Equisetidae
| Equisetidae Временной диапазон: | |
|---|---|
 | |
| Хвощ телматейский | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Трахеофиты |
| Разделение: | Полиподиофиты |
| Сорт: | Полиподиопсида |
| Подкласс: | Equisetidae Теплый. |
| Заказы | |
| Синонимы | |
См. текст. | |
Equisetidae — один из четырех подклассов Polypodiopsida девонскому (папоротников), группы сосудистых растений , летопись окаменелостей которых восходит к периоду . Они широко известны как хвощи . [2] Обычно они растут во влажных местах, с мутовками игольчатых ветвей, расходящихся через равные промежутки времени от одного вертикального стебля.
Equisetidae ранее рассматривались как отдельное подразделение споровых растений и назывались Equisetophyta , Arthrophyta , Calamophyta или Sphenophyta . При рассмотрении как класса названия Equisetopsida ss и Sphenopsida также использовались . Сейчас они признаны довольно близкими родственниками папоротников ( Polypodiopsida), из которых они образуют специализированную линию. [3] Однако разделение на хвощи и другие папоротники настолько древнее, что многие ботаники, особенно палеоботаники, до сих пор считают эту группу принципиально отдельной на более высоком уровне.
Описание
[ редактировать ]Хвощи представляют собой фотосинтезирующие «сегментированные» полые стебли, иногда наполненные сердцевиной. В месте соединения («узла», см. схему) между каждым сегментом находится мутовка листьев . У единственного сохранившегося рода Equisetum — это мелкие листья ( микрофиллы ) с единичным сосудистым следом, сросшиеся в оболочку в каждом узле стебля. Однако листья хвоща, вероятно, возникли в результате редукции мегафиллов , о чем свидетельствуют ранние ископаемые формы, такие как Sphenophyllum , у которых листья широкие с ветвящимися жилками. [4]
Сосудистые пучки разветвляются в узлах, при этом центральная ветвь становится жилкой микрофилла, а две другие перемещаются влево и вправо, чтобы слиться с новыми ветвями своих соседей. [5] сосудистых растений Сама сосудистая система напоминает систему эвстелы , которая развивалась независимо и конвергентно . [5] Очень быстрое удлинение междоузлий приводит к образованию сердцевинной полости и кольца каринальных каналов, образующихся в результате разрушения первичной ксилемы . Аналогичные пространства, валлекулярные каналы образуются и в коре. [5] Из-за более мягкой природы флоэмы их очень редко можно увидеть в ископаемых экземплярах. [ нужна ссылка ] У Calamitaceae вторичная ксилема (но не вторичная флоэма ) выделялась по мере роста камбия наружу, образуя древесный стебель и позволяя растениям вырасти до 10 метров в высоту. Все современные виды хвоща травянистые и утратили способность к вторичному росту. [5]
Подземные части растений состоят из сочленённых корневищ , из которых выходят корни и надземные оси. Растения имеют вставочные меристемы в каждом сегменте стебля и корневища, которые растут по мере того, как растение становится выше. Это контрастирует с большинством семенных растений, которые растут из апикальной меристемы, т.е. новый рост происходит только за счет роста кончиков (и расширения стеблей).
шишки (технически стробилы , поют. strobilus Хвощи несут на кончиках некоторых стеблей ). Эти шишки состоят из спирально расположенных спорангиеносцев , несущих по краям спорангии , а у современных хвощей покрывают споры снаружи — подобно мешочкам, свисающим с зонтика, ручка которого встроена в ось шишки. У вымерших групп дополнительную защиту спорам обеспечивало наличие мутовок прицветников — больших заостренных микрофиллов, выступающих из шишки.
Сохранившиеся хвощи гомоспоровые , но вымершие гетероспоровые виды, такие как Calamostachys Casheana . в летописи окаменелостей появляются [6] Спорангии открываются боковым раскрытием и высвобождают споры. Споры имеют характерные выступы , характерные пружинистые прикрепления, которые гигроскопичны : т. е. они меняют свою конфигурацию в присутствии воды, помогая спорам двигаться и способствуя их распространению.
