Pampaphoneus
| Pampaphoneus Временный диапазон: Гвадалупиан ( Вордиан ),
| |
|---|---|

| |
| Pampaphoneus biccai голотип черепа | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Synapsida |
| Клада : | Терапсида |
| Подотряд: | † Dinocephalia |
| Семья: | † anteosauridae |
| Подсемейство: | † syodontinae |
| Род: | † Pampaphoneus Cisneros et al. 2012 |
| Тип видов | |
| † Pampaphoneus biccai Cisneros et al. 2012
| |
Pampaphoneus является вымершим родом плотоядного диноцефальского TherapsId, принадлежащего к семейству Anteosauridae . Он жил от 268 до 265 миллионов лет назад в эпоху Гвадалупианского Фордиана ( в том , = среднего пермского ) периода что сейчас является Бразилии . Pampaphoneus известен почти полным черепом с нижней челюстью, все еще сформулированной, обнаруженной на землях фермы Бокирау, недалеко от города Сан -Габриэль , в штате Рио -Гранд -до Сул . В 2019 и 2020 годах был зарегистрирован второй образец из той же местности, но еще не был описан. Он состоит из черепа, связанного с посткраниальными останками. Это первый южноамериканский вид диномефальянского, который был описан. Группа была ранее известна в Южной Америке только несколькими изолированными зубами, а фрагмент челюсти сообщил в 2000 году в том же регионе Бразилии. циснеросом и проведенный , Филогенетический анализ коллегами Южная Америка и Восточная Европа , чем предполагалось ранее, способствуя реконструкции Continental Pangea B.
Этимология
[ редактировать ]Название рода происходит от Пампаса , области обширных равнин, типичная для Южной Южной Америки, из которой возникает образец, и от греческого телефона , что означает «убийца», в отношении хищных привычек животного. Конкретный эпитет чтит Хосе Бикку, землевладельца фермы, где было найдено ископаемое. [ 1 ]
Описание
[ редактировать ]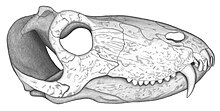
The holotype (UFRGS PV386P) of Pampaphoneus is a skull measuring approximately 32 cm in length. As with all anteosaurs, the ventral margin of the premaxilla is inclined upwards and the postorbital bar is strongly curved anteroventrally so that the temporal fenestra undercuts orbit. The skull roof is slightly pachyostosed. The temporal fenestrae open widely on the skull roof where the insertion zone of the adductor musculature of the mandible extends to the pineal boss. The frontals contribute to the insertion zone of this musculature and also participate to the anterior edge of the pineal boss. These characteristics indicate that Pampaphoneus belongs to the clade Syodontinae. Within this group it is with the Russian species Syodon biarmicum that Pampaphoneus shares the most similarities. Like the latter, it possesses strongly recurved hook-like canines and very low postcanines with serrassions. The canines of the holotype are 7 cm in length. Pampaphoneus however differs from Syodon by its larger size, more robust snout, thickened postorbital forming a supraorbital boss, and a well-developed ridge extending from the pineal boss to the orbital rim. It also differs from other anteosaurs by its squamosal with a jugal process extending beyond the anterior edge of the temporal fenestra.[1] A peculiarity of the holotype that was thought to be a diagnostic feature is the presence of only four teeth per premaxilla (the fourth incisor, small in size, being however laterally masked by the maxilla) and eight postcanines.[1] However, a second skull (yet undescribed) was reported showing some differences with the holotype such as the presence of five premaxillary teeth and only seven postcanines. Since the other cranial elements are very similar, these differences are interpreted as intraspecific variation.[2][3] Although being a Syodontinae, Pampaphoneus also shares several characters with the more derived Anteosaurinae. It thus has a well-developed medial crest on the skull roof, while a pronounced thickening of each postorbital forms a supraorbital boss similar to that of a subadult individual of the Russian Titanophoneus potens. In addition, the angular bone of the lower jaw bears a boss as in Anteosaurus and the two species of Titanophoneus. This angular boss is however much less developed in Pampaphoneus.[1]
Paleoecology
[edit]
