Виривор
Virivore (эквивалент virovore ) происходит от английского префикса viro, означающего вирус, происходящего от латинского слова, обозначающего яд. [ нужна ссылка ] и суффикс -vore от латинского слова vorare , означающего есть или пожирать; [1] следовательно, живоядное животное — это организм, потребляющий вирусы. Вириворность — это хорошо описанный процесс, при котором организмы, в первую очередь гетеротрофные протисты , [2] [3] [4] [5] [6] [7] но и некоторые многоклеточные животные [8] [9] потреблять вирусы.
Вирусы считаются главными хищниками морской среды, поскольку они могут лизировать микробы и выделять питательные вещества (т. е. вирусный шунт ). Вирусы также играют важную роль в структурировании микробных трофических взаимоотношений и регуляции потока углерода. [10] [11]
Открытие
[ редактировать ]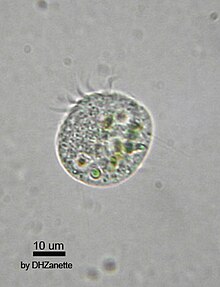
Первым описанным вировором был небольшой морской жгутиконосец, который, как было показано, способен заглатывать и переваривать вирусные частицы. [2] Впоследствии многочисленные исследования прямо и косвенно продемонстрировали потребление вирионов. [3] [4] [5] [6] [7] [8] [9] В 2022 году Делонг и др. показали, что в течение двух дней инфузории Halteria и Paramecium уменьшали количество единиц, образующих хлоровирусные бляшки, на два порядка, подтверждая идею о том, что питательные вещества передаются от вирусов к потребителям. [7]

Более того, популяция галтерий росла, когда хлоровирус был единственным источником питания, и минимально росла в отсутствие хлоровируса. [7] Однако популяция Paramecium не отличалась по росту при кормлении хлоровирусами по сравнению с контрольной группой. Поскольку размер популяции Paramecium оставался постоянным в присутствии только хлорвирусов, это указывало на то, что Paramecium способен поддерживать размер своей популяции, но не расти, используя хлоровирус в качестве единственного источника углерода. Эти данные показали, что некоторые травоядные могут расти на вирусах, но это касается не всех травоядных. Было подсчитано, что Халтерия потребляла от 10 000 до 1 000 000 вирусов в день. Известно, что мелкие протисты, такие как Halteria и Paramecium , поедаются зоопланктоном, что указывает на движение вирусной энергии и вещества вверх по водной пищевой сети. Это противоречит идее о том, что вирусный шунт ограничивает движение энергии вверх по пищевым сетям, прекращая взаимодействие травоядных и микробов. Количество передаваемой энергии и вещества будет зависеть от размера вириона и содержания питательных веществ, которые будут варьироваться в зависимости от штамма.
Биогеохимическое воздействие
[ редактировать ]
Вирусы являются наиболее распространенными биологическими объектами в Мировом океане. [12] [13] [14] Жизненный цикл литического вируса является важным процессом в Мировом океане, обеспечивающим круговорот растворенного органического вещества и твердых частиц органического вещества , т.е. вирусный шунт . [15] [12] [13] Сами вирусные частицы также составляют значительную долю богатых азотом и фосфором частиц в пуле растворенных органических веществ, поскольку они состоят из липидов, аминокислот, нуклеиновых кислот и, вероятно, углерода, включенного из клеток-хозяев. [12] [13] Считается, что вирусы могут дополнять рацион травоядных при проглатывании, и микроб не заражается. [14]
Общий выпас вирусов широко распространен во всей морской среде, при этом уровень выпаса достигает 90,3 мл. −1 день −1 . [12] Когда присутствуют и бактерии, и вирусы, вирусы могут поглощаться со скоростью, сравнимой с бактериями. [2]
Используя Oikopleura dioica и Equid Alphaherpesvirus 1 (EhV) в качестве модели, ученые оценили питательную пользу вирусов; [12]
- 24.2 C индивидуальный −1 день −1 [12]
- 2,8 из N особей −1 день −1 [12]
- 0,2 П индивидуально −1 день −1 [12]
Предполагается, что у более мелких травоядных вирусы потенциально могут оказывать более существенное влияние на питание хозяина. [12] Например, у нанофлагеллят предполагаемый вклад составляет 9% углерода, 14% азота и 28% фосфора. [12]
