Премьер-редактирование
Прайм-редактирование «поиска и замены» — это технология редактирования генома в молекулярной биологии, с помощью которой можно модифицировать геном живых организмов. Эта технология напрямую записывает новую генетическую информацию в целевой участок ДНК. Он использует гибридный белок , состоящий из каталитически нарушенной Cas9 эндонуклеазы , слитой с сконструированным ферментом обратной транскриптазы , и основной направляющей РНК редактирования (пегРНК), способной идентифицировать целевой сайт и предоставлять новую генетическую информацию для замены целевых нуклеотидов ДНК. Он обеспечивает целевые вставки , делеции и преобразования оснований без необходимости двунитевых разрывов (DSB) или матриц донорской ДНК. [1]
Эта технология привлекла внимание прессы из-за ее потенциального использования в медицинской генетике. Он использует методологии, аналогичные технологиям редактирования генома-предшественника, включая CRISPR/Cas9 и базовые редакторы . Прайм-редактирование использовалось на некоторых моделях генетических заболеваний на животных. [2] [3] [4] и растения. [5]
Редактирование генома
[ редактировать ]Компоненты
[ редактировать ]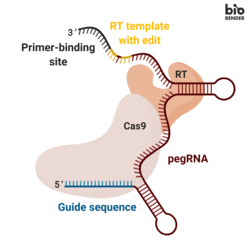
Прайм-редактирование включает в себя три основных компонента: [1]
- Основная направляющая РНК редактирования (пегРНК) , способная (i) идентифицировать целевую нуклеотидную последовательность, подлежащую редактированию, и (ii) кодировать новую генетическую информацию, которая заменяет целевую последовательность. ПегРНК состоит из расширенной одиночной направляющей РНК (сгРНК), содержащей сайт связывания праймера (PBS) и матричную последовательность обратной транскриптазы (RT). Во время редактирования генома сайт связывания праймера позволяет 3'-концу разорванной цепи ДНК гибридизоваться с пегРНК, в то время как матрица RT служит матрицей для синтеза отредактированной генетической информации. [1]
- Гибридный белок, состоящий из никазы Cas9 H840A, слитой с вируса мышиного лейкоза Молони (M-MLV) обратной транскриптазой . [1] [6] [7]
- Никаза Cas9 H840A: фермент Cas9 содержит два домена нуклеазы, которые могут расщеплять последовательности ДНК: домен RuvC, который расщепляет нецелевую цепь, и домен HNH, который расщепляет целевую цепь. Введение замены H840A в Cas9, посредством которой 840-я аминокислота гистидин заменяется аланином, инактивирует домен HNH. Имея только функциональный домен RuvC, каталитически нарушенный Cas9 вводит одноцепочечный разрыв, отсюда и название никаза . [8]
- Обратная транскриптаза M-MLV: фермент, синтезирующий ДНК из матрицы одноцепочечной РНК. [1]
- Одна направляющая РНК (sgRNA), которая направляет часть никазы Cas9 H840A слитого белка на разрыв нередактируемой цепи ДНК. [1]
Механизм
[ редактировать ]
Редактирование генома происходит путем трансфекции клеток пегРНК и гибридного белка. Трансфекцию часто осуществляют путем введения векторов в клетку. После интернализации слитый белок разрывает целевую последовательность ДНК, обнажая 3'-гидроксильную группу, которую можно использовать для инициации (праймирования) обратной транскрипции матричной части RT пегРНК. В результате образуется разветвленный промежуточный продукт, содержащий два лоскута ДНК: 3'-клапан, содержащий вновь синтезированную (отредактированную) последовательность, и 5'-клапан, содержащий ненужную неотредактированную последовательность ДНК. Затем 5'-лоскут расщепляется структурно-специфическими эндонуклеазами или 5'- экзонуклеазами . Этот процесс позволяет лигировать 3'-лоскут и создавать гетеродуплексную ДНК, состоящую из одной отредактированной цепи и одной неотредактированной цепи. Повторно отожженная двухцепочечная ДНК содержит несоответствия нуклеотидов в том месте, где произошло редактирование. Чтобы исправить несоответствия, клетки используют механизм внутренней репарации несоответствий (MMR) с двумя возможными результатами: (i) информация из отредактированной цепи копируется в дополнительную цепь, навсегда устанавливая редактирование; (ii) исходные нуклеотиды повторно включаются в отредактированную цепь, исключая редактирование. [1]
Процесс разработки
[ редактировать ]В ходе разработки этой технологии в компоненты было внесено несколько модификаций с целью повышения ее эффективности. [1]
Главный редактор 1
[ редактировать ]В первой системе обратная транскриптаза вируса мышиного лейкоза Молони (M-MLV) дикого типа была слита с С-концом никазы Cas9 H840A. Наблюдалась заметная эффективность редактирования. [1]
Главный редактор 2