Remains of Pampaphoneus comes from the Morro Pelado Member of the Rio do Rasto Formation, outcropping on the lands of the Boqueirão farm in the municipality of São Gabriel (district of Catuçaba, State of Rio Grande do Sul). The fossils are preserved there in a fine pinkish sandstone and are covered with a dark crust of iron oxide.[4] Sediments of the Morro Pelado Member were deposited in fluvio-lacustrine and deltaic settings suggesting alluvial conditions with coalescing floodplains. Aeolian facies (fossil dunes) present in the upper part of the Morro Pelado Member attest to a progressive aridification of the environment. Palaeogeographically, southeastern Brazil was located at mid-latitudes between the 35th and 45th parallel south, where a warm temperate climate probably prevailed with a prolonged dry season.[5] The flora included forests of Glossopteris communis which occupied the floodplains and the overbank channels while the more humid biotopes were covered with dense mats of equisetids including equisetales (Schizoneura and Phyllotheca ) and sphenophyllales (Sphenophyllum paranaense).[5] Besides Pampaphoneus, the Boqueirão farm site also contains the temnospondyl Konzhukovia sangabrielensis[6] and the small dicynodont Rastodon.[4] Other localities in the Morro Pelado Member have also yielded tetrapod remains. In Rio Grande do Sul, several sites around Aceguá in the district of Minuano have yielded the most complete fossils of the pareiasaur Provelosaurus.[7] The Fagundes farm locality (São Gabriel, Catuçaba district) yielded a fragmentary skull of Provelosaurus,[8] teeth of undetermined Anteosaurid, Titanosuchid and Tapinocephalid dinocephalians,[9] and an undetermined amphibian.[10] In the district of Tiarajú (also near São Gabriel) was found the basal anomodont Tiarajudens[11][12] From the Serra do Cadeado area (near Ortigueira, State of Paraná), come the amphibian Australerpeton,[10] an undetermined species of the dicynodont Endothiodon,[13] (possibly E. bathystoma[14]) as well as an indeterminate tapinocephalid dinocephalian showing similarities with the genera Moschops and Moschognathus.[10] However, there are uncertainties about the contemporaneity of all these sites. Vertebrate fossils from the Rio do Rasto Formation occur in scattered, isolated, and discontinuous outcrops due to dense vegetation cover, making it difficult to establish local correlations. The precise location of several ancient discoveries is uncertain and several taxa could come from different stratigraphic levels within the Rio do Rasto Formation. This is the case of the fossils of Paraná, where tapinocephalid and Endothiodon remains could come from two distinct levels.[10]
Age of the Rio do Rasto Formation
[edit]Estimating the age of the Rio do Rasto formation has long relied solely on the basis of biostratigraphic correlations. Since 2018, radiometric dating has made it possible to specify the age of part of the formation.[15][16] Based on conchostracan and bivalve faunas, Holz and colleagues have suggested that the sedimentary succession of this formation extended from the Wordian (middle Guadalupian) to the Wuchiapingian (base of the Lopingian).[17] Among the tetrapods, the presence of dinocephalians in Fagundes and Boqueirão sites (State of Rio Grande do Sul) as well as in the region of Serra do Cadeado (State of Paraná) indicates a Guadalupian age for these localities, given that this synapsid group is restricted to this age in the rest of the world. In the Fagundes farm site, dinocephalians coexist with the pareiasaur Provelosaurus. The latter is present in the Aceguà area about 2 m above a 30 cm thick layer of bentonite radiometrically dated at 266 ± 5.4 million years.[16][7] The broad error margin of this radiometric dating places the Brazilian dinocephalian sites in the Lower Roadian – Middle Capitanian time interval, confirming the Guadalupian age of this part of the Morro Pelado Member.[7] The other tetrapods of the Morro Pelado Member in Rio Grande do Sul suggest an age not exceeding Wordian: Tapinocephalids are only known from Wordian and Capitanian rocks, the basal anomodont Tiarajudens is closely related to the South African genus Anomocephalus of the Tapinocephalus Assemblage Zone whose age extends from the late Wordian to the late Capitanian, and the temnospondyl Konzhukovia, present at the Boqueirão farm site in São Gabriel, is also known in eastern European Russia in strata limited to the Wordian-Capitanian interval.[7] The probable Wordian age of the Morro Pelado Member is also reinforced by a radiometric age of the Serrinha Member (lower part of the Rio do Rasto formation) obtained from a volcanic ash layer which gave an age of 270.61 + 1.76/− 3.27 Ma corresponding to the Roadian (early Guadalupian).[15]