В то время как более мелкие бактерии являются идеальным источником пищи для травоядных из-за их размера и содержания углерода, вирусы маленькие, неподвижные и чрезвычайно многочисленны для травоядных, что делает их альтернативным выбором питания. [14] Обычным травоядным животным, чтобы получить такое же количество углерода из вирусов, как и из бактерий, им нужно будет потреблять в 1000 раз больше вирусов. [14] Это не делает вирусы идеальным источником углерода для травоядных. Однако, помимо роста, у потребления вирусов есть и другие преимущества. Исследования показывают, что переваренные вирусные частицы выделяют аминокислоты, которые травоядные могут затем использовать во время собственного синтеза полипептидов. [14]
Вирусная зачистка
[ редактировать ]Трофические взаимодействия между травоядными, бактериями и вирусами важны для регулирования круговорота питательных веществ и органических веществ. [14] Вирусное распространение — это механизм, при котором травоядные животные возвращают углерод обратно в классическую пищевую сеть, поглощая вирусные частицы. [12] Заражение клеток-хозяев приводит к высвобождению вирусного потомства, которое впоследствии потребляется травоядными. [12] Затем травоядные потребляются более высокими трофическими организмами, в результате чего углерод вирусов возвращается обратно в классическую пищевую сеть и на более высокие трофические уровни. [12]
На распространение вируса могут влиять многие факторы, такие как размер и численность вирусных частиц. [12] Размер вируса будет влиять на элементный состав вирусных частиц. [13] Например, вирус с более крупным капсидом будет вносить больше углерода, а вирусы с более крупными геномами будут вносить больше азота и фосфора в результате увеличения количества нуклеиновых кислот. [13] Кроме того, влияние вирусного нашествия может быть более значительным, если учитывать также травоядных, охотящихся на бактерии, зараженные вирусами. [14] В целом, потребляя бактерии и вирусы, травоядные играют важную роль в круговороте углерода. [14]
Вирусный выпас
[ редактировать ]Потребление вирусов во многом зависит от пищевого поведения организма.
Фильтрационная подача
[ редактировать ]Фильтрующее кормление – это вид суспензионного кормления. [16] Фильтраторы обычно активно захватывают отдельные частицы пищи на ресничках, волосках, слизи или других структурах. [16] Исследователи использовали Салпингеоку в качестве модельного фильтратора, чтобы наблюдать за изменением численности вируса. Salpingeoca производят лорику , которая помогает им прикрепиться к субстрату. [14] У них также есть один жгутик, создающий поток воды, который переносит к ним мелкие частицы, где крошечные ложноножки поглощают частицы добычи. [14] Когда вирусы инкубировались совместно с сальпингеокой, численность вирусов неуклонно снижалась в течение 90 дней, что показывает, что фильтрующее кормление является эффективным механизмом питания вирусов. [14]
Пасение на частицах отложений
[ редактировать ]Пасущиеся перемещаются по поверхности, собирая и поглощая пищу на ходу. [16] Исследователи использовали Thaumatomonascoliensis в качестве модели травоядного, чтобы наблюдать изменения в численности вирусов. [14] T.coloniensis скользит по субстрату и образует филоподии , которые используются для поглощения частиц, связанных с субстратом. [14] В течение 90 дней численность вируса неуклонно снижалась при совместной инкубации с T.coliensis, что показывает, что выпас является эффективным механизмом питания вирусами. [14]
Хищное кормление
[ редактировать ]Кормление хищников — это форма активного кормления, при которой организм ищет добычу. [14] Исследователи использовали Goniomonas truncata в качестве модели хищного питания. [14] G. truncata — это криптомонада, у которой есть два жгутика, которые используются для плавания близко к субстрату в поисках пищи, и у них есть вакуоли, помогающие усваивать пищу. [14] В присутствии G. truncata численность вируса существенно не снизилась в течение 90 дней. [14] Однако это не исключает возможности того, что вирусные частицы поглощаются, а затем выбрасываются обратно в окружающую среду. [14] Эти данные показывают, что кормление хищников, возможно, не является методом выпаса вируса, но может иметь и другие экологические последствия с точки зрения передачи вируса.
