Гистоноподобный белок нуклеоид-структурирования
| H-NS | |||
|---|---|---|---|
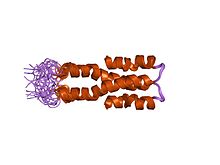 Структура раствора N-концевого домена (домен олигомеризации) бактериального нуклеоидного структурирующего белка, H-NS. | |||
| Идентификаторы | |||
| Символ | H-NS | ||
| Pfam | PF00816 | ||
| InterPro | IPR001801 | ||
| Кат - | [P0ACF8] | ||
| Краткое содержание | 1hns / scope / supfam | ||
| |||
Гистоноподобный белок-нуклеоиоид-структурирование (H-NS) является одним из двенадцати я нуклеоидных белков (SNAPS) [ 1 ] чья основная функция - организация генетического материала , включая регуляцию экспрессии генов посредством ксеногенного молчания. [ 2 ] H-NS характеризуется N-концевым доменом (NTD), состоящим из двух сайтов димеризации , области линкера, которая неструктурирована и C-концевой домен (CTD), который отвечает за связывание ДНК. [ 2 ] Хотя это маленький белок (15 кДа), [ 3 ] Он обеспечивает важное уплотнение нуклеоидов и регуляцию генов (в основном молчание) [ 2 ] и очень выражен, функционирует как димер или мультимер . [ 3 ] Изменение температуры вызывает диссоциирование H-NS от ДНК -дуплекса , что позволяет транскрипции РНК-полимеразой, а в специфических областях приводят к патогенным каскадам у энтеробактерий, таких как Escherichia coli и четырех видов шигеллы . [ 3 ]

Структура
[ редактировать ]H-NS имеет специфическую топологию, которая позволяет ей конденсировать бактериальную ДНК в сверхгелическую структуру, основанную на доказательствах рентгеновской кристаллографии . [ 2 ] Конденсированная супергелическая структура включает H-NS в репрессии генов, вызванных образованием олигомеров. Эти олигомеры образуются из-за димеризации двух сайтов в N-концевом домене H-NS. [ 2 ] Например, у бактериальных видов, таких как Salmonella typhimurium , NTD H-NS содержит сайты димеризации в спиралях Alpha 1, Alpha 2 и Alpha 3. Альфа-спирали 3 и 4 отвечают за создание сверхгелельной структуры взаимодействия H-NS-ДНК ДНК. от головы к главной ассоциации ( рис. 2 ). [ 2 ] [ 5 ] H-NS также содержит неструктурированную область линкера, также известную как Q-Linker. [ 2 ] С-концевой домен, также известный как ДНК-связывающий домен (DBD), демонстрирует высокую аффинность к областям в ДНК, которые богаты аденином и тимином и присутствуют в мотиве, подобном крючке в незначительной канавке. [ 2 ] Базовая укладка, присутствующая в этом в богатой области ДНК, позволяет незначительно расширить незначительную канавку, которая является преимущественным для связывания. [ 2 ] Общие DBD включают области AACTA и TACTA, которые могут появляться сотни раз по всему геному. [ 2 ] В рамках этих богатых регионов незначительная канавка имеет ширину 3,5 Å, [ 3 ] который является преимущественным для связывания H-NS. У E. coli наблюдалось, что H-NS реструктурирует геном в микродомены in vivo . [ 2 ] В то время как бактериальный геном разделен на четыре различных макродомены, включая ORI и TER (макродомен E. coli и Shigella spp. В котором кодируется H-NS), [ 3 ] Считается, что H-NS играет роль в формировании этих небольших 10 КБ микродоменов по всему геному. [ 2 ]

Функция
[ редактировать ]Основной функцией H-NS является влияние на топологию ДНК ( рис. 2 ). H-NS отвечает за образование нуклеофиламентов вдоль ДНК и мостов ДНК-ДНК. H-NS известен как пассивный Бриджер ДНК, что означает, что он связывает два отдаленных сегмента ДНК и остается неподвижным, образуя петлю. Это образование петли ДНК позволяет H-NS контролировать экспрессию генов. [ 2 ] Облегчение подавления с помощью H-NS может быть достигнуто путем связывания другого белка или изменений в топологии ДНК, которые могут возникнуть из-за изменений температуры и осмолярности , например. [ 6 ] CTD связывается с бактериальной ДНК таким образом, что ингибирует функцию РНК -полимеразы. Это общая особенность, наблюдаемая в горизонтально приобретенных генах. [ 7 ] Структурные исследования H-NS используют бактериальные виды, такие как E. coli и Shigella spp. Потому что С-концевой домен полностью сохраняется. [ 3 ]