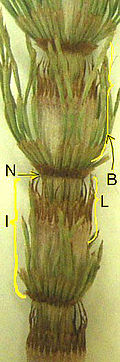
N = узел,
Я = междоузлия,
B = ветвь в мутовке,
L = сросшиеся микрофиллы


Таксономия
[ редактировать ]Классификация
[ редактировать ]Хвощи и их ископаемые родственники уже давно признаны отличными от других бессемянных сосудистых растений , таких как папоротники (Polypodiopsida). [7] До появления современных молекулярных исследований связь этой группы с другими живыми и ископаемыми растениями считалась проблематичной. [8] Из-за их неясных взаимоотношений ранги, присвоенные ботаниками хвощам, варьировались от отряда к отделу . При выделении в отдельный отдел в литературе используется множество возможных названий, в том числе Arthrophyta, [8] Каламофита, Сфенофита, [1] [9] или хвощёвые. Другие авторы рассматривали ту же группу как класс либо в составе отдела, состоящего из сосудистых растений, либо, в последнее время, в составе расширенной группы папоротников. При отнесении к классу группа получила название Equisetopsida. [10] или Сфенопсида. [5]
Современный филогенетический анализ , проведенный еще в 2001 году, продемонстрировал, что хвощи прочно принадлежат к папоротниковых группе сосудистых растений . [11] [12] Смит и др. (2006) провели первую классификацию птеридофитов более высокого уровня , опубликованную в эпоху молекулярной филогенетики , и считали, что папоротники (монилофиты) включают четыре класса , при этом хвощи относятся к классу Equisetopsida sensu stricto . [3] (Это различие необходимо из-за альтернативного использования Equisetopsida sensu lato как синонима всех наземных растений (Embryophyta) определенного класса. [13] ) Чейз и Ревил (2009) рассматривали хвощи как подкласс Equisetidae класса Equisetopsida sensu lato . Согласованная классификация, разработанная Группой Pteridophyte Phylogeny Group в 2016 году, также относит хвощи к подклассу Equisetidae, но к классу Polypodiopsida (папоротники в широком смысле). [2]
Филогения
[ редактировать ]На следующей диаграмме показаны вероятные филогенные отношения между подклассом Equisetidae и другими подклассами папоротников в соответствии с группой филогении Pteridophyte. [2]
| Полиподиопсида |
| ||||||
Исследование Elgorriaga et al., проведенное в 2018 году. предполагает, что отношения внутри Equisetidae показаны на следующей кладограмме. [14]
| |||||||||||||||||||||||||
По данным исследования, возраст кроновой группы хвощевых датируется как минимум ранним мелом , а скорее всего, юрским периодом . [14]
Подразделение
[ редактировать ]Подкласс Equisetidae содержит единственный сохранившийся отряд Equisetales . Этот порядок состоит из одного монотипного семейства Equisetaceae с одним родом Equisetum . Хвощ насчитывает около 20 видов. [12] [2]
Ископаемая запись
[ редактировать ]Сохранившиеся хвощи представляют собой лишь малую часть разнообразия хвощей прошлого. Существовало три отряда Equisetidae. Pseudoborniales . в позднем девоне впервые появились [1] Sphenophyllales . были доминирующим представителем каменноугольного подлеска и процветали до середины и перми начала Equisetales , но их разнообразие изменилось по мере того , существовали рядом со Sphenophyllales как эта группа исчезла, постепенно сокращаясь в разнообразии до сегодняшнего единственного рода Equisetum .
Впервые эти организмы появляются в палеонтологической летописи в конце девона. [1] время, когда наземные растения претерпевали быструю диверсификацию, а корни, семена и листья только что развились. (См. Эволюционную историю растений ). Однако растения существовали на земле уже почти сто миллионов лет, а первые свидетельства существования наземных растений датируются 475 миллионами лет назад . [15]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Тейлор, Теннесси и Тейлор, Эл (1993). Биология и эволюция ископаемых растений . Энглвуд Клиффс, Нью-Джерси: Прентис Холл. стр. 303–305. ISBN 978-0-13-651589-0 .