Выборочный выпас
[ редактировать ]Выпас вирусов различается в зависимости от вируса, и поэтому он подлежит избирательному кормлению. Жгутиконосцы способны поглощать множество вирусов разного размера, при этом самые маленькие вирусы имеют наименьшую скорость поглощения. [2] существует огромное Среди морских вирусов разнообразие , включая размер, форму, морфологию и поверхностный заряд, которые могут влиять на отбор и, следовательно, на скорость проглатывания. [2] Кроме того, скорость переваривания разных вирусов одними и теми же жгутиковыми была различной. Это подразумевает отбор при выпасе вирусов. [2] Например, значительные различия в удалении вируса с помощью Tetrahymenapyriformis наблюдались, когда протист одновременно инкубировали с 13 различными типами вирусов. [17] Кроме того, скорость удаления конкретных вирусов сохранялась, когда протистов одновременно инкубировали с несколькими вирусами. [17] T.pyriformis смогли идентифицировать вирусы как пищу, что определяет их перемещение и потребление одних вирусов по сравнению с другими, что подтверждает идею о том, что некоторые протисты способны к выборочному выпасу. [17]
Влияние вирусной инфекции на выпас
[ редактировать ]Вирусы обладают способностью влиять на выедание клеток-хозяев во время заражения, показывая, что вирусная инфекция играет роль в избирательном выпасе. [18] [19]
Веслоногие ракообразные являются ключевым звеном в морских пищевых цепях, поскольку они связывают первичное и вторичное производство с более высокими трофическими уровнями. [18] Когда фитопланктон Emiliania huxleyi был инфицирован кокколитовирусом EhV -86, поглощение инфицированных клеток каланоидным веслоногим раком Acartia тонна было значительно снижено по сравнению с неинфицированными клетками, что указывает на селективное выедание инфицированных клеток. [18] Эти результаты показывают, что вирусные инфекции уменьшают выпас скота и потенциально могут снизить эффективность пищевой сети, удерживая углерод внутри вирусной шунтово - микробной петли и подавляя движение углерода на более высокие трофические уровни. [18] Это подчеркивает важность вирусного распространения углерода для перевода углерода на более высокие трофические уровни.
Напротив, Oxyris marina отдавала предпочтение выпасу зараженной вирусом Emiliania huxleyi. [19] Предполагается, что предпочтение инфицированных клеток перед неинфицированными обусловлено физиологическими изменениями или изменением размеров клетки-хозяина. [19] O. marina предпочитают питаться более крупными клетками, поскольку они потенциально могут получить от них большую питательную ценность по сравнению с клетками меньшего размера, для потребления которых потребуется такое же количество энергии. [19] Зараженные E. huxleyi имеют увеличенный размер клеток по сравнению с незараженными, что делает их идеальной добычей для O. marina. [19] Инфицированные E. huxleyi также могут быть выбраны по вкусовым качествам в результате физиологических изменений во время инфекции. [19] Например, инфицированные клетки будут иметь более высокое содержание нуклеиновых кислот по сравнению с неинфицированными клетками, что может улучшить прирост питательных веществ для травоядных. [19] Кроме того, пастбищная активность O. marina связана с добычей с более низкой активностью диметилсульфониопропионат- лиазы (DMSP-лиазы), поскольку они будут производить меньше потенциально токсичного соединения акрилата . [19] Зараженные вирусом E. huxleyi демонстрируют пониженный уровень активности DMSP-лиазы, что делает их привлекательными для O. marina за счет снижения воздействия на них вредных соединений. [19] Наконец, химические сигналы, такие как выделение диметилсульфида и перекиси водорода во время инфекции, вероятно, создают градиент, облегчающий O. marina обнаружение зараженного E. huxleyi. [19] Преимущественное выедание инфицированных клеток сделает углерод доступным для более высоких трофических уровней за счет его секвестрации в форме частиц. [19]
В целом, выпас вирусных частиц и инфицированных вирусом клеток подлежит выборочному выпасу.