Процесс формирования комплексов H-NS-ДНК начинается с связывания CTD с преференциальным сайтом в геноме. Это может быть результатом большого количества положительно заряженных аминокислотных остатков, расположенных в области линкера, что заставляет CTD искать сайт связывания с высокой аффинностью. [ 2 ] После того, как CTD связан с его преимущественной областью, этап TPA, NTD может олигомеризировать и образовывать жесткие нуклеофиламенты, которые, если существуют благоприятные условия, будут более свободно связываться друг с другом, образуя мостики ДНК. Эта форма моста известна как «пассивное мостовое» и может не позволить RNAP продолжить транскрипцию. [ 2 ] Эксперименты, используемые для поддержки этого метода связывания ДНК и молчания генов, поступают из-за атомной силовой микроскопии и одномолекулярных исследований in vitro . [ 2 ]
Все бактерии должны быть чувствительными к изменениям в их физической среде, чтобы выжить. Эти механизмы позволяют включать или выключать гены в зависимости от ее внеклеточной среды. [ 3 ] Многие исследователи считают, что H-NS способствует этим сенсорным функциям. Было обнаружено, что H-NS контролирует около 60% генов, регулируемых температурой, и может диссоциации от дуплекса ДНК при 37 ° C. [ 2 ] Эта конкретная чувствительность, наблюдаемая в H-NS, допускает патогенез и является основным направлением исследования. За пределами хозяина температура 32 ° C предотвращает диссоциацию H-NS из плазмиды вирулентности в Shigella spp. Чтобы сохранить энергию для энергетически дорогостоящей продукции белков, участвующих в патогенезе. [ 8 ] Наличие ионов магния (мг 2+ Было показано, что H-NS позволяет сформировать слегка открытое для полностью открытых конформационных изменений в структуре, которое в конечном итоге изменит взаимодействие между отрицательно заряженным NTD и положительно заряженным CTD. [ 2 ] Концентрации магния ниже 2 мМ позволяют образовать жесткие нуклеопротеиновые филаменты, а высокие концентрации способствуют образованию ДНК-мостов H-NS. [ 9 ] Заряды, наблюдаемые в NTD и CTD, могут объяснить, как H-NS остается чувствительным к изменениям температуры и осмолярности (рН ниже 7,4). [ 3 ] H-NS также может взаимодействовать с другими белками и влиять на их функцию, например, он может взаимодействовать с флигером жгутикового моторного белка для повышения его активности. [ 6 ]
Клиническое значение
[ редактировать ]
H-NS играет консервативную роль в патогенности грамотрицательных бактерий, включая Shigella spp., Escherichia coli , Salmonella spp. и многие другие. Он участвует в транскрипции гена Virf , вызывающего так называемое вирф, что приводит к бациллярной дизентерии , заболеванию, затрагивающему детей, в основном наблюдаемых в развивающихся странах. Эти два вида бактерий содержат плазмиду вирулентности , которая отвечает за инвазию клеток-хозяев и регулируется H-NS. [ 10 ] Интересно, что почти 70% открытых кадров считывания (ORF) специализированной плазмиды вирулентности в Shigella SPP. AT AT, позволяя долгосрочному регулированию этой плазмиды H-NS. [ 3 ]
Вышеупомянутые, исследования показывают, что чувствительность к температуре H-NS будет диссоциации от бактериальной ДНК при 37 ° C, запуская РНК-полимеразу для транскрибирования VIRF , гена, ответственного за экспрессию VIRF. VIRF является основным регулятором каскада вирулентности и экспрессируется из-за чувствительной к температуре области, изменяющей конформацию промотора VIRF, так что это больше не является благоприятным для ДНК-мостики с помощью H-NS ( рис. 3 ). [ 3 ] После экспрессии VIRF он регулирует производство ICSA , функционирует для повышения подвижности и VIRB , кодирует следующий регуляционный белок в каскаде шигеллы . Как только Virb экспрессируется, он нарушит H-NS для остальной части плазмиды вирулентности. [ 3 ]
