Тапиноканины
| Тапиноканины Временный диапазон:
| |
|---|---|
| |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Synapsida |
| Клада : | Терапсида |
| Подотряд: | † Dinocephalia |
| Семья: | † Tapinocephalidae |
| Подсемейство: | † Тапиноканинин |
| Род: | † Тапиноканины Рубидж, 1991 |
| Разновидность: | † Т. Памела
|
| Биномиальное название | |
| † Tapinocaninus pamelae Рубидж, 1991
| |
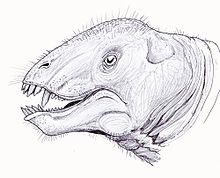
Tapinocaninus (греческий для «скромного»- тапино и «собака»- caninus) является в семействе Tapinocephalidae вымершим родом терапидов , из которых он является наиболее базальным членом. Только один вид известен, Tapinocaninus pamelae (что означает «скромный собачий Пэм»). Вид назван в честь матери Рубиджа, Пэм. [ 1 ] Окаменелости были найдены, датируемые средним пермцем ( эпохи Вордиана ).
Открытие
[ редактировать ]Окаменелости тапиноканина были впервые обнаружены в зоне сборки Эодицинодона в отложениях Кару, в нижних ложе Бофорта в Бофорте Запад. Известно пять образцов, четыре найденных на ферме Modderdrift и один из них найден на ферме Swartgrond. Голотип (NMQR 2987) и четыре паратипа (NMQR 2985, 2986, 3097 и Roz K95). [ 1 ]
Два экземпляра были обнаружены директором Института палеонтологических исследований Бернарда Прайса (в настоящее время Институт эволюционных исследований) профессором Брюсом Рубиджем. Три были найдены Джоном Ньяплули из Национального музея Блумфонтейна в том же песчаном камне. Раскопки и подготовка NMQR 2986 Голотип NMQR 2987 был большим мероприятием из -за большого размера образцов, и этот процесс имел место с 1985 по 2005 год. Для приготовления образцов использовался механизм воздушного писца, а также ручные инструменты, такие как инструменты, как инструменты, такие как ручные инструменты. молоток и долото в районах, где матрица была более распространенной. [ 1 ] [ 2 ]
Перед открытием тапиноканина , как считалось , самая примитивная диноцефалина, а тапиноцефалин - самый примитивный диноцефалиан в Южной Африке. При сравнении особенностей тапиноканина с теми, которые обсуждались о Tapinoehalinae с помощью кладистического анализа, Rubidge et al. (1991) обнаружили, что синапоморфией из них была расширенными каблуками на зубах резца. [ 3 ] Таким образом, после его открытия тапиноканин считается наиболее примитивным тапиноцеохалином. [ 1 ]
Описание
[ редактировать ]Этот вид известен из нескольких черепов, так как большинство обнаруженных образцов не хватало скелета после черепа. Это было крупное животное, имеющее длину 2,5 м от морды до подвздошного костюма. [ 2 ] С классическими формулами регрессии с использованием окружности плечевой кости и костей бедра, исследователи приблизили, что они в среднем составляли массу тела 892,63 кг для таксона (почти 2000 фунтов). [ 4 ] Тапиноканин также были самыми большими терапидами от гуадалупиан . [ 5 ]
Череп
[ редактировать ]Крыша черепа и посторбитальный батончик тапиноканина показывают пахиостотическое утолщение, что согласуется с другими диноцефальцами тапиносефалина. Крыша черепа в основном состоит из фронтала , которая простирается между орбитальным и височным фенестром . Внешний нарис граничит с дорсально, спереди и переднеровентрально костью предчезгил . Верхтка образует большую часть боковой области лица, и она опухла, чтобы обеспечить место для корня собачьего зуба. [ 6 ] Временные отверстия относительно большие, а впоследствии тапиноканин имеет узкую межвременную область, которая считается примитивной особенностью Tapinocephalinae . Временные фенестра граничат вентрально и сзади сквомозалом, а дорсально по посторбиталам. Сквамосовый и посторбитальный штрих вдоль височного отверстия, который является особенностью, обычно встречающейся в тапиносефалидах. [ 7 ] Кроме того, эти таксон имеет тонкую морду, наклонную затылочную, относительно маленькую квадратоджугальную, выдающуюся стапедиальную отверстие и относительно переднее положение квадрата. [ 1 ]
