Бочковая кора
Эта статья может быть слишком технической для понимания большинства читателей . ( январь 2016 г. ) |
| Бочковая кора | |
|---|---|
| Идентификаторы | |
| НейроЛекс ID | nlx_81 |
| Анатомические термины нейроанатомии | |

Бочковая кора — это область соматосенсорной коры , которая встречается у некоторых видов грызунов и видов по крайней мере двух других отрядов. [1] и содержит поле ствола. «Бочки» бочкообразного поля представляют собой области внутри коркового слоя IV , которые при окрашивании становятся заметно темнее, чтобы выявить присутствие цитохром-с-оксидазы , и отделены друг от друга более светлыми областями, называемыми перегородками. Эти темные области являются основной мишенью для соматосенсорных сигналов от таламуса , и каждый ствол соответствует определенной области тела. Благодаря этой отличительной клеточной структуре, организации и функциональному значению бочкообразная кора является полезным инструментом для понимания кортикальной обработки и играет важную роль в нейробиологии. [2] Большая часть того, что известно о кортикоталамической обработке, получена в результате изучения бочкообразной коры, и исследователи интенсивно изучали бочкообразную кору как модель неокортикального столба .
Наиболее отличительной чертой бочки являются стволы с усами. Эти структуры были впервые обнаружены Вулси и Ван дер Лоосом в 1970 году. [3] Окрашивание в бочонках усов более отчетливое, чем в других участках соматосенсорной коры. Признавая, что расположение вибрисс ( усов) похоже на мистициальную подушечку (область, из которой растут усы) некоторых млекопитающих, они выдвинули гипотезу, что стволы являются «кортикальными коррелятами мистициальных вибрисс» и что «один ствол представляет собой одна вибрисса». В то время как небольшие участки бочкообразной коры, не являющиеся усами, соответствуют большим и иногда перекрывающимся областям тела, каждый гораздо более крупный бочонок усов соответствует одному усу. В результате бочонки усов находятся в центре внимания большинства исследований бочковой коры, а термин «бочкообразная кора» часто используется в первую очередь для обозначения бочонков усов. Следовательно, большая часть этой статьи посвящена коре головного мозга усов грызунов.
Организация бочковых полей
[ редактировать ]
Поле ствола, как и многие области коры головного мозга, организовано в виде топографической карты . В случае бочкового поля карта соматотопическая – основана на расположении частей тела. Области, соответствующие носу и рту, расположены на карте более рострально и латерально, передние конечности, задние конечности и туловище - более медиально, причем передние конечности рострально от задних конечностей, а бочковые подполя усов - заднемедиальное подполе ствола, что соответствует основному лицевые усы (мистициальные вибриссы) и переднелатеральное бочкообразное подполе, соответствующее меньшим усам лица, — каудальные и латеральные. Хотя усы составляют относительно небольшую часть тела животного, они доминируют на соматотопической карте. [4] [5]
Стволы крупных лицевых бакенбардов
[ редактировать ]Бочонки, соответствующие основным усам лица (мистациальным вибриссам), содержатся в заднемедиальном подполе бочонков (PMBSF). Бочки здесь самые крупные и эллиптические по форме и имеют поразительную топографическую организацию, идентичную таковой усов; они организованы в 5 рядов по 4–7 больших усов, идущих почти параллельно переносице. [6] Организация мистических вибрисс и соответствующих им стволов настолько согласована, что существует соглашение об именах, позволяющее идентифицировать каждый уик у крыс и мышей. Ряды обозначаются от A до E сверху вниз, а столбцы усов в каждом ряду нумеруются сзади вперед. Первые четыре строки также имеют дополнительный ус позади столбца 1, который обозначается строчной или греческой буквой (α, β, γ или δ). Эти четыре уса также называют стрэддлерами.