[ редактировать ]Чтобы повысить аффинность ДНК-РНК, процессивность фермента и термостабильность, в обратную транскриптазу M-MLV были включены пять аминокислотных замен. Затем мутантный M-MLV RT был включен в PE1, что дало начало (Cas9 (H840A)-M-MLV RT(D200N/L603W/T330P/T306K/W313F)). Улучшение эффективности наблюдалось по сравнению с PE1. [1]

Главный редактор 3
[ редактировать ]Несмотря на повышенную эффективность, редактирование, внесенное PE2, все равно может быть удалено из-за репарации несоответствия ДНК в отредактированной цепи. Чтобы избежать этой проблемы во время разрешения гетеродуплекса ДНК, вводится дополнительная одна направляющая РНК (sgRNA). Эта sgRNA разработана так, чтобы соответствовать отредактированной последовательности, введенной pegRNA, но не исходному аллелю. Он направляет часть никазы Cas9 слитого белка на разрыв неотредактированной цепи в соседнем участке, противоположном исходному разрыву. Разрезание неотредактированной цепи заставляет естественную систему восстановления клетки копировать информацию из отредактированной цепи в дополнительную цепь, навсегда закрепляя редактирование. [1] Однако у этой системы есть недостатки, поскольку надрезание неизмененной нити может привести к дополнительным нежелательным вставкам . [9]
Главный редактор 4
[ редактировать ]Редактор Prime 4 использует тот же механизм, что и PE2, но также включает плазмиду, которая кодирует доминантно-негативный белок MMR MLH1 . Доминантно-негативный MLH1 способен по существу нокаутировать эндогенный MLH1 путем ингибирования, тем самым снижая клеточный ответ MMR и повышая эффективность первичного редактирования. [9]
Главный редактор 5
[ редактировать ]Prime editor 5 использует тот же механизм, что и PE3, но также включает плазмиду, которая кодирует доминантно-негативный MLH1. Как и PE4, это позволяет подавлять эндогенную реакцию MMR, повышая эффективность первичного редактирования. [9]
Нуклеаза Прайм Редактор
[ редактировать ]Nuclease Prime Editor использует нуклеазу Cas9 вместо никазы Cas9(H840A). В отличие от прайм-редактора 3 (PE3), который требует двойного разрыва на обеих цепях ДНК, чтобы вызвать эффективное редактирование праймов, Nuclease Prime Editor требует только одной пегРНК, поскольку одиночная гРНК уже создает двухцепочечный разрыв вместо одноцепочечного разрыва. [10]
Монтаж Твин Прайм
[ редактировать ]Механизм «редактирования двойных простых чисел» (twinPE), о котором сообщалось в 2021 году, позволяет редактировать большие последовательности ДНК – последовательности размером с гены – что устраняет ключевой недостаток метода. Он использует белок первичного редактора и две направляющие РНК первичного редактирования. [11] [12] [ нужна более подробная информация ]
История
[ редактировать ]Прайм-редактирование было разработано в лаборатории Дэвида Р. Лю в Институте Броуда и раскрыто в Anzalone et al. (2019). [13] С тех пор главное редактирование и исследования, в результате которых оно было получено, получили широкое научное признание. [14] [6] [15] называют «революционным» [7] и важная часть будущего редактирования. [13]
Разработка эпегРНК
[ редактировать ]Эффективность редактирования праймов можно повысить с помощью сконструированных пегРНК (эпегРНК). Одной из распространенных проблем традиционных пегРНК является деградация 3'-конца, что приводит к снижению эффективности PE. epegRNA имеют мотив структурированной РНК, добавленный к их 3'-концу для предотвращения деградации. [16]
Подразумеваемое
[ редактировать ]Хотя для повышения эффективности первичного редактирования необходимы дополнительные исследования, эта технология предлагает многообещающие научные улучшения по сравнению с другими инструментами редактирования генов. Технология первичного редактирования потенциально способна исправить подавляющее большинство патогенных аллелей, вызывающих генетические заболевания, поскольку она может восстанавливать вставки, делеции и нуклеотидные замены. [1]
Преимущества
[ редактировать ]Инструмент первичного редактирования предлагает преимущества по сравнению с традиционными технологиями редактирования генов. Редактирование CRISPR/Cas9 основано на негомологичном соединении концов (NHEJ) или восстановлении, направленном на гомологию (HDR) , для исправления разрывов ДНК, в то время как система первичного редактирования использует восстановление несоответствия ДНК . Это важная особенность этой технологии, учитывая, что механизмы восстановления ДНК, такие как NHEJ и HDR, генерируют нежелательные случайные вставки или делеции (INDEL). Это побочные продукты, которые усложняют извлечение ячеек, несущих правильное редактирование. [1] [17]