Экологическое значение
[ редактировать ]Исследования показали, что вирусы могут быть проглочены и переварены или проглочены и выброшены обратно в окружающую среду травоядными. [14] [20] Наблюдение о том, что травоядные животные потенциально могут выпустить вирусы обратно в окружающую среду после проглатывания, может иметь значительные экологические последствия. [20]
Способ передачи
[ редактировать ]
Проглатывание и выброс вирусов могут способствовать передаче и распространению вирусов в морской среде. [20] Используя веслоногих ракообразных в качестве модельного вектора передачи и EhV в качестве модельного вируса, Frada et al. определили потенциальный механизм распространения вируса в морской среде. [20]
Частицы EhV могут потребляться веслоногими ракообразными либо в виде отдельных частиц вириона, либо посредством инфицирования клетки-хозяина (в данном случае инфицированной Emiliania huxleyi). [20] Когда инфицированный E. huxleyi инкубировали совместно с копеподами, фекальные гранулы, вырабатываемые копеподами, содержали в среднем 4500 ЭВ на гранулу. [20] Эти содержащие вирион осадки затем совместно инкубировали со свежей культурой E. huxleyi и наблюдали быстрый вирусопосредованный лизис клеток-хозяев. [20] Когда только частицы EhV инкубировались совместно с копеподами, т.е. без E. huxleyi, собранные фекальные частицы не содержали никаких частиц вириона. [20] Однако когда они кормили веслоногих ракообразных EhV и Thalassiosira weissflogii , диатомовых водорослей, находящихся за пределами диапазона хозяев EhV, собранные фекальные гранулы содержали 200 EhV на гранулу. [20] Эти осадки при совместной инкубации со свежей культурой E. huxleyi были высокоинфекционными и полностью уничтожали культуру. [20] Отсутствие частиц вириона в фекальных гранулах, полученных в результате инкубации единственного EhV, подтверждает идею о том, что травоядные животные избирательно поглощают вирусы. EhV все еще может поглощаться веслоногими ракообразными при инфицировании клеток-хозяев и при наличии идеального источника пищи. [20] Поскольку численность вирусов следует за численностью бактерий, маловероятно, что появится морская среда, в которой вирусы будут единственным источником питательных веществ для травоядных. [21]
Результаты этого эксперимента имеют значительные экологические последствия. Веслоногие ракообразные способны перемещаться вверх и вниз по толще воды, а также мигрировать на короткие расстояния между зонами питания. [20] [22] В частности, что касается копепод и EhV, перемещение копепод может переносить вирусы в новые и незараженные популяции E. huxleyi, способствуя гибели цветения. [20] Кроме того, фекальные гранулы могут опускаться из смешанного слоя в более глубокие части океана, где они могут ассимилироваться несколько раз. [22] Эти два сценария представляют собой потенциальные механизмы проникновения вирусов в новую морскую среду.
Организмы, не являющиеся хозяевами
[ редактировать ]
Пасущиеся – не единственные организмы, способные удалять вирусы из толщи воды. Организмы, не являющиеся хозяевами, такие как анемоны , личинки полихет, асцидии , крабы, моллюски, устрицы и губки , способны значительно снизить численность вируса. [23] Было обнаружено, что губки обладают наибольшим потенциалом для удаления вирусов. [23]
Метод, при котором организмы, не являющиеся хозяином, нарушают контакт вируса с хозяином, известен как интерференция передачи. [23] Организмы, не являющиеся хозяевами, могут оказывать либо прямое воздействие, удаляя организмы-хозяева, либо косвенное, удаляя вирусы. [23] Эти механизмы вызывают снижение частоты контактов вируса с хозяином, что может существенно повлиять на динамику местных микробных популяций. [23]
Организмы, не являющиеся хозяевами, способны удалять вирусы со скоростью, сравнимой с естественными частицами пищи, бактериальными клетками и клетками водорослей, что выше по сравнению с травоядными животными, у которых уровень очистки от вирусов составляет около 4%. [2] [23] В регионах с высокой плотностью губок, таких как прибрежные и тропические регионы, вполне вероятно, что скорость удаления вируса недооценена. [23] Эффективное удаление вирусов, вероятно, будет иметь глобальные экологические последствия, которые остались незамеченными. [23]
Ссылки
[ редактировать ]- ^ Правительство Канады PS (06 декабря 2019 г.). «-vore – Советы по написанию Plus – Инструменты для письма – Ресурсы языкового портала Канады – Canada.ca» . www.noslangues-ourlanguages.gc.ca . Проверено 13 апреля 2023 г.
- ^ Jump up to: а б с д и ж г Гонсалес Дж. М., Саттл, Калифорния (1993). «Поедание морскими нанофлагеллятами вирусов и частиц размером с вирус: проглатывание и переваривание» (PDF) . Серия «Прогресс в области морской экологии» . 94 (1): 1–10. Бибкод : 1993MEPS...94....1G . дои : 10.3354/meps094001 .
