Цикл Рэндла
Цикл Рэндла , также известный как цикл глюкозы и жирных кислот , представляет собой метаболический процесс, включающий конкуренцию глюкозы и жирных кислот за субстраты . [ 1 ] Предполагается, что он играет роль в объяснении диабета 2 типа и резистентности к инсулину . [ 2 ] [ 3 ]
Он был назван в честь Филипа Рэндла , описавшего его в 1963 году. [ 4 ]
Цикл
[ редактировать ]Цикл Рэндла представляет собой биохимический механизм, включающий конкуренцию между глюкозой и жирными кислотами за их окисление и поглощение мышечной и жировой тканью . Цикл контролирует выбор топлива и адаптирует поставку и потребность субстрата в нормальных тканях. Этот цикл добавляет тонкую настройку, опосредованную питательными веществами, поверх более грубого гормонального контроля топливного метаболизма. Эта адаптация к доступности питательных веществ касается взаимодействия между жировой тканью и мышцами. Гормоны, контролирующие липолиз жировой ткани , влияют на концентрацию циркулирующих жирных кислот, которые, в свою очередь, контролируют выделение топлива в мышцах. Механизмы, участвующие в цикле Рэндла, включают аллостерический контроль, обратимое фосфорилирование и экспрессию ключевых ферментов. [ 5 ] Энергетический баланс блюд, состоящих из разного состава макронутриентов, идентичен, но балансы глюкозы и жира, которые вносят вклад в общий энергетический баланс, изменяются в зависимости от состава еды. [ 6 ]

Глюкоза сохраняется и перенаправляется
[ редактировать ]Состояние натощак
[ редактировать ]При голодании активация липолиза делает жирные кислоты предпочтительным источником топлива для дыхания. В печени β-окисление жирных кислот удовлетворяет местные энергетические потребности и может привести к кетогенезу (созданию кетоновых тел из жирных кислот). Кетоновые тела затем используются для удовлетворения потребностей других тканей, кроме печени. Это ингибирование окисления глюкозы на уровне пируватдегидрогеназы сохраняет пируват и лактат , которые являются глюконеогена . предшественниками [ 5 ]
Федеральный резерв
[ редактировать ]Цикл глюкозы и жирных кислот также наблюдается в сытом состоянии после приема пищи с высоким содержанием жиров или во время физических упражнений. Это когда концентрация жирных кислот или кетоновых тел в плазме увеличивается. Неокисленная глюкоза затем перенаправляется в гликоген . Такое перенаправление на гликоген объясняет быстрый ресинтез мышечного гликогена после тренировки, а также повышенное содержание гликогена в мышцах, наблюдаемое при голодании или диабете. Этот механизм восполняет промежуточные продукты цикла лимонной кислоты . [ 5 ]
Ингибирование гликолитического пути
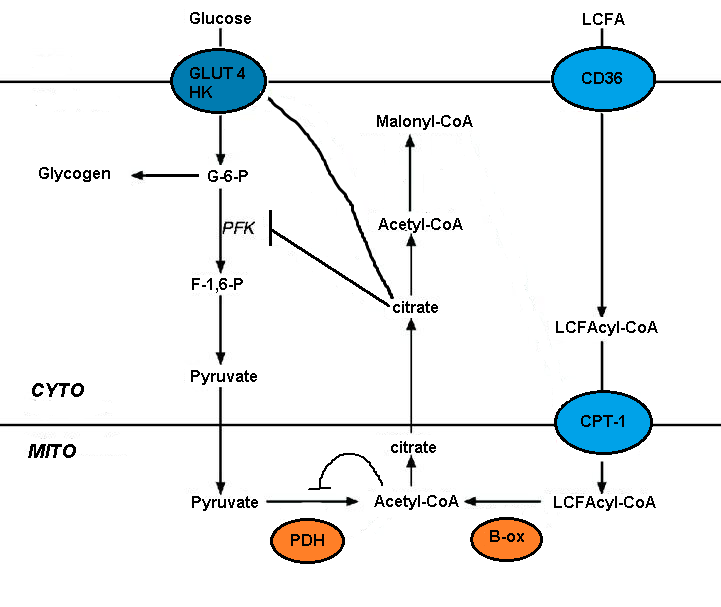
[ редактировать ]Нарушение метаболизма глюкозы за счет окисления жирных кислот опосредовано кратковременным торможением ряда гликолитических процессов. Степень ингибирования увеличивается по гликолитическому пути, будучи наиболее выраженной на уровне пируватдегидрогеназы и менее выраженной на уровне поглощения глюкозы и 6-фосфофрукто-1-киназы ( PFK-1 ). [ 5 ] Эта последовательность возникает из-за того, что начальным событием, вызванным окислением жирных кислот, является увеличение митохондриальных соотношений [ацетил-КоА]/[КоА] и [НАДН]/[НАД+]. Оба они служат для ингибирования активности пируватдегидрогеназы. [ 7 ] Было высказано предположение, что эти изменения приводят к накоплению цитозольного цитрата, который, в свою очередь, ингибирует PFK-1, с последующим увеличением уровня глюкозо-6-фосфата, который в конечном итоге ингибирует гексокиназу. [ 5 ]
Гемодинамический стресс
[ редактировать ]Гемодинамический стресс подавляет ингибирование жирными кислотами метаболизма глюкозы. В это время наблюдается уменьшение предложения субстрата и увеличение потребности в субстрате. Это приводит к активации АМФ-активируемой протеинкиназы (АМФК), поскольку концентрация АМФ во внутриклеточной жидкости повышается, а концентрация АТФ снижается. Активация AMPK, вызванная стрессом, обеспечивает немедленную метаболическую адаптацию и защищает сердце от ишемического стресса. [ 5 ] [ 8 ] [ 9 ]