Прайм-система вводит одноцепочечные разрывы ДНК вместо двухцепочечных разрывов ДНК, наблюдаемых в других инструментах редактирования, таких как редакторы оснований. В совокупности редактирование оснований и простое редактирование предлагают дополнительные сильные и слабые стороны для создания целевых переходных мутаций. Редакторы оснований обеспечивают более высокую эффективность редактирования и меньшее количество побочных продуктов INDEL, если желаемое редактирование представляет собой мутацию точки перехода и последовательность PAM существует примерно в 15 основаниях от целевого сайта. Однако, поскольку технология первичного редактирования не требует точного расположения последовательности PAM для нацеливания на нуклеотидную последовательность, она обеспечивает большую гибкость и точность редактирования. все типы замен, переходов и трансверсий . Примечательно, что прайм-редакторы позволяют вставлять в целевую последовательность [1] [17] Редактирование оснований цитозина и BE аденина уже могут выполнять точные переходы оснований , но для трансверсий оснований хороших вариантов не было. Прайм-редактирование выполняет трансверсии с хорошим удобством использования. PE может вставлять до 44 пар оснований, удалять до 80 или их комбинации. [7]
Поскольку прайм-система включает три отдельных события связывания ДНК (между (i) направляющей последовательностью и целевой ДНК, (ii) сайтом связывания праймера и целевой ДНК и (iii) 3'-концом разорванной цепи ДНК и pegRNA), предполагается, что он имеет меньше нежелательных побочных эффектов, чем CRISPR/Cas9 . [1] [17]
Ограничения
[ редактировать ]Существует значительный интерес к применению методов редактирования генов для лечения заболеваний с генетическим компонентом. Однако с этим подходом связано множество проблем. Эффективное лечение потребует редактирования большого количества клеток-мишеней, что, в свою очередь, потребует эффективного метода доставки и высокого уровня тканевой специфичности. [1] [18]
По состоянию на 2019 год первичное редактирование выглядит многообещающим для относительно небольших генетических изменений, но необходимо провести дополнительные исследования, чтобы оценить, эффективна ли технология для внесения более крупных изменений, таких как целевые вставки и удаления. Более крупные генетические изменения потребуют более длинной матрицы RT, что может препятствовать эффективной доставке пегРНК к клеткам-мишеням. Более того, пегРНК, содержащая длинную матрицу RT, может стать уязвимой для повреждений, вызванных клеточными ферментами. [1] [18] Первичное редактирование на растениях имеет низкую эффективность, варьирующуюся от нуля до нескольких процентов, и требует значительного улучшения. [19]
Некоторые из этих ограничений были смягчены недавними улучшениями основных редакторов. [2] [20] включая мотивы, которые защищают пегРНК от деградации. [21] Прежде чем можно будет использовать первичное редактирование для коррекции патогенных аллелей у людей, необходимы дальнейшие исследования. [1] [18] Исследования также показали, что ингибирование некоторых белков MMR , включая MLH1, может повысить эффективность первичного редактирования. [9]
Способ доставки
[ редактировать ]Базовые редакторы, используемые для первичного редактирования, требуют доставки молекулы белка и РНК в живые клетки. Внедрение экзогенных технологий редактирования генов в живые организмы является серьезной проблемой. Одним из потенциальных способов введения редактора оснований животным и растениям является упаковка редактора оснований в вирусный капсид. Затем целевой организм может быть трансдуцирован вирусом для синтеза базового редактора in vivo . Распространенные лабораторные векторы трансдукции, такие как лентивирус, вызывают иммунные реакции у людей, поэтому предлагаемые методы лечения людей часто сосредоточены на аденоассоциированном вирусе (ААВ), поскольку инфекции ААВ в основном протекают бессимптомно. К сожалению, эффективная емкость упаковки векторов AAV невелика, примерно 4,4 КБ, не считая инвертированных концевых повторов. [22] Для сравнения: слитый белок SpCas9-обратная транскриптаза имеет размер 6,3 т.п.н. [1] [23] что даже не учитывает удлиненную направляющую РНК, необходимую для нацеливания и праймирования интересующего сайта. Однако успешная доставка на мышей была достигнута за счет разделения редактора на два вектора AAV. [2] [3] [4] [24] или с помощью аденовируса, [3] который имеет большую упаковочную емкость.