Анатомия и соединение стволов
[ редактировать ]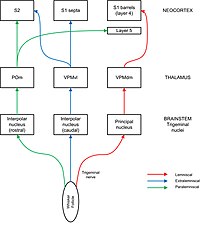
Бочонки бочкообразной коры получили свое название потому, что по плотности клетки напоминали бочонки , то есть собраны в цилиндрические формы, суженные вверху и внизу. Центр ствола обозначается полостью, а промежутки между стволами - перегородками (единственное число: перегородка). [6]
Сенсорная информация передается от фолликулов усов к бочкообразной коре через ядра тройничного нерва и таламус. Бочкообразные деления можно увидеть в некоторых, но не во всех частях ядер тройничного нерва (где они называются бочонками) и таламуса (где они называются барроидами). Тройничный нерв несет афферентные волокна от фолликулов в ствол мозга, где они соединяются с нейронами четырех различных ядер тройничного нерва: главного, межполярного, орального и каудального. Отростки ядер тройничного нерва к таламусу разделяются на пути, называемые лемнискальными, экстралемнискальными и паралемнискальными. В лемнискальном пути аксоны главного ядра тройничного нерва пересекают срединную линию и проецируются в «баррелоиды» таламуса, особенно в дорсомедиальной части вентрозаднего медиального ядра (VPMdm). Нейроны в VPMdm проецируются в основном в стволы слоя 4 первичной соматосенсорной коры (S1). По экстралемнискальному пути нейроны интерполярного ядра проецируются в вентролатеральный отдел вентрозаднего медиального ядра (VPMvl). Нейроны в VPMvl проецируются на перегородки между стволами и на вторичная соматосенсорная кора (S2). Паралемнискальный путь проходит от межполярного ядра тройничного нерва через заднее ядро (POm) таламуса к S2 и к диффузным мишеням в бочкообразной коре, особенно в слое 5. Каждый путь также имеет вторичные проекции на другие слои бочкообразной коры и другие области коры, включая моторная кора. [7] Считается, что эти разные пути передают разные модальности сенсорной информации от усов. [2] [8]
Нейрофизиология ствола усов
[ редактировать ]Кора головного мозга «усов» содержит различные типы нейронов, которые получают входные данные из ряда источников, которые сами получают и обрабатывают массив различных типов информации. В результате нейроны коры ствола усов реагируют на входные сигналы, связанные с усами, но способом, специфичным для типа и местоположения нейронов. Это может проявляться по-разному. Простейший вопрос заключается в том, реагирует ли кортикальный нейрон только на отклонение одного уса или на отклонение многих усов. Нейроны в стволах слоя 4 имеют тенденцию сильно или исключительно реагировать на один уик, в то время как нейроны в других слоях настроены менее сильно и могут реагировать на несколько усов. Нейроны, которые реагируют на отклонение нескольких усов, обычно имеют первичный уик, на который они реагируют больше всего. Разница в величине реакции между отклонением первичного и вторичного усов также может варьироваться в зависимости от нейрона. Стимуляция нескольких усов может вызвать реакцию, равную сумме ответов, если бы каждый уик стимулировался независимо, или она может быть разной. Некоторые нейроны демонстрируют более сильные реакции, когда несколько нейронов стимулируются последовательно, и последовательность может быть специфичной для направления. [9]
Нейроны могут реагировать не только на комбинации, при которых были стимулированы усы, но и на определенные типы стимуляции усов. Самая простая реакция, наблюдаемая в нейронах бочкообразного слоя IV коры, непосредственно кодирует смещение усов. То есть нейрон внутри данного ствола сработает, когда ус, который представляет собой ствол, будет перемещаться со скоростью, примерно пропорциональной угловому смещению нейрона. Эти нейроны также проявляют направленную чувствительность; определенные нейроны срабатывают только тогда, когда ус перемещается в определенном направлении. [10] [11] Запускающие нейроны, основанные на отклонении, могут поддерживать свою реакцию на протяжении всего отклонения уса. Другие нейроны реагируют на первоначальное отклонение, но затем быстро возвращаются к прежнему уровню активности. Большая часть этой активности также модулируется поведением животного — крысы и мыши активно двигают усами, исследуя окружающую среду, а реакция нейрона на тот или иной стимул может варьироваться в зависимости от того, что делает животное.
