Бочковая кора
Эта статья может быть слишком технической для большинства читателей, чтобы понять . ( Январь 2016 ) |
| Бочковая кора | |
|---|---|
| Идентификаторы | |
| Neurolex Id | NLX_81 |
| Анатомические термины нейроанатомии | |

Кора ствола - это область соматосенсорной коры , которая идентифицируется у некоторых видов грызунов и видов, по крайней мере, двух других заказов [ 1 ] и содержит поле ствола. «Бочки» поля ствола представляют собой области внутри кортикального слоя IV , которые заметно темнее, когда окрашивают, чтобы выявить присутствие цитохрома С оксидазы и отделяются друг от друга более легкими областями, называемыми септа. Эти темные области являются основной целью для соматосенсорных входов из таламуса , и каждая ствола соответствует области тела. Благодаря этой отличительной клеточной структуре, организации и функциональной значимости кора ствола является полезным инструментом для понимания обработки коры и сыграла важную роль в нейробиологии. [ 2 ] Большая часть того, что известно о обработке кортикоталамической, связана с изучением коры ствола, и исследователи интенсивно изучали кору ствола как модель неокортикальной колонки .
Наиболее отличительным аспектом бочкового поля являются бочки Whisker. Эти структуры были впервые обнаружены Вулси и Ван дер Лоосом в 1970 году. [ 3 ] Окрашивание в бочках Whisker более отчетливо, чем в других областях соматосенсорной коры. Признавая, что массив был похож на массив вибриссов (усов) на мистапиальной подушечке (области, откуда отдаваемых усы) некоторых млекопитающих, они предположили, что бочки были «корковыми коррелятами мистациальных вибриссов» и что «один бочки представляет одна вибрасса ». Принимая во внимание, что небольшие участки с бочкой, не являющиеся бешеной, соответствуют большим, а иногда и перекрывающимся областям тела, каждый гораздо более крупный бочонок усы соответствует одному усы. В результате бочки Whisker находятся в центре большинства исследований коры ствола, а «кора бочки» часто используется для обращения в основном к бочкам Whisker. Следовательно, большая часть этой статьи фокусируется на коре бочки грызунов.
Организация бочковых полей
[ редактировать ]
Поле ствола, как и многие области коры, организовано в топографическую карту . В случае поля ствола карта соматотопная - на основе расположения частей тела. Области, соответствующие носу и рту, являются более ростральными и боковыми на карте, передняя передняя конечность, задница и туловище являются более медиальными, с передней передней частью передней части задних конечностей и подполя бочки Whisker - подставка для постеромедиальной бочки, который соответствует майору усы для лица (мистапиальные вибриссы) и подполе с бочкой, который соответствует меньшим Усы лица - каудальные и боковые. Хотя усы составляют относительно небольшую часть животного, они доминируют над соматотопной картой. [ 4 ] [ 5 ]
Бочки основных усов лица
[ редактировать ]Стволы, которые соответствуют основным усам лица (мистапиальные вибриссы), содержатся в подполе с бочками (PMBSF). Бочки здесь являются крупнейшей и наиболее эллиптической формой и имеют поразительную топографическую организацию, которая идентична организации усов; Они организованы в 5 рядов из 4-7 больших усов, которые находятся рядом с параллелью с мостом. [ 6 ] Организация мистапиальных вибриссов и соответствующих бочек настолько последовательна, что существует соглашение об именах для идентификации каждого присада у крыс и мышей. Ряды обозначены от A до E сверху вниз, а столбцы усов в каждом ряду пронумерованы сзади на спереди. Первые четыре ряда также имеют дополнительный усы за столбцом 1, который обозначен буквой нижней части или греческой буквы (α, β, γ или Δ). Эти четыре усы также называются Straddlers.