Приложения
[ редактировать ]Прайм-редакторы могут использоваться в генных драйвах . Главный редактор может быть включен в Cleaver часть системы Cleave and Rescue / ClvR . В этом случае целью является не точное изменение, а просто разрушение. [25]
ПЭ относится к числу недавно внедренных технологий, которые позволяют передавать однонуклеотидные полиморфизмы (SNP) от одного отдельного сельскохозяйственного растения к другому. PE достаточно точен, чтобы его можно было использовать для воссоздания произвольного SNP в произвольной цели. [14] включая делеции, вставки и все 12-точечные мутации, без необходимости выполнения двухцепочечного разрыва или переноса донорской матрицы. [6]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Анзалоне, Эндрю В.; Рэндольф, Пейтон Б.; Дэвис, Джесси Р.; Соуза, Александр А.; Коблан, Люк В.; Леви, Джонатан М.; Чен, Питер Дж.; Уилсон, Кристофер; Ньюби, Грегори А.; Рагурам, Адитья; Лю, Дэвид Р. (21 октября 2019 г.). «Редактирование генома с поиском и заменой без двухцепочечных разрывов или донорской ДНК» . Природа . 576 (7785): 149–157. Бибкод : 2019Natur.576..149A . дои : 10.1038/s41586-019-1711-4 . ПМК 6907074 . ПМИД 31634902 .
- ^ Jump up to: Перейти обратно: а б с Лю, Пэнпэн; Лян, Шунь-Цин; Чжэн, Чуньвэй; Минцер, Эстер; Чжао, Ян Г.; Понниенсельван, Картикеян; Мир, Аамир; Сонтхаймер, Эрик Дж.; Гао, Гуанпин; Флотт, Теренс Р.; Вулф, Скот А. (9 апреля 2021 г.). «Улучшенные прайм-редакторы позволяют корректировать патогенные аллели и моделировать рак у взрослых мышей» . Природные коммуникации . 12 (1): 2121. Бибкод : 2021NatCo..12.2121L . doi : 10.1038/s41467-021-22295-w . ISSN 2041-1723 . ПМК 8035190 . ПМИД 33837189 .
- ^ Jump up to: Перейти обратно: а б с Бек, Дезире; Ротгангл, Таня; Виллигер, Лукас; Шмидхейни, Лукас; Матис, Николас; Иоанниди, Элеонора; Крейцер, Сюзанна; Контаракис, Захариас; Риманн, Николь; Гриш-Чан, Хиу Ман; Тони, Бит (17 августа 2021 г.). «Лечение метаболического заболевания печени путем первичного редактирования in vivo у мышей» : 2021.08.17.456632. дои : 10.1101/2021.08.17.456632 . S2CID 237218057 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Jump up to: Перейти обратно: а б Чан, Хевон; Джо, Дон Хён; Чо, Чанг Сик; Шин, Чон Хон; Со, Чон Хва; Ю, Гусан; Гопалаппа, Раму; Ким, Даесик; Чо, Сон-Рэ; Ким, Чон Хун; Ким, Хёнбум Генри (26 августа 2021 г.). «Применение прайм-редактирования для коррекции мутаций и фенотипов у взрослых мышей с заболеваниями печени и глаз» . Природная биомедицинская инженерия . 6 (2): 181–194. дои : 10.1038/s41551-021-00788-9 . ISSN 2157-846X . PMID 34446856 . S2CID 237321703 .
- ^ Бисвас, Судип; Бриджленд, Айя; Ирум, Самра; Томсон, Майкл Дж.; Септинингсих, Энданг М. (29 августа 2022 г.). «Оптимизация первичного редактирования в протопластах риса, арахиса, нута и вигны путем восстановления активности GFP» . Международный журнал молекулярных наук . 23 (17): 9809.