Пластичность, зависящая от опыта
[ редактировать ]Поскольку бочкообразная кора имеет хорошо организованную структуру, которая четко связана с подушечкой усов, она широко использовалась в качестве инструмента для изучения сенсорной обработки и развития, а также феномена пластичности, зависящей от опыта - изменений в активности, связности и структура нейронных цепей в ответ на опыт. Нейроны бочкообразной коры обладают свойством синаптической пластичности , которое позволяет им изменять вибриссы, на которые они реагируют, в зависимости от истории тактильного опыта грызуна. [12] Зависимая от опыта пластичность обычно изучается в бочкообразной коре путем частичного лишения ее сенсорной информации либо путем повреждения элементов афферентного пути (например, тройничного нерва), либо путем удаления, выщипывания или обрезки некоторых лицевых усов. На анатомическую структуру стволов влияет только повреждение элементов проводящего пути, но безобидные формы депривации могут вызывать быстрые изменения в кортикальной карте во взрослом возрасте без каких-либо соответствующих изменений в структурах стволов. [13] Из-за разных эффектов кажется, что эти две парадигмы действуют по разным механизмам.
Некоторые формы пластичности бочкообразной коры переживают критический период . Выщипывание усов у новорожденных крыс вызывает длительное расширение представленности сохраненного уса в слое 4. [14] Однако пластичность слоя 4 быстро снижается, если сенсорная депривация начинается после 4-го дня жизни (P4), тогда как репрезентации в слое 2/3 остаются высоко пластичными и во взрослом возрасте. [15] [16]
Два корковых процесса протекают параллельно друг другу, когда бочкообразная кора лишается сенсорной информации от некоторых усов, чтобы обеспечить репрезентативную пластичность. В депривированной коре нейрональные реакции на сохраненные усы усиливаются, а реакции на лишенные усы ослабляются. Эти два процесса имеют разные временные ходы: ослабление депривированной реакции предшествует усилению сэкономленной реакции, а это означает, что в их основе лежат разные механизмы. Эти два эффекта в совокупности приводят к расширению коркового представительства сохраненных усов в репрезентацию соседних лишенных усов. [15] [17]
Вполне вероятно, что в создании зависимого от опыта опыта задействовано несколько различных механизмов.пластичность в протоколе лишения усов (адаптировано из Фельдмана и Брехта, 2005 г.) [17] ):
- Почти сразу потеря подачи питания на колонну с лишенным стволом приводит к потере тормозной стрельбы в этой колонне. Это демаскирует горизонтальные возбуждающие соединения от соседних резервных колонн. [18] Это не объясняет более длительных пластических изменений, поскольку демаскировка исчезла бы немедленно, если бы утраченный ресурс был восстановлен (например, если позволить усам вырасти заново).
- LTP и LTD Похоже, что здесь также задействованы процессы, подобные . Об этом можно сделать вывод, используя трансгенных мышей, у которых наблюдаются изменения в экспрессии ферментов, связанных с LTP и LTD, например, кальмодулин-зависимой протеинкиназы II (CaMKII) или белка, связывающего элемент ответа циклического АМФ (CREB). У этих мышей пластичность нарушена. [19] [20] Важным фактором может быть время пика, а не частота. Ассоциативная LTP была продемонстрирована в синапсах слоев 4 и 2/3, когда нейрон слоя 4 срабатывает на 0–15 мс раньше нейрона слоя 2/3, и LTD наблюдается, когда этот временной порядок изменен на обратный. [21] Такие механизмы могут действовать быстро и вызывать пластические изменения в течение нескольких часов или дней.
- Было продемонстрировано, что сенсорная депривация вызывает изменения в синаптической динамике, такой как ВПСП амплитуда и частота . Конечным эффектом этих изменений является увеличение доли синаптической информации, которую нейроны слоя 2/3 в депривированных стволах получают от сохраненных стволов. [22] Эти наблюдения позволяют предположить, что в зависимости от опыта пластичности участвуют и другие, более специфические механизмы, помимо LTP/LTD.