- ^ Jump up to: а б Гоуинг ММ (1993). «Крупные вирусоподобные частицы из вакуолей радиолярий феодарий и других морских образцов» (PDF) . Серия «Прогресс в области морской экологии» . 101 : 33–42. Бибкод : 1993MEPS..101...33G . дои : 10.3354/meps101033 .
- ^ Jump up to: а б Денг Л., Краусс С., Файхтмайер Дж., Хофманн Р., Арндт Х., Гриблер С. (2014). «Выпас гетеротрофных жгутиконосцев вирусами обусловлен пищевым поведением» . Отчеты по экологической микробиологии . 6 (4): 325–330. Бибкод : 2014EnvMR...6..325D . дои : 10.1111/1758-2229.12119 . ПМИД 24992530 .
- ^ Jump up to: а б Браун Дж.М., Лабонте Дж.М., Браун Дж., Рекорд Н.Р., Поултон Н.Дж., Серацкий М.Е., Логарес Р., Степанаускас Р. (2020). «Одноклеточная геномика выявила вирусы, потребляемые морскими протистами» . Границы микробиологии . 11 : 524828. doi : 10.3389/fmicb.2020.524828 . ПМЦ 7541821 . ПМИД 33072003 .
- ^ Jump up to: а б Олив М., Моерман Ф., Фернандес-Касси Х, Альтерматт Ф., Кон Т. (2022). «Удаление вирусов, передающихся через воду, с помощью Tetrahymenapyriformis является вирусспецифичным и совпадает с изменениями скорости плавания простейших» . Экологические науки и технологии . 56 (7): 4062–4070. Бибкод : 2022EnST...56.4062O . doi : 10.1021/acs.est.1c05518 . ПМЦ 8988290 . ПМИД 35258957 .
- ^ Jump up to: а б с д ДеЛонг Дж.П., Ван Эттен Дж.Л., Аль-Амели З., Агаркова И.В., Дуниган Д.Д. (03.01.2023). «Потребление вирусов возвращает энергию в пищевые цепи» . Труды Национальной академии наук . 120 (1): e2215000120. Бибкод : 2023PNAS..12015000D . дои : 10.1073/pnas.2215000120 . ISSN 0027-8424 . ПМЦ 9910503 . ПМИД 36574690 . S2CID 255219850 .
- ^ Jump up to: а б Хадас Э., Мари Д., Шпигель М., Илан М. (2006). «Похищение вирусов губками — это новый путь поступления питательных веществ в пищевые сети коралловых рифов» . Лимнология и океанография . 51 (3): 1548–1550. Бибкод : 2006LimOc..51.1548H . дои : 10.4319/lo.2006.51.3.1548 .
- ^ Jump up to: а б Уэлш Дж.Э., Стенхейс П., де Мораес К.Р., ван дер Меер Дж., Тилтгес Д.В., Брюссаард К.П. (2020). «Похищение морских вирусов организмами, не являющимися хозяевами» . Научные отчеты . 10 (1): 5221. Стартовый код : 2020NatSR..10.5221W . дои : 10.1038/s41598-020-61691-y . ПМК 7089979 . ПМИД 32251308 .
- ^ Фурман Дж. А. (июнь 1999 г.). «Морские вирусы и их биогеохимические и экологические эффекты». Природа . 399 (6736): 541–548. Бибкод : 1999Natur.399..541F . дои : 10.1038/21119 . ПМИД 10376593 . S2CID 1260399 .
- ^ Саттл, Калифорния (2007). «Морские вирусы — основные игроки глобальной экосистемы» . Обзоры природы Микробиология . 5 (10): 801–812. дои : 10.1038/nrmicro1750 . ISSN 1740-1534 . ПМИД 17853907 . S2CID 4658457 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Майерс К.М., Кулиш С., Бассо Дж.Т., Салтведт М.Р., Бьюкен А., Сандаа Р.А. (февраль 2023 г.). Прасад В.Р. (ред.). «Выпас морских вирусов и его биогеохимические последствия» . мБио . 14 (1): e0192121. дои : 10.1128/mbio.01921-21 . ПМЦ 9973340 . ПМИД 36715508 .