- ^ Jump up to: Перейти обратно: а б с Линь, Сюэцю; Чемпарати, Августин; Ла Русса, Мари; Дейли, Тимоти; Ци, Лэй С. (20 июля 2020 г.). «Вычислительные методы анализа крупномасштабных экранов CRISPR» . Ежегодный обзор биомедицинских данных . 3 (1). Годовые обзоры : 137–162. doi : 10.1146/annurev-biodatasci-020520-113523 . ISSN 2574-3414 . S2CID 225570135 .
- ^ Jump up to: Перейти обратно: а б с Чжу, Хаочэн; Ли, Чао; Гао, Цайся (24 сентября 2020 г.). «Применение CRISPR – Cas в сельском хозяйстве и биотехнологии растений». Nature Reviews Молекулярно-клеточная биология . 21 (11). Портфолио природы : 661–677. дои : 10.1038/s41580-020-00288-9 . ISSN 1471-0072 . ПМИД 32973356 . S2CID 221918795 .
- ^ Ран, Ф. Энн; Сюй, Патрик Д.; Лин, Чие-Ю; Гутенберг, Джонатан С.; Конерманн, Сильвана; Тревино, Александро Э.; Скотт, Дэвид А.; Иноуэ, Азуса; Матоба, Сёго; Чжан, И; Чжан, Фэн (сентябрь 2013 г.). «Двойное Никинг с помощью CRISPR Cas9 под управлением РНК для повышения специфичности редактирования генома» . Клетка . 154 (6): 1380–1389. дои : 10.1016/j.cell.2013.08.021 . ПМК 3856256 . ПМИД 23992846 .
- ^ Jump up to: Перейти обратно: а б с д Чен, Питер (28 октября 2021 г.). «Усовершенствованные системы первичного редактирования путем манипулирования клеточными детерминантами результатов редактирования» . Клетка . 184 (22): 5635–5652.e29. дои : 10.1016/j.cell.2021.09.018 . ПМЦ 8584034 . ПМИД 34653350 .
- ^ Адикусума, фетва; Лашингтон, Калеб; Арудкумар, Джейшен; Годахева, Гельшан; Чей, Ю. CJ; Гиерус, Люк; Гейгер, Эшли; Джайн, Ятиш; Рети, Дэниел; Уилсон, Лоуренс О.В.; Бауэр, Денис С; Томас, Пол К. (17 сентября 2021 г.). «Оптимизированное первичное редактирование на основе никазы и нуклеазы в клетках человека и мыши» . Исследования нуклеиновых кислот . 49 (18): 10785–10795. дои : 10.1093/nar/gkab792 . ПМК 8501948 . ПМИД 34534334 .
- ^ Дикорато, Алессандра. «Новая система первичного редактирования вставляет целые гены в клетки человека» . Институт Броуда Массачусетского технологического института . Проверено 16 января 2022 г.
- ^ Анзалоне, Эндрю В.; Гао, Синь Д.; Подрачки, Кристофер Дж.; Нельсон, Эндрю Т.; Коблан, Люк В.; Рагурам, Адитья; Леви, Джонатан М.; Мерсер, Джарон AM; Лю, Дэвид Р. (9 декабря 2021 г.). «Программируемое удаление, замена, интеграция и инверсия больших последовательностей ДНК с редактированием двойных праймов» . Природная биотехнология . 40 (5): 731–740. дои : 10.1038/s41587-021-01133-w . ISSN 1546-1696 . ПМЦ 9117393 . ПМИД 34887556 . S2CID 245012407 .
- ^ Jump up to: Перейти обратно: а б Графф, Грегори Д.; Шерков, Джейкоб С. (31 августа 2020 г.). «Модели трансфера технологий редактирования генома». Ежегодный обзор геномики и генетики человека . 21 (1). Годовые обзоры : 509–534. doi : 10.1146/annurev-genom-121119-100145 . hdl : 2142/110346 . ISSN 1527-8204 . ПМИД 32151165 . S2CID 212652569 .
- ^ Jump up to: Перейти обратно: а б Сойк, Себастьян; Бенуа, Матиас; Липпман, Закари Б. (23 ноября 2020 г.). «Новые горизонты анализа эпистаза в изменении количественных признаков сельскохозяйственных культур». Ежегодный обзор генетики . 54 (1). Годовые обзоры : 287–307. doi : 10.1146/annurev-genet-050720-122916 . ISSN 0066-4197 . ПМИД 32870731 . S2CID 221467135 .