Анатомия и связь бочек
[ редактировать ]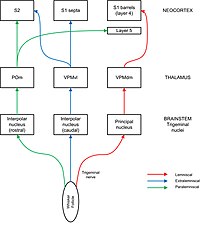
Стволы коры ствола были названы, потому что плотность клеток напоминала бочки , то есть они собираются в цилиндрические формы, которые сужаются вверху и снизу. Центр ствола обозначается впадиной, а пространства между бочками - это перегородки (единственное число: перегородка) [ 6 ]
Сенсорная информация течет от фолликулов Whisker в кору ствола через ядра тройничного нерва и таламус. Ствол, подобные подразделениям, можно увидеть в некоторых, но не во всех частях ядер тройничных ядер (где они называются баррелями) и таламуса (где они называются графролоидами). Тригемиальный нерв несет афферентные волокна из фолликулов в ствол мозга, где они соединяются с нейронами в четырех различных ядрах нервного нерва: основной, межполярной, пероральный и хвостовой. Прогнозы от ядер тройничных ядер до таламуса разделены на пути, обозначенные Lemniscal, Extralemniscal и Paralemniscal. В Lemniscal пути аксоны из основного тригеминального ядра пересекаются над средней линией и проецируют на «Баррелоиды» в таламусе, в частности в дорсомедиальном сечении вентропостериального медиального ядра (VPMDM). Нейроны в VPMDM Проект в основном на бочки в слое 4 первичной соматосенсорной коры (S1). В экстралеменсистском пути нейроны межполярного ядра проецируют в вентролатеральном участке вентропостериального медиального ядра (VPMVL). Нейроны в VPMVL Проект на септу между бочками и на Вторичная соматосенсорная кора (S2). Паралеменскальный путь проходит от межполярного тригеминального ядра через заднее ядро (POM) таламуса до S2 и к диффузным мишеням в коре ствола, в частности, слой 5. Каждый путь также имеет вторичные выступления на другие слои в коре ствола и другие области коры, включая вторичные проекции в других слоях в коре ствола и других областей, включая вторичные выступления на другие слои в коре и других областях. Моторная кора. [ 7 ] Считается, что эти разные пути передают различные методы сенсорной информации от Whisker. [ 2 ] [ 8 ]
Нейрофизиология ствола Whisker
[ редактировать ]Кора ствола Whisker содержит различные типы нейронов, которые получают входные данные из диапазона источников, которые сами получают, и обрабатывают множество различных типов информации. В результате нейроны коры ствола Whisker реагируют на ввод, связанный с усы, но в каком-то смысле, характерные для типа и местоположения нейронов. Это может проявляться по -разному. Самый простой способ - реагировать ли кортикальный нейрон только отклонение одного уса или на отклонение многих усов. Нейроны в бочках 4 -го слоя имеют тенденцию сильно или исключительно реагируют на один усы, в то время как нейроны в других слоях менее сильно настроены и могут реагировать на несколько усов. Нейроны, которые реагируют на отклонение нескольких усов, обычно имеют первичный усы, на что они реагируют больше всего. Разница в величине ответа между отклонениями первичного уса и вторичными усами также может варьироваться между нейронами. Стимуляция нескольких усов может вызывать ответ, равный сумме ответов, если каждый усы стимулировался независимо, или он может отличаться. Некоторые нейроны показывают большие ответы, когда несколько нейронов стимулируются в последовательности, а последовательность может быть специфичной для направления. [ 9 ]
Наряду с комбинациями которых стимулировались усы, нейроны также могут реагировать на конкретные типы стимуляции усов. Самый простой ответ, наблюдаемый в нейронах в коре ствола слоя IV, непосредственно кодирует для смещения Whisker. То есть, что нейрон в данной стволе будет стрелять, когда усы, который представляет собой ствол, перемещается со скоростью, которая примерно пропорциональна угловому смещению нейрона. Эти нейроны также демонстрируют чувствительность направления; Некоторые нейроны будут стрелять только тогда, когда усы перемещаются в определенном направлении. [ 10 ] [ 11 ] Основанные на отклонениях нейроны могут выдержать свой ответ на протяжении всего отклонения усов. Другие нейроны реагируют на первоначальное отклонение, но затем быстро возвращаются к своему предыдущему уровню активности. Большая часть этой деятельности также модулируется поведением животных - крыс и мышей, активно перемещающих свои усы, чтобы исследовать их окружающую среду, и реакция нейрона на конкретный стимул может варьироваться в зависимости от того, что делает животное.
