Сордария фимикола
| Сордария фимикола | |
|---|---|
 | |
| Sordaria fimicola перитеций | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | Грибы |
| Разделение: | Аскомикота |
| Сорт: | Сордариомицеты |
| Заказ: | Сордариалес |
| Семья: | Сордариевые |
| Род: | Сордария |
| Разновидность: | С. фимикола |
| Биномиальное имя | |
| Сордария фимикола | |

Sordaria fimicola — вид микроскопических грибов . Обычно он встречается в фекалиях травоядных животных . Sordaria fimicola часто используется в лабораториях вводной биологии и микологии , поскольку ее легко выращивать на питательном агаре в чашечных культурах. Род Sordaria , тесно связанный с Neurospora и Podospora , относится к большому классу Sordariomycetes , или колбчатых грибов. Естественная среда обитания трех видов Sordaria , ставших основными объектами генетических исследований, — навоз травоядных животных. Вид S. fimicola обычен и распространен по всему миру. Виды Sordaria сходны морфологически , образуя черные перитеции, содержащие аски с восемью темными аскоспорами, расположенными линейно. Эти виды имеют ряд общих характеристик, которые полезны для генетических исследований. Все они имеют короткий жизненный цикл , обычно 7–12 дней, и легко выращиваются в культуре. Большинство видов самоплодны, и каждый штамм изогенен. Все виды мутантов легко индуцируются и легко получаются с помощью мутантов определенного цвета аскоспор. Эти визуальные мутанты помогают тетрадный анализ , особенно при анализе внутригенной рекомбинации. [1]
Самая распространенная форма S. fimicola — темно-коричневая. Некоторые мутанты имеют серый или коричневый цвет. Обычный эксперимент для вводного лабораторного занятия по биологии - скрещивание одного из мутантных типов с диким типом и наблюдение за соотношением окраски у потомства. Этот эксперимент иллюстрирует концепции генетической наследственности в гаплоидном организме. Восемь аскоспор образуются внутри сумки . Кабачки сордарии могут дать нам информацию о кроссинговере во время мейоза. Если кроссинговера не происходит, получается узор 4:4: четыре черные споры и четыре коричневые споры, выстроенные в линию. Если пересечение действительно происходит, то виден паттерн 2:2:2:2 или паттерн 2:4:2.
Еще одно распространенное лабораторное применение — наблюдение мейоза и митоза в плодовых телах, называемых перитециями. Интересной особенностью S. fimicola является то, что ее плодовое тело фототрофно . Таким образом, по мере роста стебель будет наклоняться к источнику света, а когда мешочек лопнет, споры вылетят в сторону света.
Таксономия
[ редактировать ]Исследования по обновлению таксономии грибов Sordariomycete продолжаются, а положение Sordaria fimicola в таксономии Sordariomycetes исследуется и обновляется. Сордариомицеты известны как колбчатые грибы, поскольку для них характерны колбовидные перитеции и единичные аски. [2] Филогенетические исследования частичных цепей рибосомальной ДНК 18s пролили свет на филогению сордариомицетов. [3] Морфологические особенности, характеризующие Sordariaceae, включают дифференцировку оболочки гифы, окружающей аскогоний, на слои периферической стенки и псевдопаренхиматозный центр. Широкие парафизы, состоящие из нежных многоядерных клеток, возникают из клеток центра и полностью заполняют перитеций, придавливая оставшиеся псевдопаренхиматозные клетки к перитециальной стенке. [4] S. fimicola отличается от других изученных видов Sordariaceae агрегацией аскогенных клеток с образованием плацентоподобной массы в основании центра. Следовательно, аски возникают в кластере, а не в однородном пристеночном слое. [4]
Грибковая морфология
[ редактировать ]Sordaria fimicola — аскомицетный гриб, который хорошо растет на питательном агаре, навозе и в разлагающихся веществах почвы. Как аскоймцет, он имеет половую форму телеоморфа и бесполую форму анаморфа. S. fimicola обычно повторяет жизненный цикл облигатного навозного гриба; половое размножение, обязательное для навоза травоядных после прохождения через желудочно-кишечный тракт травоядных. После мейоза навоза аскоспоры выделяются и прикрепляются к поверхности растения, где, как полагают, они остаются эпифиллярными. [5] [6] У S. fimicola растут перегородчатые гифы, которые на макроуровне выглядят как небольшие темно-коричневые волокна, образующие пушистый лист над субстратом. Маленькие черные пятна образуются при половом размножении, поскольку эти пятна являются местом расположения аски. Кроме того, существуют желто-коричневые и серые мутантные штаммы