Клеточное обезвоживание
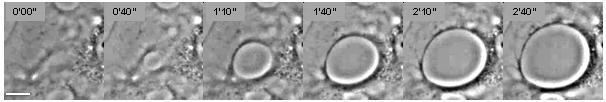
Клеточное расслаивание относится к процессу зарождения и расширения туннелей макроапертур трансэндотелиальных клеток (ТЕМ) в эндотелиальных клетках (рис. 1). [1] Это явление аналогично зарождению и росту сухих пятен в вязких жидкостях, растекающихся по несмачиваемой подложке (рис. 2). [2] Осушение клеток запускается несколькими белковыми токсинами патогенных бактерий , в частности, EDIN-подобными факторами Staphylococcus aureus и Clostridium botulinum , а также отечным токсином Bacillus anthracis . [3] [4] ТЕМ образуются в ответ на разрыв физических связей цитоскелета через цитоплазму вследствие ингибирования пути RhoA/ROCK или индукции потока циклического АМФ ( цАМФ широкой сигнальной молекулы ) . [4] [5]
Физика, лежащая в основе клеточного обезвоживания
[ редактировать ]
Явление клеточного обезвоживания можно интерпретировать с помощью физического моделирования (рис. 2). [6] Движущей силой, ответственной за спонтанное образование ТЕМ-туннелей и их открытие, является мембраны натяжение , возникающее в результате распластывания клеток за счет релаксации актомиозина. В отличие от обезвоживания жидкостью, ТЭМ достигают максимального диаметра, при котором движущая сила уравновешивается силой сопротивления, которая возникает вдоль краев ТЭМ (рис. 2). Эта сила сопротивления называется натяжением линии и не характеризуется на молекулярном уровне.
Физические параметры
[ редактировать ]Движущие силы, тянущие туннель радиусом R , как показано на рисунке 2. Здесь тяга происходит из-за натяжения клеточной мембраны (σ), которому частично противодействует натяжение линии вокруг туннеля ( T ). В этих условиях чистая движущая сила ( F D ) состоит из двух вкладов:

Осушение продолжается, если F D >0.
Натяжение мембраны ( σ ) зависит от радиуса туннеля R . Туннельное увеличение размера расслабляет мембрану , вызывая уменьшение натяжения мембраны, как описано законом Хелфриха .
Линейное натяжение ( T ) соответствует силе сопротивления вдоль края туннеля, которая противодействует натяжению мембраны и ограничивает высыхание. Это напряжение линии может иметь физические и молекулярные компоненты.
Ссылки
[ редактировать ]- ^ Лемичес, Э. (2012). «Динамика трансклеточного туннеля: контроль клеточного обезвоживания с помощью сократимости актомиозина и белков I-BAR». Биология клетки . 105 (3): 109–117. дои : 10.1111/boc.201200063 . ПМИД 23189935 . S2CID 31452113 .
- ^ Де Женн, П.-Ж. (2004). Капиллярность и явления смачивания . Нью-Йорк: Спрингер. ISBN 978-0387005928 .
- ^ Бойер, Л. (2006). «Индукция временных макроапертур в эндотелиальных клетках посредством ингибирования RhoA факторами Staphylococcus aureus» . Журнал клеточной биологии . 173 (5): 809–819. дои : 10.1083/jcb.200509009 . ПМК 2063895 . ПМИД 16754962 .
- ^ Перейти обратно: а б Маддугода, член парламента (2011). «Передача сигналов цАМФ с помощью токсина отека сибирской язвы индуцирует туннели трансэндотелиальных клеток, которые закрываются с помощью MIM посредством полимеризации актина, управляемой Arp2/3» . Клетка-хозяин и микроб . 10 (5): 464–474. дои : 10.1016/j.chom.2011.09.014 . ПМИД 22100162 .
- ^ Кай, Ю. (2010). «Цитоскелетная когерентность требует сократительной способности миозина-IIA» . Журнал клеточной науки . 123 (3): 413–423. дои : 10.1242/jcs.058297 . ПМК 2816186 . ПМИД 20067993 .
- ^ Гонсалес-Родригес, Д. (2012). «Клеточное обезвоживание: открытие макроапертур в эндотелиальных клетках» . Письма о физических отзывах . 108 (21): 218105. doi : 10.1103/PhysRevLett.108.218105 . ПМИД 23003307 .