Ингибирование окисления жирных кислот малонил-КоА
[ редактировать ]Малонил-КоА сигнализирует об утилизации глюкозы и контролирует поступление и окисление длинноцепочечных жирных кислот (ДЦЖК) в митохондриях . Циркуляция глюкозы в печени стимулирует ее усвоение. -карбоксилазы может быть преобразован в малонил-КоА При окислении глюкозы образуется цитрат, который под действием ацетил-КоА . Малонил-КоА ингибирует карнитинпальмитоилтрансферазу (CPT), которая контролирует проникновение и окисление LCFA. Получаемый из глюкозы малонил-КоА предотвращает окисление жирных кислот и способствует этерификации жирных кислот. [ 4 ] [ 5 ]

Цитозольные события, контролирующие окисление жирных кислот
[ редактировать ]Концентрация малонил-КоА
[ редактировать ]Концентрация малонил-КоА зависит от баланса между ацетил-КоА-карбоксилазой (АСС) и малонил-КоА-декарбоксилазой (MCD). Сообщается, что AMP-активируемая протеинкиназа (AMPK) фосфорилирует и инактивирует АСС печени. Это, в свою очередь, снижает концентрацию малонил-КоА, что стимулирует окисление жирных кислот и кетогенез глюкагоном в печени. AMPK фосфорилирует и инактивирует АСС в печени и других тканях. [ 4 ] [ 5 ]
Интеграция AMPK и АСС в цикле глюкоза-жирные кислоты
[ редактировать ]Ингибирование окисления жирных кислот требует активности АСС. И AMPK, и MCD неактивны, а поглощение глюкозы стимулируется. LCFA затем перенаправляются на этерификацию. [ 10 ] Эти состояния существуют в тканях, богатых кислородом, в которых AMPK неактивен, а глюкоза инактивирует AMPK (исследования проводились в скелетных мышцах). [ 11 ]
Ингибирование MCD подавляет окисление жирных кислот и стимулирует окисление глюкозы. В исследовании на мышах с дефицитом MCD не было обнаружено различий в окислении жирных кислот и глюкозы в сердце в аэробных условиях. Предполагается, что сверхэкспрессия используемых жирных кислот компенсирует недостаток MCD. [ 12 ]
Поглощение жирных кислот
[ редактировать ]Поглощение длинноцепочечных жирных кислот опосредовано несколькими переносчиками, включая FAT (транслоказу жирных кислот)/CD36. Удаление CD36 спасает липотоксическую кардиомиопатию. FAT/CD36 можно контролировать с помощью инсулина и AMPK. Усиление транспорта в сочетании с образованием производных КоА и результирующей активацией AMPK должно обеспечивать эффективное поглощение и метаболизм жирных кислот. [ 5 ]
Митохондриальные события, контролирующие выбор топлива
[ редактировать ]Жирные кислоты преимущественно окисляются из-за инактивации ПДГ путем окисления жирных кислот, ингибирующего окисление глюкозы. Это предполагает, что митохондриальный метаболизм может контролировать выбор топлива. Клеточное дыхание стимулируется жирными кислотами, и это связано с увеличением соотношения НАДН и НАД+ в митохондриях, что позволяет предположить, что обеспечение энергией опережает потребление энергии. Переключение с окисления глюкозы на окисление жирных кислот приводит к тому, что большая часть электронов транспортируется к комплексу 2, а не к комплексу 1 дыхательной цепи. Эта разница приводит к менее эффективному окислительному фосфорилированию. Окисляя жирные кислоты, митохондрии усиливают дыхание, одновременно увеличивая выработку АФК. [ 5 ]
Жирные кислоты и инсулин
[ редактировать ]Жирные кислоты могут действовать непосредственно на β-клетки поджелудочной железы, регулируя стимулируемую глюкозой секрецию инсулина. Этот эффект является двухфазным. Первоначально жирные кислоты потенцируют действие глюкозы. После длительного воздействия высоких концентраций жирных кислот это меняется на торможение. [ 13 ] Рэндл предположил, что термин «синдром жирных кислот» будет уместно применять к биохимическому синдрому, возникающему в результате высокой концентрации жирных кислот и связи с нарушениями углеводного обмена, включая голодание, диабет и синдром Кушинга . [ 4 ]
Ссылки
[ редактировать ]- ^ Бевилаква С., Буцциголи Г., Бонадонна Р. и др. (1990). «Работа цикла Рэндла у больных ИНСД». Диабет . 39 (3): 383–9. дои : 10.2337/диабет.39.3.383 . ПМИД 2307295 .