- ^ факультета Мнения Анзалоне, Эндрю В.; Рэндольф, Пейтон Б.; Дэвис, Джесси Р.; Соуза, Александр А.; Коблан, Люк В.; Леви, Джонатан М.; Чен, Питер Дж.; Уилсон, Кристофер; Ньюби, Грегори А.; Рагурам, Адитья; Лю, Дэвид Р. (2019). «Редактирование генома с поиском и заменой без двухцепочечных разрывов или донорской ДНК» . Природа . 576 (7785): 149–157. Бибкод : 2019Natur.576..149A . дои : 10.1038/s41586-019-1711-4 . ПМК 6907074 . ПМИД 31634902 . Проверено 13 ноября 2021 г.
- ^ Нельсон, Джеймс (4 октября 2021 г.). «Сконструированные пегРНК повышают эффективность первичного редактирования» . Природная биотехнология . 40 (432): 402–410. дои : 10.1038/s41587-021-01039-7 . ПМЦ 8930418 . ПМИД 34608327 .
- ^ Jump up to: Перейти обратно: а б с Шеридан, Кормак (7 ноября 2019 г.). «Редактирование генов вступает в прайм-тайм». Природная биотехнология . дои : 10.1038/d41587-019-00032-5 . ПМИД 33154577 . S2CID 209564966 .
- ^ Jump up to: Перейти обратно: а б с «Ученый Дэвид Лю отвечает на ваши вопросы о CRISPR и первичном редактировании» . СТАТ . 06.11.2019 . Проверено 28 февраля 2020 г.
- ^ Молла, Кутубуддин; Сретенович, Симон; Бансал, Кайлаш К.; Ци, Ипин (13 сентября 2021 г.). «Точное редактирование генома растений с использованием базовых и простых редакторов». Природные растения . 7 (9): 1166–1187. дои : 10.1038/s41477-021-00991-1 . ПМИД 34518669 . S2CID 237503774 .
- ^ Чен, Питер Дж.; Хуссманн, Джеффри А.; Ян, Джун; Книппинг, Фридерика; Рависанкар, Пурнима; Чен, Пин-Фан; Чен, Сиди; Нельсон, Джеймс В.; Ньюби, Грегори А.; Шахин, Мустафа; Осборн, Марк Дж. (октябрь 2021 г.). «Усовершенствованные системы первичного редактирования путем манипулирования клеточными детерминантами результатов редактирования» . Клетка . 184 (22): 5635–5652.e29. дои : 10.1016/j.cell.2021.09.018 . ISSN 0092-8674 . ПМЦ 8584034 . ПМИД 34653350 .
- ^ Нельсон, Джеймс В.; Рэндольф, Пейтон Б.; Шен, Саймон П.; Эверетт, Келси А.; Чен, Питер Дж.; Анзалоне, Эндрю В.; Ан, Мейруи; Ньюби, Грегори А.; Чен, Джонатан С.; Сюй, Элвин; Лю, Дэвид Р. (04 октября 2021 г.). «Сконструированные пегРНК повышают эффективность первичного редактирования» . Природная биотехнология . 40 (3): 402–410. дои : 10.1038/s41587-021-01039-7 . ISSN 1546-1696 . ПМЦ 8930418 . ПМИД 34608327 . S2CID 238356160 .
- ^ Ву, Чжицзянь; Ян, Хунъянь; Колози, Питер (2010). «Влияние размера генома на упаковку вектора AAV» . Молекулярная терапия . 18 (1): 80–86. дои : 10.1038/mt.2009.255 . ПМК 2839202 . ПМИД 19904234 .
- ^ «Аддген: PCMV-PE2» .
- ^ Чжи, Шэнъяо, Ву, Гуанлань; Ву, Цзиньни; Ли, Ян, Жуй; Ван, Цзяхуэй; Июль 2021 г. создание системы разделенного прайм-редактора для редактирования генома in vivo» Molecular Therapy . 30 (1): 283–294. doi : /j.ymthe.2021.07.011 . ISSN 1525-0016 . PMC 8753371. . PMID 34298129. 10.1016 S2CID 236212. 353 .
- ^ Хэй, Брюс А.; Оберхофер, Георг; Го, Мин (07 января 2021 г.). «Инженерия состава и судьбы диких популяций с помощью генного драйва» . Ежегодный обзор энтомологии . 66 (1). Годовые обзоры : 407–434. doi : 10.1146/annurev-ento-020117-043154 . ISSN 0066-4170 . ПМИД 33035437 . S2CID 222257628 .