В зависимости от опыта пластичность
[ редактировать ]Поскольку кора ствола имеет хорошо организованную структуру, которая четко связана с подушкой Whisker, она широко использовалась в качестве инструмента для изучения сенсорной обработки и развития, а также явление зависимой от опыта пластичности-изменения в активности, связности и подключении и Структура нейронных схем в ответ на опыт. Нейроны в коре ствола демонстрируют свойство синаптической пластичности , которое позволяет им изменять вибриссы, на которые они реагируют в зависимости от истории тактильного опыта грызунов. [ 12 ] В зависимости от опыта пластичность обычно изучается в коре ствола путем частично лишения его сенсорного ввода, либо путем поразительных элементов афферентного пути (например, тройного нерва), либо путем абляции, выщипывания или обрезки некоторых лицевых усов. Анатомическая структура стволов влияет только поразительные элементы пути, но безобидные формы депривации могут вызвать быстрые изменения в коры в зрелом возрасте, без каких -либо соответствующих изменений в структурах бочек. [ 13 ] Из -за их различных эффектов кажется, что эти две парадигмы работают по разным механизмам.
Некоторые формы пластичности в коре ствола отображают критический период . Усы из неонатальных крыс вызывает долгосрочное расширение представления пробежавшего усы в слое 4. [ 14 ] Тем не менее, слой 4 пластичность быстро уменьшается, если сенсорная депривация начинается после 4 -го дня жизни (P4), тогда как представления в слое 2/3 остаются очень пластическими в взрослом возрасте. [ 15 ] [ 16 ]
Два процесса коры проходят рядом друг с другом, когда кора ствола лишена сенсорного ввода от некоторых усов для создания репрезентативной пластичности. В обездоленной коре нейрональные реакции на щастные усы увеличены, а реакции на лишенные усы ослаблены. Эти два процесса имеют разные временные курсы, с ослаблением лишенного ответа, предшествующего укреплению спасенного ответа, подразумевая, что они имеют различные базовые механизмы. Эти два эффекта объединяются для получения расширения коркового представления щавших усов в представление соседних лишенных усов. [ 15 ] [ 17 ]
Вполне вероятно, что несколько различных механизмов участвуют в создании зависимых от опыта Пластичность в протоколе депривации усов (адаптирована из Feldman and Brecht, 2005 [ 17 ] ):
- Почти сразу же потеря ввода в колонку обрученного ствола приводит к потере ингибирующей стрельбы в этом столбце. Это разоблачает горизонтальные возбуждающие соединения из соседних слоев. [ 18 ] Это не объясняет более длительные пластиковые изменения, так как разоблачение немедленно исчезнет, если будет восстановлен лишенной вход (например, путем позволения усам отрастать).
- LTP - и LTD , подобные процессам, также, кажется, вовлечены. Это может быть выведено с использованием трансгенных мышей, где существуют изменения в экспрессии ферментов, связанных с LTP и LTD, например, кальмодулин-зависимой протеинкиназой II (CAMKII) или белка связывания элемента ответа циклического AMP (CREB). У этих мышей пластичность скомпрометирована [ 19 ] [ 20 ] Время шипа, а не частота может быть важным фактором. Ассоциативный LTP был продемонстрирован на слое 4 до синапсов слоя 2/3, когда нейрон слой 4 запускает 0-15 мс до нейрона уровня 2/3, а LTD наблюдается, когда этот порядок времени обращается вспять. [ 21 ] Такие механизмы могут действовать быстро, чтобы произвести пластические изменения в течение нескольких часов или дней.
- Было продемонстрировано, что сенсорная депривация вызывает изменения в синаптической динамике, таких как EPSP амплитуда и частота . Чистый эффект этих изменений заключается в увеличении доли синаптического входа, который нейроны 2/3 в обездоленных стволах получают из счетов. [ 22 ] Эти наблюдения предполагают, что другие, более специфические механизмы, помимо LTP/LTD, играют в пластичности, зависящей от опыта.