- ^ Шульдинер А.Р., МакЛенитан Дж.К. (2004). «Гены и патофизиология диабета 2 типа: больше, чем просто цикл Рэндла снова и снова» . Дж. Клин. Инвестируйте . 114 (10): 1414–7. дои : 10.1172/JCI23586 . ПМК 525752 . ПМИД 15545992 .
- ^ Деларю Дж., Маньян С. (2007). «Свободные жирные кислоты и резистентность к инсулину». Текущее мнение о клиническом питании и метаболической помощи . 10 (2): 142–8. дои : 10.1097/MCO.0b013e328042ba90 . ПМИД 17285001 . S2CID 9620797 .
- ^ Jump up to: а б с д Рэндл П.Дж., Гарланд П.Б., Хейлз К.Н., Ньюсхолм Э.А. (1963). «Цикл жирных кислот глюкозы. Его роль в чувствительности к инсулину и метаболических нарушениях при сахарном диабете». Ланцет . 1 (7285): 785–9. дои : 10.1016/S0140-6736(63)91500-9 . ПМИД 13990765 .
- ^ Jump up to: а б с д и ж г час я дж Хюэ Л., Тэгтмейер Х (2009). «Возвращение к циклу Рэндла: новая голова вместо старой шляпы» . Американский журнал физиологии. Эндокринология и обмен веществ . 297 (3): E578–E591. дои : 10.1152/ajpendo.00093.2009 . ПМЦ 2739696 . ПМИД 19531645 .
- ^ Фрейн К.Н. (2003). «Цикл глюкозы-жирных кислот: физиологический взгляд». Биохим Соц Транс . 31 (Часть 6): 1115–9. дои : 10.1042/bst0311115 . ПМИД 14641007 .
- ^ Боукер-Кинли М.М., Дэвис В.И., Ву П., Харрис Р.А., Попов К.М. (1998). «Доказательства существования тканеспецифической регуляции комплекса пируватдегидрогеназы млекопитающих» . Биохим. Дж . 329 : 191–6. дои : 10.1042/bj3290191 . ПМК 1219031 . ПМИД 9405293 .
- ^ Кудо Н., Гиллеспи Дж.Г., Кунг Л., Виттерс Л.А., Шульц Р., Кланачан А.С., Лопащук Г.Д. (1996). «Характеристика активности 5'АМФ-активируемой протеинкиназы в сердце и ее роли в ингибировании ацетил-КоА-карбоксилазы во время реперфузии после ишемии» . Биохим Биофиз Акта . 1301 (1–2): 67–75. дои : 10.1016/0005-2760(96)00013-6 . ПМИД 8652652 .
- ^ Гудвин Г.В., Тэгтмейер Х (2000). «Улучшение энергетического гомеостаза сердца в метаболическом состоянии при физической нагрузке». Американский журнал физиологии. Физиология сердца и кровообращения . 279 (4): H1490–H1501. дои : 10.1152/ajpheart.2000.279.4.H1490 . ПМИД 11009433 .
- ^ Кларк Х., Карлинг Д., Саггерсон Д. (2004). «Ковалентная активация сердечной AMP-активируемой протеинкиназы в ответ на физиологические концентрации длинноцепочечных жирных кислот» . Eur J Biochem . 271 (11): 2215–24. дои : 10.1111/j.1432-1033.2004.04151.x . ПМИД 15153111 .
- ^ Итани СИ; Саха АК; Куровский Т.Г.; Гроб HR; Торнхейм К; Рудерман Н.Б. (2003). «Глюкоза автоматически регулирует свое поглощение при участии скелетных мышц АМФ-активируемой протеинкиназы» . Диабет . 52 (7): 1635–1640. дои : 10.2337/диабет.52.7.1635 . ПМИД 12829626 .
- ^ Дайк Дж.Р.Б., Хопкинс Т.А., Боннет С., Микелакис Э.Д., Янг М.Э., Ватанабэ М., Кавасе Ю., Джишаге К., Лопащук Г.Д. (2006). «Отсутствие малонил-коэнзима А-декарбоксилазы у мышей увеличивает окисление сердечной глюкозы и защищает сердце от ишемического повреждения» . Журнал Американской кардиологической ассоциации . 114 (16): 1721–1728. doi : 10.1161/CIRCULATIONAHA.106.642009 . ПМИД 17030679 .
- ^ Гриль V, Квигстад Э (2000). «Жирные кислоты и секреция инсулина» . Британский журнал питания . 83 : S79–S84. дои : 10.1017/S0007114500000994 . ПМИД 10889796 .