- ^ Перейти обратно: а б с д Группа филогении птеридофитов I (ноябрь 2016 г.), «Общественная классификация современных ликофитов и папоротников», Journal of Systematics and Evolution , 54 (6): 563–603, doi : 10.1111/jse.12229 , S2CID 39980610
- ^ Перейти обратно: а б Смит, Арканзас; Прайер, К.М.; Шуэттпельц, Э.; Коралл, П.; Шнайдер, Х. и Вольф, П.Г. (2006). «Классификация современных папоротников» (PDF) . Таксон . 55 (3): 705–731. дои : 10.2307/25065646 . JSTOR 25065646 . Архивировано из оригинала (PDF) 26 февраля 2008 г. Проверено 28 октября 2019 г.
- ^ Рутисхаузер, Р. (1999). «Полимерные мутовки листьев сосудистых растений: морфология развития и нечеткость особенностей органов». Международный журнал наук о растениях . 160 (6): 81–103. дои : 10.1086/314221 . JSTOR 10.1086/314221 . ПМИД 10572024 . S2CID 4658142 .
- ^ Перейти обратно: а б с д и Стюарт, В.Н. и Ротвелл, Г.В. (1993). Палеоботаника и эволюция растений (2-е изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-38294-6 .
- ^ Бауэр, Ф.О. (1959) [1935]. Происхождение наземной флоры, теория, основанная на фактах чередования . Нью-Йорк: Hafner Publishing Co. p. 381.
- ^ Имс, Эй Джей (1936). Морфология сосудистых растений (Низшие группы) . Нью-Йорк и Лондон: Книжная компания McGraw-Hill. стр. 110–115.
- ^ Перейти обратно: а б Смелый, ХК; Алексопулос, CJ; Делеворяс, Т. (1987). Морфология растений и грибов (5-е изд.). Нью-Йорк: Харпер-Коллинз. стр. 371–387, 478, 506–514. ISBN 978-0-06-040839-8 .
- ^ Гиффорд, EM; Фостер, А.С. (1988). Морфология и эволюция сосудистых растений (3-е изд.). Нью-Йорк: WH Freeman and Company. стр. 175–207 . ISBN 978-0-7167-1946-5 .
- ^ Кенрик, П.; Крейн, PR (1997). Происхождение и раннее разнообразие наземных растений: кладистическое исследование . Вашингтон, округ Колумбия: Издательство Смитсоновского института. стр. 241–242. ISBN 978-1-56098-730-7 .
- ^ Прайер, К.М.; Шнайдер, Х.; Смит, Арканзас; Крэн, Р.; Вольф, П.Г.; Хант, Дж. С.; Сайпс, С.Д. (2001). «Хвощи и папоротники — монофилетическая группа и ближайшие ныне живущие родственники семенных растений» (PDF) . Природа . 409 (6820): 618–621. Бибкод : 2001Natur.409..618S . дои : 10.1038/35054555 . ПМИД 11214320 . S2CID 4367248 . Архивировано из оригинала (PDF) 2 сентября 2009 г.
- ^ Перейти обратно: а б Кристенхуш, Мартен Дж. М. и Чейз, Марк В. (2014). «Тенденции и понятия в классификации папоротников» . Анналы ботаники . 113 (4): 571–594. дои : 10.1093/aob/mct299 . ПМЦ 3936591 . ПМИД 24532607 .
- ^ Чейз, Марк В. и Ревил, Джеймс Л. (2009), «Филогенетическая классификация наземных растений, сопровождающих APG III», Ботанический журнал Линнеевского общества , 161 (2): 122–127, doi : 10.1111/j. .1095-8339.2009.01002.x
- ^ Перейти обратно: а б Эльгорриага, А.; Побег, IH; Ротвелл, GW; Томеску, АМФ; Кунео, Северная Каролина (2018). «Происхождение Equisetum: эволюция хвощей (Equisetales) внутри основной клады эуфиллофитов Sphenopsida» . Американский журнал ботаники . 105 (8): 1286–1303. дои : 10.1002/ajb2.1125 . ПМИД 30025163 .
- ^ Веллман, Швейцария; Остерлофф, Польша; Мохиуддин, У. (2003). «Фрагменты древнейших наземных растений» (PDF) . Природа . 425 (6955): 282–285. Бибкод : 2003Natur.425..282W . дои : 10.1038/nature01884 . ПМИД 13679913 . S2CID 4383813 .
![]() Данные, относящиеся к Equisetopsida, на Wikispecies
Данные, относящиеся к Equisetopsida, на Wikispecies
Классификация Archaeplastida или Plantae sl |
|---|