Непрерывная эволюция с помощью фага
Непрерывная эволюция с помощью фагов ( PACE ) — это основанный на фагах метод автоматизированной направленной эволюции белков. Он основан на связи желаемой активности целевого белка с приспособленностью инфекционного бактериофага, несущего соответствующий ген белка. Белки с более высокой желаемой активностью, следовательно, придают большую инфекционность своему фагу-носителю. Более инфекционные фаги размножаются более эффективно, отбирая выгодные мутации. Генетические вариации генерируются с помощью склонных к ошибкам полимераз на фаговых векторах , и со временем в белке накапливаются полезные мутации. Этот метод примечателен тем, что выполняет сотни раундов отбора с минимальным вмешательством человека.
Принцип
[ редактировать ]Центральным компонентом ПАСЕ является судно фиксированного объема, известное как «лагуна». Лагуна содержит векторы бактериофага M13, несущие интересующий ген (известный как селективная плазмида или SP), а также клетки-хозяева E. coli , которые позволяют фагу реплицироваться. Лагуну постоянно разбавляют путем добавления и слива жидкой среды, содержащей E. coli клетки . Скорость потока жидкости устанавливают такой, чтобы скорость разведения была быстрее, чем скорость размножения E.coli , но медленнее, чем скорость размножения фага. свежий запас клеток E. coli , но сохранить фаг можно только за счет достаточно быстрой репликации. Следовательно, в лагуне постоянно присутствует [ 1 ]
Репликация фага требует заражения E. coli , которая в случае фага M13 зависит от белка III (pIII). [ 2 ] При использовании PACE в фаговых векторах отсутствует ген, продуцирующий pIII. Вместо этого продукция pIII связана с активностью интересующего белка посредством механизма, который варьируется в зависимости от случая использования, часто включает дополнительную плазмиду, содержащую экспрессирующий pIII ген III (gIII), известный как вспомогательная плазмида, или AP. Примечательно, что продукция инфекционных фаговых чешуек с продукцией pIII. [ 3 ] Следовательно, чем выше активность белка, тем выше скорость продукции pIII и тем более инфекционный фаг генерируется для этого конкретного гена.
Используя склонные к ошибкам полимеразы (закодированные в мутагенезной плазмиде или MP), генетические вариации вводятся в белковую генную часть фаговых векторов. Из-за селективного давления, оказываемого постоянным осушением лагуны, в лагуне могут оставаться только фаги, которые могут реплицироваться достаточно быстро, поэтому со временем в фаге, реплицирующемся в лагуне, накапливаются полезные мутации. Таким образом, циклы эволюции выполняются непрерывно, позволяя пройти сотням раундов при минимальном вмешательстве человека. [ 1 ]

Приложения
[ редактировать ]Специфичность промотора полимеразы
[ редактировать ]В первой статье, посвященной этой методике, РНК-полимеразы Т7 были разработаны для распознавания различных промоторов , таких как промоторы Т3 или SP6. [ 4 ] Это было сделано путем назначения целевого промотора единственным промотором gIII. [ 5 ] Следовательно, мутантные полимеразы с большей специфичностью к желаемому промотору вызывают большую продукцию pIII. В результате были получены полимеразы с активностью на ~3-4 порядка большей для целевого промотора, чем исходный промотор Т3. [ 4 ] Хотя эта оригинальная система PACE осуществляла только положительный отбор, был разработан вариант, допускающий также отрицательный отбор. Это достигается путем связывания нежелательной активности с выработкой нефункционального pIII, что уменьшает количество образующегося инфекционного фага. [ 6 ]
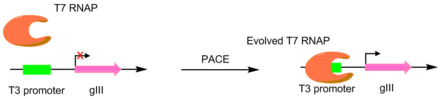
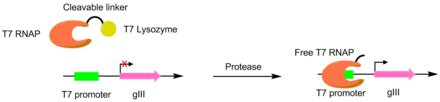
