Деполяризующий преимпульс
( Деполяризующий преимпульс ДПП ) — это электрический стимул, который заставляет разность потенциалов, измеренную на нейрона, мембране становиться более положительной или менее отрицательной и предшествует другому электрическому стимулу. [1] ДПП могут относиться к стимулам напряжения или тока и используются для подавления нейронной активности. [1] избирательно возбуждают нейроны, [2] и повысить болевой порог, связанный с электрокожной стимуляцией . [3]
Биофизические механизмы
[ редактировать ]Модель Ходжкина – Хаксли
[ редактировать ]

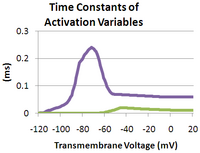
Типичные потенциалы действия инициируются потенциалзависимыми натриевыми каналами . По мере увеличения трансмембранного напряжения увеличивается вероятность того, что данный потенциалзависимый натриевый канал открыт, что обеспечивает приток Na. + ионы. Как только приток натрия становится больше, чем отток калия, петля положительной обратной связи поступления натрия замыкается и, таким образом, активируется потенциал действия.
В начале 1950-х годов д-р. Ходжкин и Хаксли провели эксперименты на гигантском аксоне кальмара и в процессе разработали модель ( модель Ходжкина-Хаксли ) проводимости натриевых каналов. Было обнаружено, что проводимость можно выразить как:

где — максимальная проводимость натрия, m — ворота активации, а h — ворота инактивации (оба ворота показаны на соседнем изображении). [4] Значения m и h варьируются от 0 до 1 в зависимости от трансмембранного потенциала.


По мере повышения трансмембранного потенциала значение m увеличивается, тем самым увеличивая вероятность того, что активационные ворота будут открыты. А по мере падения трансмембранного потенциала значение h увеличивается вместе с вероятностью того, что ворота инактивации будут открыты. Скорость изменения для ворот h намного медленнее, чем для ворот m , поэтому, если подпороговой стимуляции напряжением предшествует гиперполяризующий предимпульс, значение h может быть временно увеличено, позволяя нейрону запустить потенциал действия. .
И наоборот, если надпороговой вольтажной стимуляции предшествовать деполяризующим преимпульсом, значение h может быть временно уменьшено, что позволит затормозить нейрон. Иллюстрацию того, как может отличаться трансмембранная вольтажная реакция на надпороговый стимул в зависимости от наличия деполяризующего предымпульса, можно наблюдать на соседнем изображении.
Модель Ходжкина-Хаксли немного неточна, поскольку она не учитывает некоторые зависимости, например, ворота инактивации не должны иметь возможность закрыться, если ворота активации не открыты, а ворота инактивации, будучи закрытыми, расположены внутри клеточной мембраны , где они не могут быть закрыты. напрямую зависит от трансмембранного потенциала. [5] Однако эта модель полезна для получения высокого уровня понимания гиперполяризующих и деполяризующих предимпульсов. Деполяризация нейронов создает более вероятный исход возбуждения нейронов.
Потенциал-управляемый натриевый канал
[ редактировать ]
С тех пор как модель Ходжкина-Хаксли была впервые предложена в 1950-х годах, многое было изучено относительно структуры и функциональности потенциал-управляемых натриевых каналов . Хотя точная трехмерная структура натриевого канала остается неизвестной, его состав и функциональность отдельных компонентов определены. Потенциал-управляемые натриевые каналы представляют собой большие мультимерные комплексы, состоящие из одной субъединицы α и одной или нескольких субъединиц β, иллюстрацию которых можно наблюдать на соседнем изображении. α-субъединица складывается в четыре гомологичных домена, каждый из которых содержит шесть α- спиральных трансмембранных сегментов. [6] Сегменты S 4 каждого домена служат датчиками напряжения для активации. Каждый сегмент S 4 состоит из повторяющейся структуры, состоящей из одного положительно заряженного остатка и двух гидрофобных остатков, которые объединяются, образуя спиральную структуру. Когда канал деполяризуется, эти сегменты S 4 натриевого канала претерпевают конформационные изменения, которые расширяют спиральное расположение и открывают пору . В течение миллисекунд после открытия поры внутриклеточная петля, соединяющая домены III и IV, связывается с внутриклеточной порой канала, инактивируя канал. [7] Таким образом, обеспечивая деполяризующий предимпульс перед стимулом, существует большая вероятность того, что инактивирующие домены натриевых каналов связались с соответствующими порами, уменьшая вызванный стимулом приток натрия и влияние стимула.