- Кажется интуитивно вероятным, что структурные изменения на уровне аксонов, ветвей дендритов и дендритных шипов лежат в основе некоторых долгосрочных пластических изменений в коре. Изменения в структуре аксонов были зарегистрированы в пластичности после поражений [ 23 ] и совсем недавно в исследованиях с использованием обрезки Whisker. [ 24 ] Дендритное ветвление важно во время пренатального и неонатального развития, участвует в пластичности, вызванной поражениями, но не участвует в зависимой от опыта пластичности. [ 25 ] Двухфотонная микроскопия in vivo показывает, что дендритные шипы в коре мышиного бочка являются высоко динамическими и подвержены непрерывному обороту и могут быть связаны с образованием или удалением синапсов. [ 25 ] Вполне вероятно, что оборот позвоночника необходим, но не достаточна для создания зависимой от опыта пластичности, и другие механизмы, такие как ремоделирование аксонов, также необходимы для объяснения таких функций, как экономия от предыдущего опыта. [ 24 ]
Пластичность и ремоделирование коры ствола также изучались в контексте черепно -мозговой травмы , [ 26 ] Если было показано, что обогащение окружающей среды стимулов вызывает пластичность/восстановление [ 27 ] и паттерны временного кодирования были изменены с помощью пластичности и механизмов восстановления. [ 28 ]
Примечания
[ редактировать ]- ^ Woolsey et al., 1975
- ^ Jump up to: а беременный Фокс, 2008
- ^ Woolsey & van der Loos, 1970
- ^ Hoover et al., 2003
- ^ Enriquez-Barreto et al., 2012
- ^ Jump up to: а беременный Woolsey & van der Loos, 1970
- ^ Bosman et al., 2011
- ^ Diamond et al., 2008
- ^ Bosman et al., 2011
- ^ Swadlow, 1989
- ^ Swadlow HA (1991). «Эфферентные нейроны и подозреваемые интернейроны во второй соматосенсорной коре кролика бодрствования: восприимчивые поля и аксональные свойства». J Нейрофизиол . 66 (4): 1392–1409. doi : 10.1152/jn.1991.66.4.1392 . PMID 1761989 .
- ^ Hardingham N, Glazewski S, Pakhotin P, Mizuno K, Chapman PF, Giese KP, Fox K. Некортикальный долгосрочный потенцирование и зависимая от опыта синаптической пластичности требует альфа-кальциум/кальмодулинозависимого протеинкиназы II. J Neurosci. 2003 1 июня; 23 (11): 4428-36.
- ^ Fox K (2002). «Анатомические пути и молекулярные механизмы для пластичности в коре ствола». Нейробиология . 111 (4): 799–814. doi : 10.1016/s0306-4522 (02) 00027-1 . PMID 12031405 . S2CID 39423181 .
- ^ Fox K (1992). «Критический период для зависимой от опыта синаптической пластичности в коре крысы» . J Neurosci . 12 (5): 1826–1838. doi : 10.1523/jneurosci.12-05-01826.1992 . PMC 6575898 . PMID 1578273 .
- ^ Jump up to: а беременный Глазевский С., Фокс К. (1996). «Временный ход зависимого от опыта синаптического потенцирования и депрессии в коре ствола у подростковых крыс». J Нейрофизиол . 75 (4): 1714–1729. doi : 10.1152/jn.1996.75.4.1714 . PMID 8727408 .
- ^ Stern EA, Maravall M, Svoboda K (2001). «Быстрое развитие и пластичность карт 2/3 слоя в коре крысы in vivo» . Нейрон . 31 (2): 305–315. doi : 10.1016/s0896-6273 (01) 00360-9 . PMID 11502260 . S2CID 2819415 .
- ^ Jump up to: а беременный Фельдман Д.Е., Брехт М. (2005). «Карта пластичности в соматосенсорной коре». Наука . 310 (5749): 810–815. doi : 10.1126/science.1115807 . PMID 16272113 . S2CID 2892382 .