S. fimicola . [5] S. fimicola лучше всего растет на богатых углеродом субстратах, но ей также необходим доступ к азоту. Исследования показали, что S. fimicola лучше всего растет при соотношении углерода: азота от 5: 1 до 10: 1, поскольку S. fimicola, выращенная в этих условиях, производит заметно больше перитеция, чем те, у кого более высокое соотношение углерода или азота. S. fimicola хорошо растет в лабораторных условиях, поэтому его часто используют в лабораториях вводной биологии, а также в качестве модельного организма для исследований. S. fimicola также является хорошим инструментом для обучения мейозу, поскольку он быстро производит диплоидный перитеций, который подвергается мейозу с образованием аский с упорядоченными линейными тетрадами гаплоидных аскоспор. [3]
Экология
[ редактировать ]Хотя традиционно считалось, что S. fimicola растет на навозе травоядных животных или в разлагающихся растительных веществах, также было обнаружено, что она растет в симбиозе с некоторыми растениями. Новые исследования показывают, что S. fimicola является факультативным навозным грибом, и было обнаружено, что S. fimicola растет среди корней райграса и пшеницы. [7] В стерилизованной и нестерилизованной почве S. fimicola способствовала росту хозяина и предотвращала смертность. Исследования показывают, что S. fimicola в культурах картофельной декстрозы способна продуцировать триаконтанол и индол-3-карбоксальдегид , оба из которых обладают антибактериальными свойствами, которые могут помочь предотвратить заболевание хозяина. [8]
Также было показано, что S. fimicola подавляет рост других видов грибов, патогенных для растений, включая Pestalotiopsis guepinii , Colletotrichum capsici , Curvularia lunata [ Cochliobolus lunatus ], Alternaria alternata и Fusarium oxysporum . [9]
Также было показано, что S. fimicola отрицательно влияет на здоровье некоторых растений, на которых она растет. Когда S. fimicola была впервые выделена из кукурузы, исследователи полагали, что это, скорее всего, условно-патогенный патоген. На здоровых растениях кукурузы в лаборатории не росла S. fimicola , что указывает на то, что здоровые растения кукурузы могут исключить ее. S. fimicola действительно колонизировала здоровый B. tectorum , что привело к снижению плодовитости и замедлению роста. [7]
S. fimicola также фототропен, и его реакция двоякая: 1) свет влияет на количество плодовых тел, вырабатываемых грибом, и 2) также влияет направление роста плодовых тел. Этот ответ, вероятно, опосредован специфическим грибковым рецептором, называемым фоторецептором белого воротника-1 S. fimicola (SfWC-1). Другие виды фототропных грибов имеют гомологичные рецепторы со схожими рецепторными доменами. [10] S. fimicola с нефункциональным мутантным рецептором SfWC-1 имел запоздалое и менее выраженное формирование плодовых тел, был дефектен по фототропизму перитециальных клювов и не имел паттерна зональности плодовых тел по сравнению с диким типом. [10]
Конверсия генов
[ редактировать ]Каждый отдельный мейоз генерирует четыре гаплоидных продукта, а после еще одного раунда митоза образуются восемь продуктов, и все они сохраняются в виде гаплоидных спор внутри мешкообразной сумки ( pl. asci). Сохранение продуктов индивидуального мейоза в отдельной сумке облегчило некоторые виды генетического анализа, в частности анализ молекулярного механизма генетической рекомбинации . Когда штамм дикого типа (+) скрещивают с мутантным штаммом (m), обычно каждая аска содержит четыре + и четыре m споры. Однако было обнаружено, что при низкой частоте некоторые аски имели соотношения, отличающиеся от ожидаемых 4+:4m (например, 6+:2m или 2+:6m или даже 5+:3m или 3+:5m). В этих случаях оказалось, что ген m был преобразован в ген + или наоборот. Это явление было названо « генной конверсией ». Генная конверсия была впервые обнаружена у S. fimicola в 1951 году Линдсеем Оливом и окончательно охарактеризована им в 1959 году. [11] Олив считала, что эти события генной конверсии являются результатом «транс-репликации, при которой локус копируется больше, чем обычно, количество раз во время репликации в профазе мейоза». С тех пор было проведено множество исследований феномена конверсии генов на S. fimicola и других организмах, особенно на других аскомицетах [см. обзор Whitehouse [12] (1982)]. Попытки понять конверсию генов на молекулярном уровне позволили понять механизм и адаптивную функцию мейотической рекомбинации, которая, в свою очередь, влияет на адаптивную функцию полового размножения.