Деполяризующие свойства предымпульса
[ редактировать ]Продолжительность ДПП
[ редактировать ]Связь между продолжительностью ДПП и рекрутированием нейронов следующая. Если длительность ДПП относительно невелика, т. е. значительно меньше 100 мкс, то порог возбуждения окружающих нервов будет снижаться, а не повышаться. Возможно, это результат деполяризации сегментов S 4 и небольшого времени, отведенного на инактивацию. Для ДПП длительного действия доменам III и IV натриевых каналов (обсуждаемым выше) дается больше времени для связывания с порами соответствующих каналов, поэтому наблюдается увеличение порогового тока с увеличением продолжительности ДПП. [2]
Амплитуда ДПП
[ редактировать ]По мере увеличения амплитуды ДПП от нуля до близкого к порогу результирующее увеличение порогового тока также будет расти. [2] [8] Это связано с тем, что более высокая амплитуда активирует больше натриевых каналов, что позволяет большему количеству каналов инактивироваться их доменами III и IV.
Межфазная задержка DPP
[ редактировать ]Увеличение задержки между ДПП и стимулом обеспечивает больше времени, в течение которого сегменты S4 натриевого канала могут закрыться, а домены III и IV могут отделиться от соответствующих пор. Таким образом, увеличение межфазной задержки ДПП уменьшит эффективное увеличение порогового тока, индуцируемое ДПП. [2]
Деполяризующие преимпульсные приложения
[ редактировать ]Повышение болевого порога
[ редактировать ]Одно из немедленных применений деполяризующих преимпульсов, исследованное докторами. Полетто и Ван Дорена заключается в повышении болевого порога, связанного с электрокожной стимуляцией. [3] Электрокожная стимуляция обладает большим потенциалом как механизм передачи дополнительной сенсорной информации. Следовательно, этот метод стимуляции может быть непосредственно применен к таким областям, как виртуальная реальность , сенсорная замена и сенсорное увеличение. Однако многие из этих применений требуют использования небольших решеток электродов , стимуляция которых часто бывает болезненной, что ограничивает полезность этой технологии. Экспериментальная установка, построенная докторами. Полетто и Ван Дорен, заключалась в следующем:

- 4 человека, каждый из которых продемонстрировал способность давать надежные оценки боли в предыдущих исследованиях.
- Средний палец левой руки опирается на дисковые электроды из полированной нержавеющей стали диаметром 1 мм.
- одиночный стимул состоял из серии трех одинаковых пар преимпульсов и стимул-импульсов, представленных в начале, середине и конце 1-секундного интервала.
- ширина преимпульса и стимулирующего импульса была согласована с длительностью 10 миллисекунд, так что пороговые значения были одинаковыми для обоих.
- использовали различные амплитуды предимпульса: 0%, 79%, 63%, 50%, 40% и 32%, чтобы изучить их влияние на испытываемую боль.
- опыты были проведены таким образом, что раздражитель без предимпульса был болезненным примерно в половине случаев; это было достигнуто путем увеличения и уменьшения амплитуды стимулирующего импульса для следующего испытания в зависимости от того, было ли оно отмечено как болезненное.
Их результаты показали, что предимпульс перед стимулирующим импульсом эффективно снижает вероятность возникновения боли из-за электрокожной стимуляции. Как ни удивительно, предимпульс в 32% от амплитуды стимулирующего импульса позволил почти вдвое снизить вероятность возникновения боли. Следовательно, в средах, в которых болевой порог трудно определить, может быть достаточно подать предимпульс относительно низкой амплитуды перед стимулом для достижения желаемого эффекта.
Порядок рекрутирования нервных волокон
[ редактировать ]
Было замечено, что помимо ингибирования нервной возбудимости предшествующий электрическому стимулу деполяризующий преимпульс позволяет инвертировать соотношение ток-расстояние, контролирующее рекрутирование нервных волокон , где соотношение ток-расстояние описывает, как изменяется пороговый ток для возбуждения нервных волокон. пропорциональна квадрату расстояния между нервным волокном и электродом. [2] Следовательно, если область воздействия деполяризующего предымпульса меньше, чем область действия стимула, то нервные волокна, расположенные ближе к электроду, будут испытывать большее увеличение порогового тока возбуждения. Таким образом, при наличии такого стимула нервные волокна, ближайшие к электроду, могут тормозиться, а те, которые находятся дальше, могут возбуждаться. [2] [9] Моделирование этой стимуляции, созданное докторами. Уоррена Гриля и Дж. Томаса Мортимера можно увидеть на соседнем изображении. Исходя из этого, стимул с двумя деполяризующими предимпульсами, каждый из которых имеет амплитуду немного ниже порогового тока (на момент доставки), должен увеличивать радиусы воздействия для инактивации близлежащих нервных волокон и возбуждения отдаленных нервных волокон.