- Интуитивно кажется вероятным, что структурные изменения на уровне аксонов, ветвей дендритов и отростков дендритов лежат в основе некоторых долговременных пластических изменений в коре. Сообщалось об изменениях в структуре аксонов в пластичности после поражений. [23] и совсем недавно в ходе исследований по обрезке усов. [24] Дендритное ветвление важно во время пренатального и неонатального развития, участвует в пластичности, индуцированной повреждениями, но не участвует в пластичности, зависящей от опыта. [25] Двухфотонная микроскопия in vivo показывает, что дендритные шипы в бочкообразной коре мыши очень динамичны и подвержены непрерывному обновлению и могут быть связаны с образованием или удалением синапсов. [25] Вполне вероятно, что оборот позвоночника необходим, но недостаточен для обеспечения пластичности, зависящей от опыта, и другие механизмы, такие как ремоделирование аксонов, также необходимы для объяснения таких особенностей, как экономия на предыдущем опыте. [24]
Пластичность и ремоделирование бочкообразной коры также изучались в контексте черепно-мозговой травмы . [26] где было показано, что обогащение стимулов окружающей средой вызывает пластичность / восстановление [27] а модели временного кодирования были изменены с помощью механизмов пластичности и восстановления. [28]
Примечания
[ редактировать ]- ^ Вулси и др., 1975.
- ^ Jump up to: а б Фокс, 2008 г.
- ^ Вулси и Ван дер Лоос, 1970.
- ^ Гувер и др., 2003 г.
- ^ Энрикес-Баррето и др., 2012 г.
- ^ Jump up to: а б Вулси и Ван дер Лоос, 1970 г.
- ^ Босман и др., 2011.
- ^ Даймонд и др., 2008 г.
- ^ Босман и др., 2011.
- ^ Свадлоу, 1989.
- ^ Свадлоу Х.А. (1991). «Эфферентные нейроны и предполагаемые интернейроны во второй соматосенсорной коре бодрствующего кролика: рецептивные поля и свойства аксонов». J Нейрофизиология . 66 (4): 1392–1409. дои : 10.1152/jn.1991.66.4.1392 . ПМИД 1761989 .
- ^ Хардингем Н., Глазевски С., Пахотин П., Мизуно К., Чепмен П.Ф., Гизе К.П., Фокс К. Неокортикальная долговременная потенциация и зависящая от опыта синаптическая пластичность требуют аутофосфорилирования альфа-кальций/кальмодулин-зависимой протеинкиназы II. Дж. Нейроски. 2003, 1 июня; 23(11): 4428-36.
- ^ Фокс К. (2002). «Анатомические пути и молекулярные механизмы пластичности в бочкообразной коре». Нейронаука . 111 (4): 799–814. дои : 10.1016/s0306-4522(02)00027-1 . ПМИД 12031405 . S2CID 39423181 .
- ^ Фокс К. (1992). «Критический период зависящей от опыта синаптической пластичности в коре головного мозга крысы» . Дж. Нейроски . 12 (5): 1826–1838. doi : 10.1523/JNEUROSCI.12-05-01826.1992 . ПМК 6575898 . ПМИД 1578273 .
- ^ Jump up to: а б Глазевски С., Фокс К. (1996). «Временной ход зависимой от опыта синаптической потенциации и депрессии в бочкообразной коре крыс-подростков». J Нейрофизиология . 75 (4): 1714–1729. дои : 10.1152/jn.1996.75.4.1714 . ПМИД 8727408 .
- ^ Стерн Э.А., Маравалл М., Свобода К. (2001). «Быстрое развитие и пластичность карт слоя 2/3 в коре головного мозга крысы in vivo» . Нейрон . 31 (2): 305–315. дои : 10.1016/s0896-6273(01)00360-9 . ПМИД 11502260 . S2CID 2819415 .
- ^ Jump up to: а б Фельдман Д.Е., Брехт М. (2005). «Карта пластичности соматосенсорной коры». Наука . 310 (5749): 810–815. дои : 10.1126/science.1115807 . ПМИД 16272113 . S2CID 2892382 .
- ^ Келли МК, Карвелл GE, Коджер Дж. М., Саймонс DJ (1999). «Потеря чувствительности в результате удаления усов вызывает немедленное растормаживание соматосенсорной коры ведущих крыс» . Дж. Нейроски . 19 (20): 9117–25. doi : 10.1523/JNEUROSCI.19-20-09117.1999 . ПМК 6782760 . ПМИД 10516329 .