- ^ Jump up to: а б с д и Ховер Л.Ф., Эффлер Т.К., Бьюкен А., Вильгельм С.В., Вайц Дж.С. (июль 2014 г.). «Элементный состав вирусных частиц: значение для морских биогеохимических циклов». Обзоры природы. Микробиология . 12 (7): 519–528. дои : 10.1038/nrmicro3289 . ПМИД 24931044 . S2CID 17421986 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Денг Л., Краусс С., Файхтмайер Дж., Хофманн Р., Арндт Х., Гриблер К. (август 2014 г.). «Выпас гетеротрофных жгутиковых на вирусах обусловлен пищевым поведением». Отчеты по экологической микробиологии . 6 (4): 325–330. Бибкод : 2014EnvMR...6..325D . дои : 10.1111/1758-2229.12119 . ПМИД 24992530 .
- ^ Вильгельм С.В., Саттл, Калифорния (1999). «Вирусы и циклы питательных веществ в море» . Бионаука . 49 (10): 781–788. дои : 10.2307/1313569 . ISSN 1525-3244 . JSTOR 1313569 .
- ^ Jump up to: а б с «Кормление государств» . depts.washington.edu . Проверено 29 марта 2023 г.
- ^ Jump up to: а б с Олив М., Моерман Ф., Фернандес-Касси Х, Альтерматт Ф., Кон Т. (апрель 2022 г.). «Удаление вирусов, передающихся через воду, с помощью Tetrahymenapyriformis является вирусспецифичным и совпадает с изменениями скорости плавания простейших» . Экологические науки и технологии . 56 (7): 4062–4070. Бибкод : 2022EnST...56.4062O . doi : 10.1021/acs.est.1c05518 . ПМЦ 8988290 . ПМИД 35258957 .
- ^ Jump up to: а б с д Вермонт А.И., Мартинес Мартинес Дж., Уоллер Дж.Д., Гилг И.С., Ливитт А.Х., Флоге С.А. и др. (сентябрь 2016 г.). «Вирусная инфекция Emiliania huxleyi отпугивает веслоногих ракообразных Acartia тонна» . Журнал исследований планктона . 38 (5): 1194–1205. дои : 10.1093/plankt/fbw064 .
- ^ Jump up to: а б с д и ж г час я дж к Эванс С., Уилсон WH (2008). «Преимущественный выпас пристани Oxyrris на зараженной вирусом Emiliania huxleyi» . Лимнология и океанография . 53 (5): 2035–2040. Бибкод : 2008LimOc..53.2035E . дои : 10.4319/lo.2008.53.5.2035 . S2CID 16440441 .
- ^ Jump up to: а б с д и ж г час я дж к л м Фрада М.Дж., Шац Д., Фарстей В., Оссолински Дж.Э., Сабанай Х., Бен-Дор С. и др. (ноябрь 2014 г.). «Зоопланктон может служить переносчиком вирусов, поражающих цветение водорослей в океане» . Современная биология . 24 (21): 2592–2597. дои : 10.1016/j.cub.2014.09.031 . ПМИД 25438947 . S2CID 2682946 .
- ^ Хеврони Дж., Флорес-Урибе Х., Бежа О., Философ А. (ноябрь 2020 г.). «Сезонные и суточные закономерности численности и активности вирусов в Красном море» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (47): 29738–29747. Бибкод : 2020PNAS..11729738H . дои : 10.1073/pnas.2010783117 . ПМЦ 7703586 . ПМИД 33172994 .
- ^ Jump up to: а б Белчер А., Манно С., Уорд П., Хенсон С.А., Сандерс Р., Тарлинг Г.А. (24 марта 2017 г.). «Перенос фекальных гранул копепод через мезо- и батипелагические слои Южного океана весной» . Биогеонауки . 14 (6): 1511–1525. Бибкод : 2017BGeo...14.1511B . дои : 10.5194/bg-14-1511-2017 . ISSN 1726-4170 .
- ^ Jump up to: а б с д и ж г час я Уэлш Дж.Э., Стинхейс П., де Мораес К.Р., ван дер Меер Дж., Тилтгес Д.В., Бруссаард К.П. (март 2020 г.). «Похищение морских вирусов организмами, не являющимися хозяевами» . Научные отчеты . 10 (1): 5221. Бибкод : 2020NatSR..10.5221W . дои : 10.1038/s41598-020-61691-y . ПМК 7089979 . ПМИД 32251308 .
Дальнейшее чтение
[ редактировать ]- Криадо Ма (12 января 2023 г.). «Живоядные организмы могут съесть до миллиона вирусов в день» . ЭЛЬ-ПАИС английский .
- Шраге С. (3 января 2023 г.). «Поедание вирусов может способствовать росту и размножению микроорганизмов» . Небраска сегодня .