Филотипическая стадия
В эмбриологии или филотипическая стадия филотипический период — это особая стадия развития или период развития в середине эмбриогенеза , когда эмбрионы родственных видов внутри типа проявляют высшую степень морфологического и молекулярного сходства. Недавние молекулярные исследования на различных видах растений и животных позволили количественно оценить экспрессию генов, охватывающих важнейшие стадии развития эмбриона, и обнаружили, что в течение морфологически определенного филотипического периода эволюционно самые старые гены, гены со схожими временными паттернами экспрессии и гены, находящиеся под сильнейшим очищающим отбором. наиболее активны на протяжении всего филотипического периода. [ 1 ]
Историческое происхождение понятия
[ редактировать ]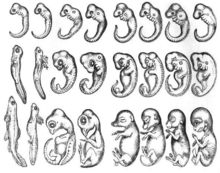

Идея о том, что эмбрионы разных видов на определенном этапе развития имеют схожую морфологию, восходит к Аристотелю . Аристотель наблюдал ряд развивающихся эмбрионов позвоночных, отмечая в своем тексте «Поколение животных» , что морфологические различия между различными эмбрионами возникли на поздних стадиях развития. В 1828 году Карл Эрнст фон Бэр создал свои законы эмбриологии , обобщившие результаты его сравнительных исследований эмбриогенеза . [ 2 ] В своем первом законе он предположил, что более общие признаки группы появляются у эмбрионов раньше, чем более специальные признаки. [ 2 ] В 1866 году Эрнст Геккель предположил, что каждый развивающийся организм проходит эволюционные стадии своих предков, т. е. онтогенез повторяет филогению . [ 3 ] Гипотеза о том, что разные организмы проходят стадии развития близкородственных организмов, устарела. Однако идея о том, что ранние стадии развития сохраняются среди видов, а дивергенция увеличивается по мере развития, оказала влияние на современную эволюционную биологию и биологию развития. [ 4 ] Ранняя консервативная или воронкообразная модель развития (см. ниже) тесно связана с этим историческим происхождением.
Филотипический период
[ редактировать ]Первая формулировка концепции филотипического периода возникла в 1960 году на основе основной формы тела Фридриха Зайделя: [ 5 ] что переводится как «основная форма тела». В 1977 году Коэн определил филетическую стадию как первую стадию, на которой выявляются общие признаки, присущие всем членам этого типа. [ 6 ] Клаус Сандер пересмотрел эту концепцию в 1983 году и назвал ее филотипической стадией. [ 7 ] что является «стадией наибольшего сходства между формами, которые в ходе эволюции по-разному специализировались как в образе взрослой жизни, так и в отношении самых ранних стадий онтогенеза». В недавних работах филотипический период или филотипическая стадия рассматриваются как период максимального сходства между видами внутри каждого типа животных. [ 8 ]
Хотя эта концепция изначально была разработана на основе морфологического сравнения развивающихся эмбрионов разных видов, [ 7 ] период максимального сходства недавно был идентифицирован с помощью молекулярных данных. Филотипический период был идентифицирован с использованием сохранения экспрессии генов, [ 8 ] [ 9 ] оценки возраста генов, [ 10 ] [ 11 ] [ 12 ] сохранение последовательности генов, [ 13 ] экспрессия регуляторных генов и факторов транскрипции, [ 13 ] и взаимосвязь генов и белков. [ 14 ]
Модели воронки и песочных часов
[ редактировать ]Модель воронки — это гипотеза о том, что наиболее консервативная стадия развития (филотипический период) возникает в начале эмбриогенеза , с увеличением дивергенции по мере развития. Это также известно как ранняя консервативная модель развития.
Доказательства альтернативной модели возникли в результате тщательного сравнения временных различий в морфологии эмбрионов разных видов. Например, Клаус Зандер заметил, что «невероятное разнообразие личинок и взрослых особей» насекомых происходит после того, как они «развиваются из почти идентичных зачатков на стадии зародышевого круга». [ 7 ] Наиболее консервативная стадия развития, стадия зародышевого круга, происходит ближе к середине развития, а не в начале, поддерживая средний период развития максимального сходства между видами. Эта модель, называемая моделью песочных часов, [ 15 ] [ 16 ] Это идея о том, что ранние эмбрионы разных видов демонстрируют расходящиеся формы, но их морфологии сходятся в середине развития, после чего наступает период растущей дивергенции.