- ^ Глазевски С., Чен С.М., Сильва А., Фокс К. (1996). «Требование к альфа-CaMKII для пластичности коры головного мозга, зависящей от опыта». Наука . 272 (5260): 421–423. Бибкод : 1996Sci...272..421G . дои : 10.1126/science.272.5260.421 . ПМИД 8602534 . S2CID 84433995 .
- ^ Глазевски С., Барт А.Л., Уоллес Х., Маккенна М., Сильва А., Фокс К. (1999). «Нарушение пластичности, зависящей от опыта, в бочкообразной коре мышей, у которых отсутствуют альфа- и дельта-изоформы CREB» . Кора головного мозга . 9 (3): 249–256. дои : 10.1093/cercor/9.3.249 . ПМИД 10355905 .
- ^ Фельдман Д.Е. (2000). «LTP и LTD на основе синхронизации при вертикальных входах в пирамидные клетки слоя II/III в бочкообразной коре головного мозга крысы» . Нейрон . 27 (1): 45–56. дои : 10.1016/s0896-6273(00)00008-8 . ПМИД 10939330 . S2CID 17650728 .
- ^ Финнерти Г.Т., Робертс Л.С., Коннорс Б.В. (1999). «Сенсорный опыт изменяет краткосрочную динамику неокортикальных синапсов». Природа . 400 (6742): 367–371. Бибкод : 1999Natur.400..367F . дои : 10.1038/22553 . ПМИД 10432115 . S2CID 4413560 .
- ^ Чкловский Д.Б., Мел Б.В., Свобода К. (2004). «Корковая перепроводка и хранение информации». Природа . 431 (7010): 782–788. Бибкод : 2004Natur.431..782C . дои : 10.1038/nature03012 . ПМИД 15483599 . S2CID 4430167 .
- ^ Jump up to: а б Cheetham CE, Hammond MS, MacFarlane R, Finnerty GT (2008)Измененный сенсорный опыт вызывает целенаправленную перестройку локальных возбуждающих связей в зрелом неокортексе. J Neurosci (в печати).
- ^ Jump up to: а б Трахтенберг Дж.Т., Чен Б.Е., Нотт Г.В., Фэн Г., Санес Дж.Р., Велкер Э., Свобода К. (2002). «Долгосрочная визуализация in vivo зависимой от опыта синаптической пластичности коры головного мозга взрослых». Природа . 420 (6917): 788–794. Бибкод : 2002Natur.420..788T . дои : 10.1038/nature01273 . ПМИД 12490942 . S2CID 4341820 .
- ^ Каррон, Симона Ф.; Алвис, Дасуни С.; Раджан, Рамеш (2016). «Каррон С.Ф., Алвис Д.С., Раджан Р. Черепно-мозговая травма и изменения функциональности нейронов в сенсорной коре головного мозга. Front Syst Neurosci. 2016;10 (июнь):47. doi:10.3389/fnsys.2016.00047» . Границы системной нейронауки . 10 : 47. дои : 10.3389/fnsys.2016.00047 . ПМЦ 4889613 . ПМИД 27313514 .
- ^ Алвис, Д.С.; Ян, Е.Б.; Джонстон, В.; Каррон, С.; Хеллевелл, С.; Морганти-Коссманн, MC; Раджан, Р. (2016). «Алвис Д.С., Ян Э.Б., Джонстон В. и др. Обогащение окружающей среды ослабляет черепно-мозговую травму: индуцированная гипервозбудимость нейронов в супрагранулярных слоях сенсорной коры. J Neurotrauma. 2016;33(11). doi:10.1089/neu.2014.3774» . Журнал нейротравмы . 33 (11): 1084–1101. дои : 10.1089/neu.2014.3774 . ПМИД 26715144 .
- ^ ТОМАС ФРЭНСИС БЕРНС (2019). Бернс (2019) Паттерны активности временных нейронов в бочкообразной коре головного мозга в зависимости от простых и сложных стимулов и последствий черепно-мозговой травмы. Университет Монаша. Диссертация. 10.26180/5б7166ад13е47 (диссертация). Университет Монаша. дои : 10.26180/5b7166ad13e47 .