Обычно нервные волокна большего диаметра могут активироваться одиночными импульсными стимулами меньшей интенсивности и, таким образом, могут быть задействованы легче. [1] [2] [10] Однако DPP продемонстрировали дополнительную возможность отменить этот порядок набора персонала. Поскольку электрические стимулы оказывают большее влияние на нервные волокна большего диаметра, ДПП, в свою очередь, вызывают большую степень инактивации натриевой проводимости внутри таких нервных волокон, поэтому нервные волокна меньшего диаметра будут иметь более низкий пороговый ток.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Гриль В.М., Мортимер Дж.Т. (1995). «Формы стимулов для избирательной нейронной стимуляции». Журнал IEEE Engineering in Medicine and Biology . 14 (4): 375–385. дои : 10.1109/51.395310 .
- ^ Jump up to: а б с д и ж г час Гриль В.М., Мортимер Дж.Т. (1997). «Инверсия соотношения ток-расстояние путем временной деполяризации». Транзакции IEEE по биомедицинской инженерии . 44 (1): 1–9. дои : 10.1109/10.553708 . ПМИД 9214779 . S2CID 7149611 .
- ^ Jump up to: а б Полетто CJ, Ван Дорен CL (2002). «Повышение болевого порога у людей с помощью деполяризующих преимпульсов». Транзакции IEEE по биомедицинской инженерии . 49 (10): 1221–1224. дои : 10.1109/TBME.2002.803563 . ПМИД 12374350 . S2CID 1817833 .
- ^ Ходжкин А.Л., Хаксли А.Ф. (1952). «Количественное описание ионных токов и его применение к проводимости и возбуждению в нервных мембранах» . Журнал физиологии . 117 (4): 500–544. doi : 10.1113/jphysicalol.1952.sp004764 . ПМЦ 1392413 . ПМИД 12991237 .
- ^ Даян, Питер; Эбботт, Ларри (2001). Теоретическая нейронаука . Кембридж, Массачусетс: MIT Press. ISBN 0-262-04199-5 .
- ^ Каттералл, Вашингтон (2000). «От ионных токов к молекулярным механизмам: структура и функции потенциалзависимых натриевых каналов» . Нейрон . 26 (1): 13–25. дои : 10.1016/S0896-6273(00)81133-2 . ПМИД 10798388 .
- ^ Ю Ф.Х., Каттералл, Вашингтон (2003). «Обзор семейства потенциал-управляемых натриевых каналов» . Геномная биология . 4 (3): 207. doi : 10.1186/gb-2003-4-3-207 . ПМК 153452 . ПМИД 12620097 .
- ^ Вукович А., Тосато М., Стрейк Дж.Дж. (2008). «Сравнительное исследование трех методов избирательной активации волокон блуждающего нерва по диаметру: анодная блокада, деполяризующие преимпульсы и медленно нарастающие импульсы». Журнал нейронной инженерии . 5 (3): 275–286. Бибкод : 2008JNEng...5..275В . дои : 10.1088/1741-2560/5/3/002 . ПМИД 18566504 . S2CID 7250314 .
- ^ Деурлоо К.Е., Хольшаймер Дж., Бергвелд П. (2001). «Влияние подпороговых предимпульсов на порядок рекрутирования в нервном стволе анализируется с помощью простой и реалистичной модели объемного проводника» (PDF) . Биологическая кибернетика . 85 (4): 281–291. дои : 10.1007/s004220100253 . ПМИД 11592625 . S2CID 8098294 .
- ^ Хеннингс К., Камавуако Е.Н., Фарина Д. (2007). «Порядок рекрутирования электрически активированных мотонейронов, исследованный с помощью новой техники столкновений». Клиническая нейрофизиология . 118 (2): 283–291. дои : 10.1016/j.clinph.2006.10.017 . ПМИД 17174598 . S2CID 25968304 .