Поддержка модели песочных часов
[ редактировать ]Вопреки ранним морфологическим работам фон Баера и Геккеля , недавние морфологические исследования продемонстрировали наибольшее расхождение среди близкородственных видов как на ранних стадиях развития ( гаструляция ), так и на поздних стадиях развития. [ 17 ] поддержка модели песочных часов. Дополнительную поддержку модели песочных часов оказало открытие того, что гены Hox , группа последовательно активируемых генов, которые регулируют формирование передне-задней оси тела, активируются в середине развития на филотипической стадии. [ 18 ] Поскольку эти гены высококонсервативны и участвуют в формировании осей тела, активация Hox-генов может играть важную роль в повышении консервативности среди эмбрионов близкородственных видов в середине развития. [ 15 ]
Появление секвенирования нового поколения позволило ученым использовать молекулярные методы для определения периода развития, в котором закономерности экспрессии генов наиболее консервативны среди разных видов. В 2010 году два исследования обнаружили молекулярные доказательства, подтверждающие модель песочных часов. [ 8 ] [ 10 ] Kalinka et al . [ 8 ] секвенировали транскриптом шести видов дрозофил в течение периода развития, выявив наиболее консервативную экспрессию генов в середине развития на стадии развития зародышевых групп членистоногих. Гены, которые были обогащены развивающимися эмбрионами на стадии зародышевой группы, участвуют в развитии клеток и организма. Домазет-Лошо и Таутц [ 10 ] проанализировали транскриптом рыбок данио ( Danio rerio ) в течение периода развития, от неоплодотворенных яиц до взрослых особей. Они использовали метод, называемый геномной филостратиграфией, чтобы оценить возраст каждого гена во время развития. У рыбок данио, а также в дополнительных наборах транскриптомных данных дрозофилы , комара Anopheles и нематоды Caenorhabditis elegans авторы обнаружили, что гены, экспрессируемые в середине развития, старше, чем гены, экспрессируемые в начале и конце развития, что подтверждает модель песочных часов. [ нужна ссылка ]
Другие недавние геномные исследования подтвердили филотипическую стадию среднего развития у позвоночных. [ 9 ] и у растения Arabidopsis thaliana . [ 11 ] [ 12 ] [ 19 ] Временные профили экспрессии генов у развивающейся мыши ( Mus musculus ), курицы ( Gallus Gallus ), лягушки ( Xenopus laevis ) и рыбки данио ( Danio rerio ) показали, что наиболее консервативная экспрессия генов у позвоночных происходит в середине развития на стадии глоточного эмбриона. . Стадия глотки наступает, когда развиваются четыре отличительных признака позвоночных (хорда, дорсальный полый нервный канатик, постанальный хвост и ряд парных жаберных щелей). [ нужна ссылка ]
Поддержка модели раннего сохранения (воронки)
[ редактировать ]Недавние молекулярные данные также подтверждают раннюю модель сохранения. Например, Пясекка и др . [ 13 ] повторно проанализировали набор данных о рыбках данио, опубликованный Домазет-Лошо и Таутцем. [ 10 ] Они обнаружили, что применение логарифмического преобразования к данным об экспрессии генов изменило результаты, подтвердив максимальную консервативность на раннем этапе развития. Кроме того, после кластеризации данных об экспрессии генов рыбок данио в «модули транскрипции», отражающие каждую стадию развития, они обнаружили множество доказательств, подтверждающих модель ранней консервации (последовательность генов, возраст, размер семейства генов и сохранение экспрессии), в то время как только анализ генов регионы регуляции поддержали модель песочных часов. [ 13 ]
Одна из гипотез эволюционного сохранения в течение филотипического периода состоит в том, что это период, характеризующийся высоким уровнем взаимодействия при формировании плана тела. [ 14 ] У рыбок данио взаимосвязь белков во время развития оказалась самой высокой на ранних стадиях развития, что подтверждает модель ранней консервации. [ 14 ] Другой способ изучить тот момент развития, на котором ограничения развития являются наиболее сильными, — это экспериментальная потеря гена, поскольку удаление гена должно быть более вредным, когда оно выражается на стадии развития с более сильными эволюционными ограничениями. [ 20 ] Эксперименты по нокауту генов на мышах и рыбках данио показали, что соотношение незаменимых генов и несущественных генов уменьшается с течением времени развития, что позволяет предположить, что на раннем этапе развития существуют более сильные ограничения, которые со временем ослабляются. [ 20 ] Несмотря на растущее количество доказательств в поддержку модели песочных часов, определение точки развития, которая наиболее консервативна среди видов определенного типа (филотипический период), является спорным вопросом в области биологии развития. [ нужна ссылка ]