- ^ Келли М.К., Карвелл Г.Е., Коджер Дж. М., Саймонс DJ (1999). «Сенсорная потеря путем выбранного удаления Whisker вызывает немедленное рассеяние в соматосенсорной коре поведения крыс» . J. Neurosci . 19 (20): 9117–25. doi : 10.1523/jneurosci.19-20-09117.1999 . PMC 6782760 . PMID 10516329 .
- ^ Glazewski S, Chen CM, Silva A, Fox K (1996). «Требование для альфа-камки в зависимой от опыта пластичности коры бочки». Наука . 272 (5260): 421–423. Bibcode : 1996sci ... 272..421g . doi : 10.1126/science.272.5260.421 . PMID 8602534 . S2CID 84433995 .
- ^ Глазевский С., Барт А.Л., Уоллес Х, МакКенна М., Сильва А., Фокс К. (1999). «Нарушающая зависимая пластичность в бочковой коре мышей, в которых отсутствуют альфа и дельта изоформы CREB» . Мозговая кора . 9 (3): 249–256. doi : 10.1093/cercor/9.3.249 . PMID 10355905 .
- ^ Фельдман де (2000). «LTP и LTD на основе ГРМ на вертикальных входах в пирамидальные клетки слоя II/III в коре крысы» . Нейрон . 27 (1): 45–56. doi : 10.1016/s0896-6273 (00) 00008-8 . PMID 10939330 . S2CID 17650728 .
- ^ Финнерти Г.Т., Робертс Л.С., Коннорс Б.В. (1999). «Сенсорный опыт изменяет кратковременную динамику неокортикальных синапсов». Природа . 400 (6742): 367–371. Bibcode : 1999natur.400..367f . doi : 10.1038/22553 . PMID 10432115 . S2CID 4413560 .
- ^ Chklovskii DB, Mel BW, Svoboda K (2004). «Переводка кортикала и хранение информации». Природа . 431 (7010): 782–788. Bibcode : 2004natur.431..782c . doi : 10.1038/nature03012 . PMID 15483599 . S2CID 4430167 .
- ^ Jump up to: а беременный Cheetham CE, Hammond MS, Macfarlane R, Finnerty GT (2008) Измененный сенсорный опыт вызывает целенаправленное повторное поведение локальных возбуждающих соединений в зрелом неокортексе. J Neurosci (в прессе).
- ^ Jump up to: а беременный Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR, Welker E, Svoboda K (2002). «Долгосрочная визуализация in vivo зависимой от опыта синаптической пластичности у коры взрослых». Природа . 420 (6917): 788–794. Bibcode : 2002natur.420..788t . doi : 10.1038/nature01273 . PMID 12490942 . S2CID 4341820 .
- ^ Кэррон, Симона Ф.; Alwis, Dasuni S.; Раджан, Рамеш (2016). «Кэррон С.Ф., Алвис Д.С., Раджан Р. Травматическая травма головного мозга и изменения функциональности нейронов в сенсорной коре. Фронт Syst Syst Neurosci. 2016; 10 (июнь): 47. doi: 10.3389/fnsys.2016.00047» . Границы в системах нейробиологии . 10 : 47. doi : 10.3389/fnsys.2016.00047 . PMC 4889613 . PMID 27313514 .
- ^ Alwis, DS; Ян, Эб; Johnstone, V.; Кэррон, с.; Hellewell, S.; Morganti-Kossmann, MC; Раджан Р. (2016). «Alwis DS, Yan EB, Johnstone V и др. Обогащение окружающей среды ослабляет травматическую травму головного мозга: индуцированная гипергибируемость нейронов в супрагранулярных слоях сенсорной коры. J Neurotrauma. 2016; 33 (11). DOI: 10.1089/neu.2014.3774» . Журнал Neurotrauma . 33 (11): 1084–1101. doi : 10.1089/neu.2014.3774 . PMID 26715144 .
- ^ Томас Фрэнсис Бернс (2019). Бернс (2019) Паттерны временной нейрональной активности в коре ствола к простым и сложным стимулам и эффектам черепно -мозговой травмы. Университет Монаш. Тезис. 10.26180/5B7166AD13E47 (тезис). Университет Монаш. doi : 10.26180/5B7166AD13E47 .