Classification
[edit]In describing Pampaphoneus, Cisneros et al. presented several cladograms confirming the recognition of the clades Anteosaurinae and Syodontinae erected a year earlier by Christian Kammerer. In all their analyzes, Pampaphoneus is identified as the most basal Syodontinae:[1]
Palaeobiogeography
[edit]During the Permian, most of the landmasses were united into a single supercontinent, Pangea, which was roughly C-shaped. Its northern (Laurasia) and southern (Gondwana) parts were connected to the west but separated to the east by a very large oceanic bay - the Tethys.[18] A long string of microcontinents, grouped under the name of Cimmeria, divided the Tethys in two: the Paleo-Tethys in the north, and the Neo-Tethys in the south.[19] In the northern hemisphere, anteosaurs are known from eastern European Russia, Kazakhstan, and China, territories corresponding to mid-paleolatitudes between the 30th and 40th parallel north.[20] In the southern hemisphere, anteosaur remains are known in South Africa, Zambia and Zimbabwe, territories which were all located at high paleolatitudes located at the level of the 60th parallel south.[20][21] The Brazilian localities were located a little further north, between the 35th and the 45th parallel south.[5] The close relationship of the Brazilian Pampaphoneus with some South African and Russian anteosaurs suggests dispersal through western Pangea rather than eastern Pangea via the Cathaysian Bridge. The latter included parts of northern and southern China, Korea, and Indochina and played an important role in the dispersal of therapsids in the late Permian and Triassic. However, this bridge was probably not yet formed in the Middle Permian and marine barriers broke up the different Cathaysian blocks, making it difficult for animals to disperse from Cathaysia to Southern Africa via Eastern Europe. Cisneros and colleagues suggest a seemingly easier migration route via western Pangea by favoring a Pangea B-type continental reconstruction where South America was juxtaposed with the Appalachia and was thus closer to Russia than in the Pangea A reconstruction. In the latter, the Appalachian and Mauritanides mountains would have been difficult natural barriers to cross. In contrast, in a Pangea B configuration, Brazil was not only closer to Eastern Europe, but the only mountain barrier along the way was the moderately high European Hercynides. Anteosaurs would have migrated from Russia (where the oldest specimens come from) to southern Africa, passing through eastern Europe and western Africa (bypassing the Mauritanides chain) and then through Brazil.[1] The discovery of probable anteosaur footprints in southern France,[N 1] which was then located at low paleolatitude, at the level of the 10th parallel north, also supports this hypothesis, the south-western Europe being in this migration corridor.[22][23][24][25]
Notes
[edit]- ^ These footprints, initially named Planipes caudatus and P. brachydactylus, are now grouped under the name Brontopus antecursor (cf. references Marchetti et al. 2019).
References
[edit]- ^ Jump up to: a b c d e f Cisneros, J.C.; Abdala, F.; Atayman-Güven, S.; Rubidge, B.S.; Şengör, A.M.C.; Schultz, C.L. (2012). "Carnivorous dinocephalian from the Middle Permian of Brazil and tetrapod dispersal in Pangaea" (PDF). Proceedings of the National Academy of Sciences of the United States of America. 109 (5): 1584–1588. Bibcode:2012PNAS..109.1584C. doi:10.1073/pnas.1115975109. PMC 3277192. PMID 22307615.
- ^ Anilson Costa Santos, M.; Bulsing, K.P.; Santos, A.T.; Ferraz, J.S.; Oliveira, N.; Protti, S.W.L.; Dos Santos Rodriguez, L.A.; Xavier, P.L.; De Queiroz, L.C.; Cisneros, J.C.; Pinheiro, F.L. (2019). "Um Esqueleto Parcial do Dinocefálio Anteossaurídea Pampaphoneus biccai (Permiano do Rio Grande do Sul)". Anais do XXVI Congresso Brasileiro de Paleontologia. Campinas: Galoá. 2: 270–271.
- ^ Perceval Paz, C.; Anilson Costa Santos, M.; Lima Pinheiro, F. (2020). "O Primeiro Registro de Pampaphoneus biccai (Anteosauridae) Com Ossos Pós-Cranianos". Revista Multidisciplinar de Educação e Meio Ambiente. 1 (1): 133.