Не вдоль
[ редактировать ]тапиноканина В небе есть узкие рваные рты с гребнем между ними , который окружает внутренние NARE. Пенсисилла, которая перекрывает верхнюю челюсть спереди и вентрально, и верхняя челюсть контактирует как с палатиновым, так и птеригоидным медиально. Более половины длины неба - птеригоид, и имеет боковые фланцы, которые простираются вентрально и поперечно. Кроме того, птеригоид касается базисфеноида за межптеригоидной вазостью. [ 1 ]
Позвонки
[ редактировать ]Позвонки тапиноканина отличаются от их боковой обработки фовеа у основания нервного позвоночника. Эта особенность является примитивной для TherapsID, еще одна наследственная черта для этого клады также включает в себя наличие интерцентра между шейками -позвонками. Тапиноканинус имеет 36 позвонков, 8 шейки матки , 20 дорсалов , 5 поясниц , 2 связки и 7 хвостах . У них есть длинный нервный позвоночник, который простирается до каудальных позвонков. [ 2 ] Их структура позвонка предполагает, что у тапиноканина были очень короткие хвосты, похожие на аномодонты , но в отличие от антеозавридов . [ 8 ] Присутствие только двух сакральных позвонков в тапиноканине отличается от других терапидов, таких как мосшопы , которые имеют три. [ 9 ]
Ребра
[ редактировать ]Вдоль всей позвоночной колонки присутствуют ребра. В шейной области ребра укорочены и плоские. Самые длинные ребра присутствуют в средней части дорсальной области, а спинные ребра 11-15 имеют ствол в форме ствола, чтобы вместить большую пищеварительную систему тапиноканина . В каудальной области ребра снова укорочены, в дополнение к дорсовентрально сплющенным, сзади направлено и не слиты с хвостовыми позвонками [ 2 ]
Групный пояс
[ редактировать ]Групный пояс тапиноканина имеет выдающуюся лопасть, с увеличенным дорсальным концом. Гленоид прямой, вентральная обработка и утолщен, чтобы содержать коракоиды и лопасть. Гребник проходит по диагонали от заднего дорсального конца гленоида до переднего конца лопатки, и это ограничивает движение ключицы над лопатой. Лопатка также вспыхивает, чтобы соединиться с передним коракоидом, который представляет собой круглую большую кость. [ 2 ]
Тазовый пояс
[ редактировать ]Тазовый пояс голотипа NMQR 2987 частично сохранен, а другие образцы также отсутствуют полной структуры. Тем не менее, параллельные канавки обнаружены на поверхности ишемира, что предполагает, что они могли быть связаны с помощью ткани. [ 9 ]
Плечевая кость
[ редактировать ]Три гуммери были сохранены из различных образцов тапиноканина , хотя правильная плечевая кость голотипа является наиболее полным и хорошо описанным в исследованиях. Кость имеет узкий центр и расширяется на концах, а проксимальный конец больше дистального. Дельтопекторный гребень вспыхивает и составляет большую часть длины кости. Дистальный конец плечевой кости имеет небольшой энтепикондиль и больший эктопикондиль с ямкой, разделяющей два. [ 2 ] Тапиноканин отличается от Ulemosaurus и Moschops тем, что он имеет энтепикондиларный отверстие, в то время как последние два таксона имеют эктипикондилар и энтепикондилар. [ 3 ] [ 10 ]
Бедро
[ редактировать ]Голотип NMQR 2987 имеет как левую, так и правую бедру. Этот образец показывает, что бедра более стройная, чем плечевая кость. Бедра та же ширина на проксимальных и дистальных концах, и она сплющена передне. Подобно другим тапиносефалидам, тапиноканинус имеет медиальногированную головку бедренной кости. На дистальном конце бедренной кости есть боковые и медиальные мыщелки , а боковой мыщелок немного больше по размеру. [ 2 ]
