маллиган М.Э., Ланг А.С. (октябрь 2018 г.). «Маленькие внеклеточные частицы с большим потенциалом горизонтального переноса генов: мембранные везикулы и агенты переноса генов» . Письма FEMS по микробиологии . 365 (19). дои : 10.1093/femsle/fny192 . ПМИД 30085064 .
- ^ Когай, Р; Коппенхёфер, С; Битти, Джей Ти; Кун, Дж. Х.; Ланг, А.С.; Жаксыбаева О (2022). «Формальное признание и классификация агентов переноса генов как вириформ» . Эволюция вирусов . 8 (2): veac100. дои : 10.1093/ve/veac100 . ПМЦ 9662315 . ПМИД 36381234 .
- ^ Маррс Б. (март 1974 г.). «Генетическая рекомбинация у Rhodopseudomonas capsulata» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (3): 971–3. Бибкод : 1974ПНАС...71..971М . дои : 10.1073/pnas.71.3.971 . ПМЦ 388139 . ПМИД 4522805 .
- ^ Westbye AB, Beatty JT, Lang AS (август 2017 г.). «Гарантия плененной аудитории: скоординированное регулирование производства агента переноса генов (GTA) и возможностей реципиента с помощью клеточных регуляторов». Современное мнение в микробиологии . 38 : 122–129. дои : 10.1016/j.mib.2017.05.003 . ПМИД 28599143 .
- ^ Jump up to: а б с Шакья М., Суси С.М., Жаксыбаева О. (июль 2017 г.). «Понимание происхождения и эволюции агентов переноса генов α-протеобактерий» . Эволюция вирусов . 3 (2): vex036. дои : 10.1093/ve/vex036 . ПМЦ 5721377 . ПМИД 29250433 .
- ^ Jump up to: а б Тамарит Д., Нойвонен М.М., Энгель П., Гай Л., Андерссон С.Г. (февраль 2018 г.). «Происхождение и эволюция агента переноса гена бартонеллы» . Молекулярная биология и эволюция . 35 (2): 451–464. дои : 10.1093/molbev/msx299 . ПМИД 29161442 .
- ^ Редфилд Р.Дж., Суси СМ (2018). «Эволюция бактериальных агентов переноса генов» . Границы микробиологии . 9 : 2527. дои : 10.3389/fmicb.2018.02527 . ПМК 6209664 . ПМИД 30410473 .
- ^ Jump up to: а б Хайнс А.П., Мерсер Р.Г., Уоттон Д.Э., Бакли С.Б., Ланг А.С. (июль 2012 г.). «Смещение упаковки ДНК и дифференциальная экспрессия генов агента переноса генов внутри популяции во время производства и выпуска агента переноса генов Rhodobacter capsulatus, RcGTA» . Молекулярная микробиология . 85 (2): 314–25. дои : 10.1111/j.1365-2958.2012.08113.x . ПМИД 22640804 .
- ^ Jump up to: а б Фогг ПК, Вестбай АБ, Битти Дж.Т. (2012). Банфилд Б.В. (ред.). «Один за всех или все за одного: гетерогенная экспрессия и лизис клеток-хозяев являются ключом к активности агента переноса генов у Rhodobacter capsulatus» . ПЛОС ОДИН . 7 (8): е43772. Бибкод : 2012PLoSO...743772F . дои : 10.1371/journal.pone.0043772 . ПМЦ 3423380 . ПМИД 22916305 .
- ^ Jump up to: а б Берглунд Э.К., Франк А.К., Калто А., Виннер Петтерссон О., Гранберг Ф., Эрикссон А.С., Нэслунд К., Хольмберг М., Линдроос Х., Андерссон С.Г. (июль 2009 г.). «Репликация генов адаптации хозяина связана с агентами переноса генов в геноме мыши, заражающей Bartonella grahamii» . ПЛОС Генетика . 5 (7): e1000546. дои : 10.1371/journal.pgen.1000546 . ПМЦ 2697382 . ПМИД 19578403 .
- ^ Маррс, Б.; Йен, ХК; Солиоз, М. (1 августа 1975 г.). «Высвобождение и поглощение агента переноса генов Rhodopseudomonas capsulata» . Журнал бактериологии . 123 (2): 651–657. дои : 10.1128/jb.123.2.651-657.1975 . ISSN 1098-5530 . ПМК 235772 . ПМИД 1150627 .