Ссылки
[ редактировать ]- Bosman LW, Hold AR, Owens CB, Tanke N, Shefcho OT, Rahmati N, Tuniss WH, Jug W, Pickbook SK, Zeged CI (2011). «Анатумальные пути следует при создании и восприятии ритмических шепот движений» . Передний. Интеграция Нейроски . 5 : 53. DOI : 10 3389/fnt.2011 00053. PMC 3207327 . PMID 22065951 .
- Diamond Me, Von Heimendahl M, Knutsen PM, Kleinfeld D, Ahissar E (2008). « Где» и «что» в сенсомоторной системе Whisker ». Nat Rev Neurosci . 9 (8): 601–612. doi : 10.1038/nrn2411 . PMID 18641667 . S2CID 6450408 .
- Энрикес-Баррето, L; Palazzetti C; Бреннаман LH; Maness Pf; Fairén A (2012). «Молекула адгезии нейронных клеток, NCAM, регулирует исходное определение пута таламокортикального аксона и организацию коркового соматосенсорного представления у мышей» . Границы в молекулярной нейробиологии . 5 : 76. doi : 10.3389/fnmol.2012.00076 . PMC 3378950 . PMID 22723769 .
- Fox, K (2008). Бочковая кора . Кембридж, Великобритания: издательство Кембриджского университета. ISBN 978-0-521-85217-3 .
- Hoover, JE, Hoffer, ZS, Allingay, KD (2003). «Проекции от первичной соматосенсорной коры до неостриатума: роль соматотопной непрерывности в кортикостриатальной конвергенции». Журнал нейрофизиологии . 89 (3): 1576–1587. doi : 10.1152/jn.01009.2002 . PMID 12611938 . S2CID 3002038 .
- Swadlow HA (1989). «Эфферентные нейроны и подозреваемые интернейроны в коре вибриссы S-1 из бодрствующего кролика: восприимчивые поля и аксональные свойства». J Нейрофизиол . 62 (1): 288–308. doi : 10.1152/jn.1989.62.1.288 . PMID 2754479 .
- Вулси, та; Van Der Loos, H (1970). «Структурная организация уровня IV в соматосенсорной области (SI) коры головного мозга мыши: описание коркового поля, состоящего из дискретных цитоархитектонических единиц». Исследование мозга . 17 (2): 205–242. doi : 10.1016/0006-8993 (70) 90079-X . PMID 4904874 .
- Woolsey TA, Welker C, Schwartz RH (1975). «Сравнительные анатомические исследования коры лица SML с особой ссылкой на возникновение« бочек »в слое IV» . Журнал сравнительной неврологии . 164 (1): 79–94. doi : 10.1002/cne.901640107 . PMID 809494 . S2CID 12374398 . Архивировано из оригинала (PDF) 2012-12-23.
Внешние ссылки
[ редактировать ]Исследовательские группы, работающие на бочковой коре:
- Ахиссар Лаборатория, Израиль
- Barth Lab, Питтсбург, США
- Bruno Lab, Оксфорд, Великобритания
- Тактильное восприятие и учебная лаборатория (Diamond Lab), Триест, Италия
- Feldman Lab, Беркли, США
- Finnerty Lab, MRC Center of Neurodegeneration Research, Лондон
- Barrel Cortex Group (Fox Lab), Кардифф
- Sensory Systems & Neural Engineering Group (Hartmann Lab), Чикаго, США.
- Helmchen Lab, Цюрих, Швейцария
- Нанимайте лабораторию, Лос -Анджелес, США.
- Kleinfeld Lab, Сан -Диего, США
- Maravall Lab, Сассекс, Великобритания
- Moore Lab, Провиденс, США
- Оберландер -лаборатория, Бонн, Германия
- Lab O'Connor, Балтимор, США
- Лаборатория сенсорной обработки (Petersen Lab), Лозанна, Швейцария
- Schwarz Lab, Tübingen, Германия
- Barrel Group (Staiger Lab), Göttingen, Германия
- Stanley Lab, Атланта, США
- Svoboda Lab, Сиэтл, США
Книги на бочковой коре