- ^ Jump up to: a b Boos, A.D.S.; Kammerer, C.F.; Schultz, C.L.; Soares, M.B.; Ilha, A.L.R. (2016). "A New Dicynodont (Therapsida: Anomodontia) from the Permian of Southern Brazil and Its Implications for Bidentalian Origins". PLOS ONE. 11 (5): e0155000. Bibcode:2016PLoSO..1155000B. doi:10.1371/journal.pone.0155000. PMC 4880204. PMID 27224287.
- ^ Jump up to: a b c Cúneo, N.R. (1996). "Permian phytogeography in Gondwana". Palaeogeography, Palaeoclimatology, Palaeoecology. 125 (1–4): 75–104. Bibcode:1996PPP...125...75C. doi:10.1016/S0031-0182(96)00025-9.
- ^ Pacheco, C.P.; Eltink, E.; Müller, R.T.; Dias-da-Silva, S.A. (2016). "A new Permian temnospondyl with Russian affinities from South America, the new family Konzhukoviidae, and the phylogenetic status of Archegosauroidea". Journal of Systematic Palaeontology. 15 (3): 241–256. doi:10.1080/14772019.2016.1164763. S2CID 87860271.
- ^ Jump up to: a b c d Cisneros, J.C.; Dentzien-Dias, P.; Francischini, H. (2021). "The Brazilian Pareiasaur revisited". Frontiers in Ecology and Evolution. 9: 758802. doi:10.3389/fevo.2021.758802.
- ^ Cisneros, J.C.; Abdala, F.; Malabarba, M.C. (2005). "Pareiasaurids from the Rio do Rasto Formation, southern Brazil: Biostratigraphic implications for Permian faunas of the Paraná Basin". Revista Brasileira de Paleontologia. 8 (1): 13–24. doi:10.4072/rbp.2005.1.02.
- ^ Langer, M.C. (2000). "The first record of dinocephalians in South America: Late Permian (Rio do Rasto Formation) of the Paraná Basin, Brazil". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 215: 69–95. doi:10.1127/njgpa/215/2000/69.
- ^ Jump up to: a b c d Boos, A.D.S.; Kammerer, C.F.; Schultz, C.L.; Paes Neto, V.D. (2015). "A tapinocephalid dinocephalian (Synapsida, Therapsida) from the Rio do Rasto Formation (Paraná Basin, Brazil): Taxonomic, ontogenetic and biostratigraphic considerations". Journal of South American Earth Sciences. 63: 375–384. Bibcode:2015JSAES..63..375B. doi:10.1016/j.jsames.2015.09.003.
- ^ Cisneros, J.C.; Abdala, F.; Rubidge, B.S.; Dentzien-Dias, P.C.; Bueno, A.O. (2011). "Dental Occlusion in a 260-Million-Year-Old Therapsid with Saber Canines from the Permian of Brazil". Science. 331 (6024): 1603–1605. Bibcode:2011Sci...331.1603C. doi:10.1126/science.1200305. PMID 21436452. S2CID 8178585.
- ^ Cisneros, J.C.; Abdala, F.; Jashashvili, T.; Bueno, A.O.; Dentzien-Dias, P. (2015). "Tiarajudens eccentricus and Anomocephalus africanus, two bizarre anomodonts (Synapsida, Therapsida) with dental occlusion from the Permian of Gondwana". Royal Society Open Science. 2 (7): 1–27. Bibcode:2015RSOS....250090C. doi:10.1098/rsos.150090. PMC 4632579. PMID 26587266.
- ^ Boos, A.D.S.; Schultz, C.L.; Vega, C.S.; Aumond, J.J. (2013). "On the presence of the Late Permian dicynodont Endothiodon in Brazil". Palaeontology. 56 (4): 837–848. doi:10.1111/pala.12020. S2CID 129679835.
- ^ Kammerer, C.F.; Ordoñez, M.A. (2021). "Dicynodonts (Therapsida: Anomodontia) of South America". Journal of South American Earth Sciences. 108: 103171. Bibcode:2021JSAES.10803171K. doi:10.1016/j.jsames.2021.103171. S2CID 233565963.