Зубной зуб
[ редактировать ]Несмотря на то, что в настоящее время нет образца с идеально сохранившимися зубами, зубное описание для тапиноканина было взято из различных черепов (NMQR 2984, NMQR 2986, NMQR 2987, Roz K95). У этого диномефуфяна есть гетеродонт , состоящий из резков, клыков и постов. Клыки. В премьсилле присутствует максимум пять зубов резца, которые демонстрируют морфологию Talon и пятки. Единственный собака - первый зуб в верхней челюсти, он изогнут назад и не имеет каблука. Следуйте собачьим зубами, характеризующимися заостренными коронками и небольшими, языковыми каблуками. Расточие зубцы указывают на то, что эти животные, вероятно, были травоядными или всеядными. [ 1 ]
Палеобиология
[ редактировать ]Осанка и передвижение
[ редактировать ]Исследователи предполагают, что осанка тапиноканина предшествует , чтобы быть «промежуточным» между обширным и вертикальным положением. Предполагается, что они более честны, чем сфенакодонты, но более растянуты, чем Theroidont Theraspids. Их промежуточная поза можно объяснить их длинными костями, в дополнение к форме и позиционированию плечевой кости и медиальногированной бедренной кости. Эта постуральная позиция также будет более поддерживающей больший размер тела тапиноканина . [ 2 ] Кроме того, присутствие интерцентра только в передних дорсальных позвонках и медиальных режиссеры зигапофиза в тапиноканине предполагает, что они имели менее неравноводную локомоцию по сравнению со спгенакодонтами. [ 11 ]
Кормление
[ редактировать ]Гетеродонтный зубной ряд указывает на то, что зубы морфологически дифференцируются по форме. [ 12 ] В тапиноканине это включает в себя резцы, собаки и клыки, все из которых имеют разные формы, которые позволяют выполнять различные функции. Тапиноканин, вероятно, был травоядным или плотоядным животным. [ 1 ]
Геологическая и биостратиграфическая информация
[ редактировать ]Эодицинодона находится в юго-западной части бассейна Кару, и она является самой низкой Зона сборки биозоной группы Бофорта, под эозимопс-гланосухуса субзоном зоны сборки Тапиноцефала . Нижняя граница лежит на стратиграфическом горизонте между группой ECCA (Waterford Formation) и группой Бофорта (формация Абрахамскаала), которую исследователи Рубидж и Оелофсен считали палеохорной линией. [ 13 ] Он был назван Рубейдом в 1995 году после наиболее распространенного терапида, присутствующего в этом районе, эодицинодон Оостхюзени . Нижняя граница устанавливается первым появлением эодицинодонского оосхуйзени в зоне, а верхняя граница - первым появлением Eosimops Newtoni , дицинодонта. В боковом направлении биозона бежит из Лайнгсбурга к югу от Ритброна. [ 14 ]
Зона идентифицируется по существованию эодицинодона Оостхюзени , дицинодонта , наряду с тапиноканином Памела и Австралоодон Ньяпля , два типа диномефальцев . Окаменелости тапиноканина составляют приблизительно 10% всех ископаемых тетрапода, обнаруженных в этой биозоне. Толщина этой биозоны колеблется от 320 до 1100 м, с максимальной толщиной на станции Принц Альберт -роуд и постепенно истончение к востоку и западу от этой точки. Биозона состоит из алевок, песчаников и грязевых камней. Окаменелости с тетраподами редки в этой области, однако существуют обильные впечатления о эквизеталовых и глоссоптеризах и листьях на поверхностях мудряка. При обнаружении окаменелости TherapsId обычно сохраняются в грязных скалах, а диномефальцы встречаются в мелкозернистого песчаника. [ 14 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час Рубидж, Б.С. (1991). «Новая примитивная диноцефальная рептилия, похожая на млекопитающие, от Пермия Южной Африки» . Палеонтология . 34 : 547–559.