Внутритиповый и межтиповый филотипический период
[ редактировать ]Филотипический период определяется как период максимального сходства между видами внутри типа, но недавнее исследование сравнило филотипический период между разными типами, чтобы выяснить, сохраняются ли одни и те же консервативные периоды во время развития в более глубоких филогенетических отношениях. Левин и др . [ 21 ] сравнили закономерности экспрессии генов развития у десяти особей из десяти различных типов животных и нашли доказательства существования модели обратных песочных часов расхождения в экспрессии генов среди разных типов. [ 21 ] Эта модель обратных песочных часов отражает наблюдение о том, что экспрессия генов значительно более различалась между видами на переходном этапе в середине развития, в то время как экспрессия генов была более консервативной на ранних и поздних стадиях развития. [ 21 ] Хотя эта интригующая закономерность может иметь значение для определения типа, [ 21 ] в последующей статье утверждается, что существует несколько методических проблем, которые необходимо решить, чтобы проверить гипотезу о том, что сроки ограничений развития различаются между типами и внутри типа. Во-первых, сравнение одного представителя десяти различных типов может отражать различия между типами, а также более глубокие или более мелкие филогенетические ветви, которые попадают между этими десятью особями, поэтому необходима большая выборка внутри каждого типа. [ 22 ] Во-вторых, при попарных сравнениях каждый из десяти видов рассматривается как независимые наблюдения, но некоторые виды более тесно связаны, чем другие. [ 22 ]
Ссылки
[ редактировать ]- ^ Дрост, Хайк-Георг; Яница, Филипп; Гросс, Иво; Квинт, Марсель (2017). «Сравнение песочных часов развития между королевствами» . Текущее мнение в области генетики и развития . 45 : 69–75. дои : 10.1016/j.где.2017.03.003 . ПМИД 28347942 .
- ^ Jump up to: а б фон Баер, Карл Эрнст (1828). К истории эволюции животных. Наблюдение и размышление . Кенигсберг: Борнтрегер.
- ^ Геккель, Эрнст (1866). Общая морфология организмов . Берлин: Георг Раймер.
- ^ Сандер, Клаус; Шмидт-Отт, Урс (2004). «Эво-Дево-аспекты классических и молекулярных данных в исторической перспективе» . Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 302Б (1): 69–91. дои : 10.1002/jez.b.20003 . ПМИД 14760654 .
- ^ Зайдель, Ф. (1960). «Основная форма тела и зародышевая структура. Обсуждение основ сравнительной и экспериментальной эмбриологии и их обоснованность в филогенетических соображениях». Зоологический вестник . 164 : 245–305.
- ^ Коэн, Дж. (1977). Размножение . Лондон: Баттерворт.
- ^ Jump up to: а б с Сандер, К. (1983). В развитии и эволюции: шестой симпозиум Британского общества биологии развития . Издательство Кембриджского университета.
- ^ Jump up to: а б с д Калинка, Алекс Т.; Варга, Каролина М.; Джеррард, Дэйв Т.; Прейбиш, Стефан; Коркоран, Дэвид Л.; Джарреллс, Джулия; Олер, Уве; Бергман, Кейси М.; Томанчак, Павел (09 декабря 2010 г.). «Расхождение в экспрессии генов повторяет модель песочных часов развития». Природа . 468 (7325): 811–814. Бибкод : 2010Natur.468..811K . дои : 10.1038/nature09634 . ISSN 0028-0836 . ПМИД 21150996 . S2CID 4416340 .
- ^ Jump up to: а б Ирие, Наоки; Куратани, Сигэру (22 марта 2011 г.). «Сравнительный анализ транскриптома выявляет филотипический период позвоночных в ходе органогенеза» . Природные коммуникации . 2 : 248. Бибкод : 2011NatCo...2..248I . дои : 10.1038/ncomms1248 . ISSN 2041-1723 . ПМК 3109953 . ПМИД 21427719 .
- ^ Jump up to: а б с д Домазет-Лошо, Томислав; Таутц, Дитхард (9 декабря 2010 г.). «Филогенетически основанный индекс возраста транскриптома отражает закономерности онтогенетических расхождений». Природа . 468 (7325): 815–818. Бибкод : 2010Natur.468..815D . дои : 10.1038/nature09632 . ISSN 0028-0836 . ПМИД 21150997 . S2CID 1417664 .
- ^ Jump up to: а б Квинт, Марсель; Дрост, Хайк-Георг; Габель, Александр; Ульрих, Кристиан Карстен; Бонн, Маркус; Гроссе, Иво (4 октября 2012 г.). «Транскриптомные песочные часы в эмбриогенезе растений». Природа . 490 (7418): 98-101. Стартовый код : 2012Nature.490...98Q . дои : 10.1038/nature11394 . ISSN 0028-0836 . ПМИД 22951968 . S2CID 4404460 .