Ссылки
[ редактировать ]- Босман Л.В., Хаувелинг А.Р., Оуэнс CB, Танке Н., Шевчук О.Т., Рахмати Н., Теуниссен В.Х., Джу К., Гонг В., Куккук С.К., Де Зеув К.И. (2011). «Анатомические пути, участвующие в создании и восприятии ритмических движений усов» . Передний. Интегр. Нейроски . 5 : 53. дои : 10.3389/fnint.2011.00053 . ПМК 3207327 . ПМИД 22065951 .
- Даймонд М.Э., фон Хеймендал М., Кнутсен П.М., Кляйнфельд Д., Ахиссар Э. (2008). « Где» и «что» в сенсомоторной системе усов». Nat Rev Neurosci . 9 (8): 601–612. дои : 10.1038/nrn2411 . ПМИД 18641667 . S2CID 6450408 .
- Энрикес-Баррето, л; Палацетти С; Бреннаман Л.Х.; Манесс П.Ф.; Файрен А (2012). «Молекула адгезии нервных клеток, NCAM, регулирует поиск пути таламокортикальных аксонов и организацию коркового соматосенсорного представительства у мышей» . Границы молекулярной нейронауки . 5 : 76. doi : 10.3389/fnmol.2012.00076 . ПМЦ 3378950 . ПМИД 22723769 .
- Фокс, К. (2008). Баррель Кортекс . Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-0-521-85217-3 .
- Гувер, Дж. Э., Хоффер, З. С., Аллоуэй, К. Д. (2003). «Проекции первичной соматосенсорной коры на неостриатум: роль соматотопической непрерывности в кортикостриарной конвергенции». Журнал нейрофизиологии . 89 (3): 1576–1587. дои : 10.1152/jn.01009.2002 . ПМИД 12611938 . S2CID 3002038 .
- Свадлоу Х.А. (1989). «Эфферентные нейроны и предполагаемые интернейроны в коре вибрисс S-1 бодрствующего кролика: рецептивные поля и свойства аксонов». J Нейрофизиология . 62 (1): 288–308. дои : 10.1152/jn.1989.62.1.288 . ПМИД 2754479 .
- Вулси, штат Техас; Ван дер Лоос, Х (1970). «Структурная организация слоя IV в соматосенсорной области (SI) коры головного мозга мыши: описание коркового поля, состоящего из дискретных цитоархитектонических единиц». Исследования мозга . 17 (2): 205–242. дои : 10.1016/0006-8993(70)90079-x . ПМИД 4904874 .
- Вулси Т.А., Велкер С., Шварц Р.Х. (1975). «Сравнительные анатомические исследования коры головного мозга SmL с особым упором на возникновение «бочек» в IV слое» . Журнал сравнительной неврологии . 164 (1): 79–94. дои : 10.1002/cne.901640107 . ПМИД 809494 . S2CID 12374398 . Архивировано из оригинала (pdf) 23 декабря 2012 г.
Внешние ссылки
[ редактировать ]Исследовательские группы, работающие над бочкообразной корой:
- Ахиссар Лаборатория, Израиль
- Лаборатория Барта, Питтсбург, США
- Лаборатория Бруно, Оксфорд, Великобритания
- Лаборатория тактильного восприятия и обучения (Diamond Lab), Триест, Италия
- Лаборатория Фельдмана, Беркли, США
- Лаборатория Финнерти, Центр исследований нейродегенерации MRC, Лондон
- Группа бочковой коры (Fox Lab), Кардифф
- Группа сенсорных систем и нейронной инженерии (Лаборатория Хартмана), Чикаго, США
- Лаборатория Хельмхена, Цюрих, Швейцария
- Hires Lab, Лос-Анджелес, США
- Лаборатория Кляйнфельда, Сан-Диего, США
- Лаборатория Маравалл, Сассекс, Великобритания
- Лаборатория Мура, Провиденс, США
- Оберлендерская лаборатория, Бонн, Германия
- Лаборатория О'Коннора, Балтимор, США
- Лаборатория сенсорной обработки (Petersen Lab), Лозанна, Швейцария
- Schwarz Lab, Тюбинген, Германия
- Barrel Group (Staiger Lab), Геттинген, Германия
- Стэнли Лаборатория, Атланта, США
- Лаборатория Свобода, Сиэтл, США
Книги о бочковой коре