- ^ Jump up to: а беременный в дюймовый и фон глин час Рубидж, Брюс С.; Говендер, Ромала; Романо, Марко (2019-10-18). «Посткраниальный скелет базального тапиноцефалидного диноцефального тапиноканина Pamelae (Synapsida: Therapsida) из южноафриканской супергруппы Karoo» . Журнал систематической палеонтологии . 17 (20): 1767–1789. doi : 10.1080/14772019.2018.1559244 . ISSN 1477-2019 .
- ^ Jump up to: а беременный Кинг, GM 1988. Аномодонтия. 1-174. В Wellnhofer, P. (ed.). Энциклопедия палеохерпетологии, 17с. Густав Фишер, Штутгарт.
- ^ Романо, М. и Рубидж, Б. (2019). Первая трехмерная реконструкция и объемная масса тела массы тапиноцефалидного диноцефального тапиноканина Pamelae (Synapsida: Therapsida). Историческая биология, 33 (4), 498–505. https://doi.org/10.1080/08912963.2019.1640219
- ^ Сидней), Рубидж, Б.С. (Брюс (1995). млекопитающие в Африке? Совместно ли 0-86813-160-1 Полем OCLC 813446450 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Boostra, Lieuwe Dirk (1962). Зубная зубца титаносухинских диноцефалинов (48 изд.). Анналы южноамериканского музея. С. 233–236.
- ^ Гювен, С. и Рубидж, Брюс и Абдала, Фернандо. (2013). : 48. 24-3
- ^ Говендер, Ромала; Рубидж, Брюс С. (июль 2002 г.). «Первая полная позвоночная колонна базальной тапиноцефалидной диноцефальной (Synapsida: Therapsida)» . Южноафриканский журнал науки . 98 (7): 391–392 - через африканские журналы Sabinet.
- ^ Jump up to: а беременный Грегори, Уильям К.; Метла, Роберт (1926). «Скелет Moschops Capensis Broom, рептилия Dinocephalian от Pommian в Южной Африке» . Бюллетень Американского музея естественной истории . 56 (56): 179–251.
- ^ Riabinin, 1938. Фауна позвоночных из верхних пермских отложений бассейна Sviaga. I. Новый Dinocephalian Ulemosaurus svijagenesis ngnsp. Анналы Центрального геологического и проспективного музея научных исследований (Музей Чернишью), 1, 1-75.
- ^ Kemp, TS 2005. Происхождение и эволюция млекопитающих. Издательство Оксфордского университета, Оксфорд, 331 стр.
- ^ Kemp, TS, & Kemp, DF (1982). Млекопитающие рептилии и происхождение млекопитающих . Академическая пресса.
- ^ Rubidge, BS and Oelofsen, BW, 1981. Рептильная фауна из скал Ecca возле принца Альберта, Южная Африка. Южноафриканский журнал науки 77, 425-426.
- ^ Jump up to: а беременный Рубидж, BS; День, Мо (2020-06-01). «Биостратиграфия зоны сборки Эодицинодона (Beaufort Group, Supergroup Karoo), Южная Африка» . Южноафриканский журнал геологии . 123 (2): 141–148. doi : 10.25131/sajg.123.0010 . ISSN 1996-8590 .
Внешние ссылки
[ редактировать ]- Премер и Хепер


Эта статья, связанная с TherapsId, является заглушкой . Вы можете помочь Википедии, расширив ее . |