Shigella spp. содержат «молекулярные резервные копии», или паралоги , для H-NS, которые были подробно изучены из-за их очевидной помощи в организации плазмиды вирулентности. [ 3 ] STPA -это паралог H-NS, который сохраняется по всем видам, но другой, SFH экспрессируется исключительно в штамме S. flexneri мутанта 2457T. [ 3 ] Этот мутантный штамм представляет большой интерес для исследователей, поскольку он действует как замена для H-NS, поскольку 2457T не содержит гена HNS . Корреляция между H-NS и его паралогами плохо изучена в это время. [ 3 ] Из-за важности этих паралогов в отсутствие H-NS у мутанта дальнейшие исследования и сосредоточение на этих паралогах могут привести к многообещающим антибактериальным методам лечения. [ 3 ]
Ссылки
[ редактировать ]- ^ Winardhi RS, Yan J, Kenney LJ (октябрь 2015). «H-NS регулирует экспрессию генов и уплотнения нуклеоида: понимание экспериментов с одной молекулярной» . Биофизический журнал . 109 (7): 1321–1329. Bibcode : 2015bpj ... 109.1321W . doi : 10.1016/j.bpj.2015.08.016 . PMC 4601063 . PMID 26445432 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т Цинь Л., Эркеленс А.М., Бен Бдира Ф., Дам Р.Т. (декабрь 2019 г.). «Архитекторы бактериальных ДНК -мостов: структурно и функционально консервативное семейство белков» . Открытая биология . 9 (12): 190223. doi : 10.1098/rsob.190223 . PMC 6936261 . PMID 31795918 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Picker MA, Wing HJ (декабрь 2016 г.). «H-NS, его члены семьи и их регуляция генов вирулентности у видов Shigella» . Гены . 7 (12): 112. doi : 10.3390/genes7120112 . PMC 5192488 . PMID 27916940 .
- ^ Shinddo H, Ieda T, Ieda R, Kurumizaka H, Ueguchi C, Minduno T, et al. (Февраль 1995). «Структура раствола связывающего ДНК домена ядерного белка, ассоциированного с ядерной, H-NS, из Escheria coli » Febs Письма 360 (2): 125–1 Doi : 10.1016/0014-5793 (95) 00079- O 7875316PMID S2CID 44479751
- ^ Bloch V, Yang Y, Margeat E, Chavanieu A, Auge MT, Robert B, et al. (Март 2003 г.). «Домен димеризации H-NS определяет новую складку, способствующую распознаванию ДНК». Природа структурная биология . 10 (3): 212–218. doi : 10.1038/nsb904 . PMID 12592399 . S2CID 25761309 .
- ^ Jump up to: а беременный Donato GM, Kawula TH (сентябрь 1998 г.). «Увеличенное связывание измененного белка H-NS с белком ротора жгутиков вызывает повышенную скорость вращения жгутиков и гипермотильность в Escherichia coli» . Журнал биологической химии . 273 (37): 24030–24036. doi : 10.1074/jbc.273.37.24030 . PMID 9727020 .
- ^ Lucchini S, Rowley G, Goldberg MD, Hurd D, Harrison M, Hinton JC (август 2006 г.). «H-NS опосредует молчание латерально приобретенных генов у бактерий» . PLO -патогены . 2 (8): E81. doi : 10.1371/journal.ppat.0020081 . PMC 1550270 . PMID 16933988 .
- ^ Дорман CJ (сентябрь 2014 г.). «H-NS-подобные нуклеоидные белки, мобильные генетические элементы и горизонтальный перенос генов у бактерий» . Плазмида . 75 : 1–11. doi : 10.1016/j.plasmid.2014.06.004 . PMID 24998344 .
- ^ Верма SC, Qian Z, Adhya SL (декабрь 2019 г.). «Архитектура я нуклеоид Escherichia coli» . PLOS Genetics . 15 (12): E1008456. doi : 10.1371/journal.pgen.1008456 . PMC 6907758 . PMID 31830036 .
- ^ Picker MA, Wing HJ (декабрь 2016 г.). «H-NS, его члены семьи и их регуляция генов вирулентности у видов Shigella» . Гены . 7 (12): E112. doi : 10.3390/genes7120112 . PMC 5192488 . PMID 27916940 .