- ^ Бримакомб Калифорния, Стивенс А., Джун Д., Мерсер Р., Лэнг А.С., Битти Дж.Т. (февраль 2013 г.). «Кворум-чувствительная регуляция капсулярного полисахаридного рецептора агента переноса гена Rhodobacter capsulatus (RcGTA)» . Молекулярная микробиология . 87 (4): 802–17. дои : 10.1111/mmi.12132 . ПМК 3641046 . ПМИД 23279213 .
- ^ Бримакомб, Калифорния, Дин Х, Джонсон Дж. А., Битти Дж. Т. (август 2015 г.). «Гомологи генетических трансформационных генов импорта ДНК необходимы для способности реципиента агента переноса генов Rhodobacter capsulatus, регулируемой регулятором ответа CtrA» . Журнал бактериологии . 197 (16): 2653–63. дои : 10.1128/JB.00332-15 . ПМЦ 4507343 . ПМИД 26031909 .
- ^ Вестбай А.Б., Люнг М.М., Флоризон С.М., Тейлор Т.А., Джонсон Дж.А., Фогг ПК, Битти Дж.Т. (ноябрь 2013 г.). «Концентрация фосфата и предполагаемый белок сенсорной киназы CckA модулируют лизис клеток и высвобождение агента переноса гена Rhodobacter capsulatus» . Журнал бактериологии . 195 (22): 5025–40. дои : 10.1128/JB.00669-13 . ПМЦ 3811591 . ПМИД 23995641 .
- ^ Томаш Дж., Ван Х., Холл А.Т., Патцельт Д., Пройсс М., Петерсен Дж., Бринкманн Х., Банк Б., Бхуджу С., Ярек М., Гефферс Р., Ланг А.С., Вагнер-Дёблер И. (январь 2018 г.). «Упаковка ДНК Dinoroseobacter shibae в частицы агента переноса генов не случайна» . Геномная биология и эволюция . 10 (1): 359–369. дои : 10.1093/gbe/evy005 . ПМЦ 5786225 . ПМИД 29325123 .
- ^ Кебатт М., Кристен М., Хармс А., Кернер Дж., Кристен Б., Дехио С. (июнь 2017 г.). «Агент переноса генов способствует развитию наиболее приспособленной субпопуляции бактериального патогена» . Клеточные системы . 4 (6): 611–621.e6. дои : 10.1016/j.cels.2017.05.011 . ПМК 5496983 . ПМИД 28624614 .
- ^ Мотро И., Ла Т., Беллгард М.И., Данн Д.С., Филлипс Н.Д., Хэмпсон DJ (март 2009 г.). «Идентификация генов, связанных с профагоподобными агентами переноса генов, у патогенных кишечных спирохет Brachyspira hyodysenteriae, Brachyspira pilosicoli и Brachyspira intermedia» . Ветеринарная микробиология . 134 (3–4): 340–5. дои : 10.1016/j.vetmic.2008.09.051 . ПМИД 18950961 .
- ^ Ланг, А.С.; Жахыбаева О; Битти, Джей Ти (11 июня 2012 г.). «Агенты переноса генов: фагоподобные элементы генетического обмена» . Обзоры природы. Микробиология . 10 (7): 472–82. дои : 10.1038/nrmicro2802 . ПМЦ 3626599 . ПМИД 22683880 .
- ^ Бертани Дж. (май 1999 г.). «Трансдукционно-подобный перенос генов в метаногене Methanococcus voltae» . Журнал бактериологии . 181 (10): 2992–3002. дои : 10.1128/JB.181.10.2992-3002.1999 . ПМК 93752 . ПМИД 10321998 .
- ^ Крупович, М; Фортерре, П; Бэмфорд, Д.Х. (2010). «Сравнительный анализ мозаичных геномов хвостатых вирусов архей и провирусов позволяет предположить общие закономерности архитектуры и сборки вирионов с хвостатыми вирусами бактерий». Журнал молекулярной биологии . 397 (1): 144–60. дои : 10.1016/j.jmb.2010.01.037 . ПМИД 20109464 .
- ^ Шерлок, Д; Леонг, JX; Фогг, PCM (1 декабря 2019 г.). «Идентификация малой терминазы первого агента переноса гена (GTA) у Rhodobacter capsulatus и ее роль в производстве GTA и упаковке ДНК» . Журнал вирусологии . 93 (23). дои : 10.1128/JVI.01328-19 . ПМК 6854486 . ПМИД 31534034 .