Специфичность субстрата протеазы
[ редактировать ]Протеазы были разработаны для разрезания различных пептидов с помощью PACE. В этих системах желаемый сайт разреза протеазы используется для связывания РНК-полимеразы Т7 и лизоцима Т7 . Лизоцим Т7 предотвращает транскрипцию gIII полимеразой Т7. Когда пептидный линкер расщепляется, полимераза Т7 активируется, обеспечивая транскрипцию гена pIII. Этот метод был использован для создания протеазы TEV с существенно другим пептидным субстратом. [ 6 ] [ 7 ]
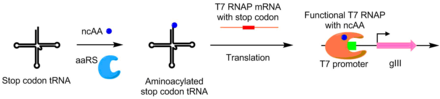
Ортогональные аминоацил-тРНК-синтетазы
[ редактировать ]С помощью PACE аминоацил-тРНК-синтетазы были разработаны для неканонических аминокислот (aaRS) и . Активность aaRS связана с производством pIII за счет добавления стоп-кодона TAG в середине gIII. Синтетазы, которые аминоацилируют тРНК-супрессор кодона TAG, предотвращают активность стоп-кодона , позволяя производить функциональный pIII. С помощью этой системы были разработаны aaRS, в которых используются неканонические аминокислоты п -нитрофенилаланин, йодфенилаланин и Вос-лизин. [ 8 ]
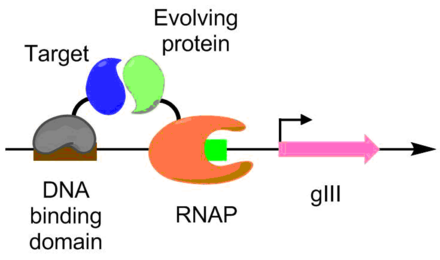
Белко-белковые взаимодействия
[ редактировать ]Белко-белковые взаимодействия также были разработаны с использованием PACE. Согласно этой схеме целевой белок слит с ДНК-связывающим белком, который связывается с целевой последовательностью, расположенной выше промотора gIII. Белок, подвергающийся эволюции, слит с РНК-полимеразой. Чем лучше белок-белковое взаимодействие, тем больше происходит транскрипции pIII, что позволяет эволюцию белок-белкового взаимодействия в условиях PACE. [ 6 ] Этот метод был использован для создания Bacillus thuringiensis вариантов эндотоксина , способных преодолевать устойчивость насекомых к токсинам. [ 6 ] [ 9 ]
Базовые редакторы
[ редактировать ]PACE использовался для развития APOBEC1 для большей растворимой экспрессии. APOBEC1 представляет собой цитидиндезаминазу , которая нашла применение в редакторах оснований для катализа редактирования одного нуклеотида C-->T. [ 10 ] В E. coli APOBEC1 обычно выпадает из раствора в нерастворимую фракцию. [ 11 ] Чтобы развить APOBEC1 для лучшей растворимой экспрессии, N-конец полимеразы T7 был слит с APOBEC1, а оставшаяся часть полимеразы экспрессировалась отдельно. Полимераза Т7 может функционировать только тогда, когда N-концевая часть может связываться с остальной частью полимеразы. Поскольку APOBEC1 должен быть правильно свернут, чтобы N-концевая часть была правильно обнажена, активность полимеразы T7 коррелирует со сворачиванием APOBEC1. Таким образом, транскрипция и продукция pIII связана с растворимой экспрессией APOBEC1 через полимеразу T7. Используя этот подход, экспрессия растворимого APOBEC1 увеличивалась в 4 раза без изменения функции. [ 7 ] [ 9 ]
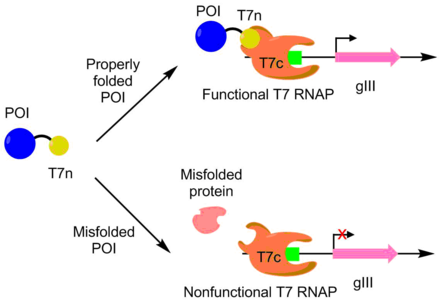
PACE также использовался для создания более каталитически активной дезоксиаденозиндезаминазы. Дезоксиаденозиндезаминаза используется в редакторах оснований для редактирования одного нуклеотида A-->T. Это было сделано путем помещения аденозинсодержащих стоп-кодонов в ген полимеразы Т7. Если базовый редактор способен исправить ошибку, образуется функциональная полимераза Т7, позволяющая производить pIII. Используя эту систему, они разработали дезоксиаденозиндезаминазу с активностью, в 590 раз превышающей активность дикого типа. [ 12 ]

Ссылки
[ редактировать ]- ^ Jump up to: а б Эсвелт, К.; Карлсон, Дж.; Лю, Д.Р. (2011). «Система непрерывной направленной эволюции биомолекул» . Природа . 472 (7344): 499–503. Бибкод : 2011Natur.472..499E . дои : 10.1038/nature09929 . ПМК 3084352 . ПМИД 21478873 .