Изображения
[ редактировать ] Спаривание колоний дикого типа (темно-коричневые) и мутантных (коричневых) Sordaria fimicola . колоний
Спаривание колоний дикого типа (темно-коричневые) и мутантных (коричневых) Sordaria fimicola . колоний Sordaria fimicola peritecia с аскоспорами как мутантного, так и дикого типа.
Sordaria fimicola peritecia с аскоспорами как мутантного, так и дикого типа.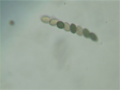 Sordaria fimicola ascus с необычным рисунком 2:1:1:1:1:2.
Sordaria fimicola ascus с необычным рисунком 2:1:1:1:1:2.



Ссылки
[ редактировать ]- ^ Алексопулос CJ, Мимс CW, Блэквелл М (1996). Вводная микология . Джон Уайли и сыновья. п. 361. ИСБН 0-471-52229-5 .
- ^ Хонгсанан, Синанг; Махараччикумбура, Саджива С.Н.; Хайд, Кевин Д.; Самаракун, Милан К.; Дживон, Раджеш; Чжао, Ци; Аль-Сади, Абдулла М.; Бахкали, Али Х. (1 мая 2017 г.). «Обновленная филогения сордариомицетов, основанная на данных филогенетики и молекулярных часов» . Грибное разнообразие . 84 (1): 25–41. дои : 10.1007/s13225-017-0384-2 . ISSN 1878-9129 . S2CID 5062936 .
- ^ Jump up to: а б Дуонг, Лам (2004). «Emarcea castanopsidicola gen. et sp. nov. из Таиланда, новый таксон ксилярийных, основанный на морфологии и последовательностях ДНК». Исследования по микологии . 50 : 253–260.
- ^ Jump up to: а б Май, Шинг Х. (1977). «Морфологические исследования Sordaria Fimicola и Gelasinospora Longispora» . Американский журнал ботаники . 64 (5): 489–495. дои : 10.1002/j.1537-2197.1977.tb11884.x . ISSN 1537-2197 .
- ^ Jump up to: а б Алексопулус, CJ (1996). Вводная микология . Джон Уайли и сыновья. п. 361.
- ^ «Жизненный цикл Sordaria Fimicola» . Наука . Проверено 20 мая 2021 г.
- ^ Jump up to: а б Ньюкомб, Джордж; Кэмпбелл, Джейсон; Гриффит, Дэвид; Бэйнс, Мелисса; Лаунбо, Карен; Пендлтон, Розмари (3 февраля 2016 г.). Альбректсен, Бенедикт Рибер (ред.). «Возврат к жизненному циклу навозных грибов, включая Sordaria fimicola» . ПЛОС ОДИН . 11 (2): e0147425. Бибкод : 2016PLoSO..1147425N . дои : 10.1371/journal.pone.0147425 . ISSN 1932-6203 . ПМЦ 4739622 . ПМИД 26839959 .
- ^ Деван, ММ; Гисалбертиб, Эль; Роуленд, К.; Сиваситхампарам, К. (апрель 1994 г.). «Снижение симптомов поражения проростков пшеницы и райграса почвенным грибом Sordaria fimicola» . Прикладная экология почв . 1 (1): 45–51. дои : 10.1016/0929-1393(94)90022-1 . ISSN 0929-1393 .
- ^ Джемджитт, О. (2007). «Копрофильные грибы и антагонистическое действие Sordaria fimicola против фитопатогенных грибов in vitro» . Растения : 593–600.
- ^ Jump up to: а б Кробанан, Кулсумпун; Лян, Сюн-Вун; Чиу, Хо-Чен; Шен, Вэй-Чан (12 апреля 2019 г.). Дружинина, Ирина С. (ред.). «Ген фоторецептора синего света Sfwc-1 регулирует фототропную реакцию и развитие плодовых тел у гомоталлического аскомицета Sordaria fimicola» . Прикладная и экологическая микробиология . 85 (12): e02206–18, /aem/85/12/AEM.02206–18.atom. Бибкод : 2019ApEnM..85E2206K . дои : 10.1128/АЕМ.02206-18 . ISSN 0099-2240 . ПМК 6544823 . ПМИД 30979837 .
- ^ Олив Л.С. (1959). «Аберрантные тетрады в Сордарии Фимикола» . Учеб. Натл. акад. наук. США . 45 (5): 727–32. Бибкод : 1959PNAS...45..727O . дои : 10.1073/pnas.45.5.727 . ПМК 222623 . ПМИД 16590433 .
- ^ Гарольд Л.К. Уайтхаус. 1982. Генетическая рекомбинация. Нью-Йорк: Уайли ISBN 978-0471102052
| Базы данных органов управления : Национальные |
|---|