- ^ Jump up to: a b Francischini, H.; Dentzien-Dias, P.; Guerra-Sommer, M.; Менегат, Р.; Сантос, Джос; Manfroi, J.; Schultz, CL (2018). «Средняя пермская (дорожная) эстивация легких рыбков из формации Рио-до Расто (бассейн Парана, Бразилия) и связанные с ними знакомства с U-PB». Палаис . 33 (2): 69–84. Bibcode : 2018palai..33 ... 69f . doi : 10.2110/palo.2017.050 . S2CID 134435628 .
- ^ Jump up to: а беременный Rocha-Campos, AC; Basei, mas; Натман, AP; Santos, Pr; Пассарелли, кр .; Canile, FM; Роза, OCR; Фернандес, MT; Санта -Ана, Х.; Верославский, Г. (2019). «U-PB Циркон датируется отложениями пепла из палеозойского бассейна Парана в Бразилии и Уругвае: переоценка стратиграфических корреляций». Журнал геологии . 127 (2): 167–182. Bibcode : 2019jg .... 127..167r . doi : 10.1086/701254 . HDL : 1885/202604 . S2CID 134671451 .
- ^ Holz, M.; Франса, AB; Souza, PA; Яннуцци, Р.; Рон Р. (2010). «Стратиграфическая карта поздней каменноугольной/пермской преемственности восточной границы бассейна Парана, Бразилия, Южная Америка». Журнал южноамериканских наук о Земле . 29 (2): 381–399. Bibcode : 2010jsaes..29..381h . doi : 10.1016/j.jsames.2009.04.004 .
- ^ McLoughlin, S. (2001). «История разрыва Гондваны и ее влияние на пре-кузозойский флористический провинциализм». Австралийский журнал ботаники . 49 (3): 271–300. doi : 10.1071/bt00023 .
- ^ Шенгёр, AMC (1987). «Тектоника тетизидов: орогенное развитие коллажа в столкновении». Ежегодный обзор земли и планетарных наук . 15 : 214–244. Bibcode : 1987Areps..15..213c . doi : 10.1146/annurev.ea.15.050187.001241 .
- ^ Jump up to: а беременный Кемп, Т.С. (2006). «Происхождение и раннее излучение рептилий, подобных млекопитающим,: палеобиологическая гипотеза» . Журнал эволюционной биологии . 19 (4): 1231–1247. doi : 10.1111/j.1420-9101.2005.01076.x . PMID 16780524 . S2CID 3184629 .
- ^ Bamford, MK (2016). «Ископаемые леса от верхнего карбона до нижнего бассейна Юрского периода Кару и их интерпретации окружающей среды». В Линоле, б.; де Вит, MJ (ред.). Происхождение и эволюция горов мыса и бассейна Кару . CHAM: региональные обзоры геологии, Springer. С. 159–167. ISBN 978-3-319-40858-3 .
- ^ Ганд, Г.; Завтра, Г.; Крестобал Ф. (1995). «Палихнофауна тетрапод верхней части Эстереля (Прованс, Франция)» Abteilung palaeontography a 4–6 (4–6): 97–139. два 10.1127/pala/235/1995/97: 247392880S2CID
- ^ Ганд, Г.; Гаррик, Дж.; Завтра, Г.; Элленбергер, П. (2000). «Палихнофауна тетрапод верхнего разрешения бассейна Лодева (Лангедок-Франс) » -переживание Палайер 29 (1): 1–82.
- ^ Marchetti, L.; Klein, H.; Бухвиц, М.; Рончи, А.; Смит, RMH; Де Клерк, WJ; Sciscio, L.; Groenewald, GH (2019). «Пермские триасовые следы позвоночных из Южной Африки: Ichnotaxonomy, продюсеры и биостратиграфия через два основных кризиса фауны» . Gondwana Research . 72 : 139–168. Bibcode : 2019gondr..72..139M . doi : 10.1016/j.gr.2019.03.009 . S2CID 133781923 .
- ^ Olroyd, Sl; Сидор, Калифорния (2017). «Обзор Гвадалупийской (средней пермской) глобальной тетрапод ископаемых записей» . Земля-наука обзоров . 171 : 583–597. Bibcode : 2017esrv..171..583o . doi : 10.1016/j.earscirev.2017.07.001 .