- ^ Рихманн, Л.; Холлигер, П. (1997). «С-концевой домен TolA является корецептором инфекции нитчатым фагом E. coli » . Клетка . 90 (2): 351–360. дои : 10.1016/s0092-8674(00)80342-6 . ПМИД 9244308 .
- ^ Раконьяк, Дж.; Модель, П. (1998). «Роль pIII в сборке нитчатых фагов». Дж. Мол. Биол . 282 (1): 25–41. дои : 10.1006/jmbi.1998.2006 . ПМИД 9733639 .
- ^ Jump up to: а б Лейн, доктор медицины; Силиг, Б. (2014). «Достижения в области направленной эволюции белков» . Курс. Мнение. хим. Биол . 22 : 129–136. дои : 10.1016/j.cbpa.2014.09.013 . ПМЦ 4253873 . ПМИД 25309990 .
- ^ Лемир, С.; Йель, К.М.; Лу, ТК (2018). «Применение фагов в синтетической биологии» . Анну. Преподобный Вирол . 5 (1): 453–476. doi : 10.1146/annurev-virology-092917-043544 . ПМЦ 6953747 . ПМИД 30001182 .
- ^ Jump up to: а б с д Бредель, АК; Исалан, М.; Харамилло, А. (2018). «Инженерия биомолекул путем эволюции, направленной бактериофагами» . Курс. Мнение. Биотехнология . 51 : 32–38. дои : 10.1016/j.copbio.2017.11.004 . hdl : 10261/184372 . ПМИД 29175708 .
- ^ Jump up to: а б Ким, JY; Йоу, ХВ; Ли, PG; Ли, СГ; Со, Дж. Х.; Ким, Б.Г. (2019). « Эволюция белков in vivo , стратегия белковой инженерии следующего поколения: от случайного подхода к целенаправленному подходу». Биотехнология. Биопрок. Э. 24 : 85–94. дои : 10.1007/s12257-018-0394-2 . S2CID 91687131 .
- ^ Варгас-Родригес, О.; Севостьянова А.; Зёлль, Д.; Црнкович, А. (2018). «Модернизация аминоацил-тРНК-синтетаз для расширения генетического кода» . Курс. Мнение. хим. Биол . 46С : 115–122. дои : 10.1016/j.cbpa.2018.07.011 . ПМК 7083171 . ПМИД 30056281 .
- ^ Jump up to: а б Саймон, Эй Джей; д'Ольсниц, С.; Эллингтон, AD (2018). «Синтетическая эволюция». Нат. Биотехнология . 37 (7): 730–743. дои : 10.1038/s41587-019-0157-4 . ПМИД 31209374 . S2CID 189927244 .
- ^ Гауделли, Нью-Мексико; Комор, AC; Рис, штат Ха; Пакер, Миссисипи; Бадран, Ах; Брайсон, ДИ; Лю, ДР (2017). «Программируемое редактирование оснований A·T на G·C в геномной ДНК без расщепления ДНК» . Природа . 551 (7681): 464–471. дои : 10.1038/nature24644 . ПМК 5726555 . ПМИД 29160308 .
- ^ Ван, Т.; Бадран, Ах; Хуанг, ТП; Лю, ДР (2018). «Непрерывная направленная эволюция белков с улучшенной растворимой экспрессией» . Нат. хим. Биол . 14 (10): 972–980. дои : 10.1038/s41589-018-0121-5 . ПМК 6143403 . ПМИД 30127387 .
- ^ Рихтер, МФ; Чжао, Коннектикут; Итон, Э.; Лапинайте А.; Ньюби, Джорджия; Турони, Б.В.; Уилсон, К.; Коблан, ЛВ; Цзэн, Дж.; Бауэр, Делавэр; Дудна, Дж.А.; Лю, ДР (2020). «Фаговая эволюция редактора адениновых оснований с улучшенной совместимостью и активностью Cas-домена» . Нат. Биотехнология . 38 (7): 883–891. дои : 10.1038/s41587-020-0453-z . ПМЦ 7357821 . ПМИД 32433547 .