- ^ Jump up to: а б Дрост, Хайк-Георг; Габель, Александр; Гросс, Иво; Квинт, Марсель (01 мая 2015 г.). «Доказательства активного поддержания филотранскриптомных паттернов песочных часов в эмбриогенезе животных и растений» . Молекулярная биология и эволюция . 32 (5): 1221–1231. дои : 10.1093/molbev/msv012 . ISSN 0737-4038 . ПМК 4408408 . ПМИД 25631928 .
- ^ Jump up to: а б с д Пясецка, Барбара; Лихоцкий, Павел; Моретти, Себастьян; Бергманн, Свен; Робинсон-Речави, Марк (25 апреля 2013 г.). «Песочные часы и ранние модели сохранения — сосуществующие модели ограничений развития у позвоночных» . ПЛОС Генетика . 9 (4): e1003476. дои : 10.1371/journal.pgen.1003476 . ISSN 1553-7404 . ПМК 3636041 . ПМИД 23637639 .
- ^ Jump up to: а б с Конт, Орели; Ру, Жюльен; Робинсон-Речави, Марк (01 марта 2010 г.). «Молекулярная передача сигналов в развитии рыбок данио и филотипический период позвоночных» . Эволюция и развитие . 12 (2): 144–156. дои : 10.1111/j.1525-142X.2010.00400.x . ISSN 1525-142X . ПМЦ 2855863 . ПМИД 20433455 .
- ^ Jump up to: а б Дюбул, Д. (1 января 1994 г.). «Временная коллинеарность и филотипическая прогрессия: основа стабильности бауплана позвоночных и эволюция морфологии посредством гетерохронии». Разработка. Приложение : 135–142. ПМИД 7579514 .
- ^ Рафф, Рудольф А. (1996). Форма жизни: гены, развитие и эволюция животных форм . Издательство Чикагского университета. ISBN 9780226256573 .
- ^ Шмидт, Кай; Старк, Дж. Матиас (15 сентября 2004 г.). «Изменчивость развития на раннем этапе эмбрионального развития рыбки-зебры Danio rerio». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 302Б (5): 446–457. дои : 10.1002/jez.b.21010 . ISSN 1552-5015 . ПМИД 15580642 .
- ^ Слэк, JMW; Голландия, PWH; Грэм, CF (11 февраля 1993 г.). «Зоотип и филотипический этап». Природа . 361 (6412): 490–492. Бибкод : 1993Natur.361..490S . дои : 10.1038/361490a0 . ПМИД 8094230 . S2CID 4362531 .
- ^ Дрост, Хайк-Георг; Беллстедт, Джулия; О'Маойлеиди, Диармуид С.; Сильва, Андерсон Т.; Габель, Александр; Вайнхольдт, Клаус; Райан, Патрик Т.; Деккерс, Басс Дж.В.; Бенцинк, Леони; Хилхорст, Хенк В.М.; Лайтеринк, Вилко; Веллмер, Фрэнк; Гросс, Иво; Квинт, Марсель (23 февраля 2016 г.). «Постэмбриональные модели песочных часов отмечают онтогенетические переходы в развитии растений» . Молекулярная биология и эволюция . 33 (5): 1158–1163. дои : 10.1093/molbev/msw039 . ПМЦ 4839224 . ПМИД 26912813 .
- ^ Jump up to: а б Ру, Жюльен; Робинсон-Речави, Марк (19 декабря 2008 г.). «Ограничения развития эволюции генома позвоночных» . ПЛОС Генетика . 4 (12): e1000311. дои : 10.1371/journal.pgen.1000311 . ISSN 1553-7404 . ПМК 2600815 . ПМИД 19096706 .
- ^ Jump up to: а б с д Левин, Михал; Анави, Леон; Коул, Элисон Г.; Зима, Эйтан; Мостов, Наталья; Хайр, Салли; Сендерович, Нафтали; Ковалева Екатерина; Сильвер, Дэвид Х. (31 марта 2016 г.). «Переход в середине развития и эволюция строения тела животных» . Природа . 531 (7596): 637–641. Бибкод : 2016Natur.531..637L . дои : 10.1038/nature16994 . ISSN 0028-0836 . ПМЦ 4817236 . ПМИД 26886793 .
- ^ Jump up to: а б Хейнол, Андреас; Данн, Кейси В. (2016). «Эволюция животных: реальны ли типы?» . Современная биология . 26 (10): Р424–Р426. дои : 10.1016/j.cub.2016.03.058 . ПМИД 27218852 .