Пресинаптическое торможение
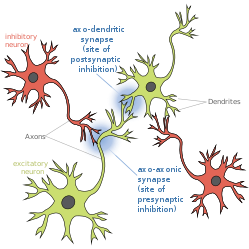
Пресинаптическое торможение — это явление, при котором тормозной нейрон обеспечивает синаптический вход в аксон другого нейрона ( аксо-аксональный синапс ), чтобы снизить вероятность возникновения потенциала действия . Пресинаптическое торможение происходит, когда тормозной нейромедиатор, такой как ГАМК , действует на ГАМК-рецепторы на окончаниях аксонов . Или когда эндоканнабиноиды действуют как ретроградные мессенджеры , связываясь с пресинаптическими рецепторами CB1 , тем самым косвенно модулируя ГАМК и возбудимость дофаминовых нейронов за счет снижения ее и других пресинаптических нейротрансмиттеров . [1] Пресинаптическое торможение широко распространено среди сенсорных нейронов. [2]
Функция
[ редактировать ]Сенсорные стимулы, такие как боль, проприоцепция и соматоощущение , воспринимаются первичными афферентными волокнами. Соматосенсорные нейроны кодируют информацию о текущем состоянии тела (например, температуре, боли, давлении, положении и т. д.). У позвоночных животных эти первичные афферентные волокна образуют синапсы на спинном мозге, особенно в области задних рогов , на различные нижестоящие мишени, включая как возбуждающие нейроны, так и тормозные нейроны. Синапсы между первичными афферентными волокнами и их мишенями представляют собой первую возможность модуляции сенсорной информации. [3] Первичные афферентные волокна содержат множество рецепторов вдоль своих проекций, что делает их поддающимися сложной модуляции. Постоянный приток стимулов окружающей среды, воспринимаемый первичными афферентными волокнами, подлежит модуляции для усиления или ослабления стимулов (см. также: теория контроля ворот контроль усиления и биологический ). Поскольку существует практически неограниченное количество стимулов, крайне важно, чтобы эти сигналы соответствующим образом фильтровались.
Чтобы проверить, подвергается ли соматоощущение, в частности боль, торможению, ученые ввели в спинной мозг грызуна химическое вещество, блокирующее активность первичного тормозного нейромедиатора ( бикукулин , агонист рецептора ГАМК). [4] ). Они обнаружили, что фармакологическая блокировка рецепторов ГАМК на самом деле усиливает восприятие боли; другими словами, ГАМК обычно уменьшает восприятие боли. [5]
Метод, с помощью которого ГАМК модулирует синаптическую передачу от первичных афферентных волокон к их нижестоящим мишеням, оспаривается (см. раздел «Механизмы» ниже). Независимо от механики, ГАМК действует тормозно, снижая вероятность синаптического высвобождения первичного афферентного волокна.
Модулирование первичных афферентных волокон имеет решающее значение для поддержания общего комфорта. Одно исследование показало, что животные без определенного типа рецептора ГАМК на ноцицепторах были гиперчувствительны к боли. [6] тем самым поддерживая функцию пресинаптического торможения как анальгетика. Считается, что некоторые патологические состояния, такие как аллодиния , вызваны немодулированной стимуляцией ноцицепторов . Помимо ослабления боли, нарушение пресинаптического торможения связано со многими неврологическими расстройствами, такими как спастичность после травмы спинного мозга, [7] эпилепсия , аутизм и синдром хрупкой Х-хромосомы . [8] [9] [10] [11] [12]
Механизмы
[ редактировать ]Первичные сенсорные афференты содержат на своих окончаниях рецепторы ГАМК (обзор: [13] Таблица 1). ГАМК-рецепторы представляют собой лиганд-управляемые хлоридные каналы , образованные сборкой пяти субъединиц ГАМК-рецептора . Помимо присутствия ГАМК-рецепторов вдоль сенсорных афферентных аксонов, пресинаптическая терминаль также имеет отчетливый ионный состав с высоким содержанием хлоридов. Это происходит из-за котранспортеров катион-хлорида (например, NKCC1 ), которые поддерживают высокий уровень внутриклеточного хлорида. [14]
Обычно активация ГАМК-рецепторов вызывает приток хлоридов, что гиперполяризует клетку. Однако в первичных афферентных волокнах из-за высокой концентрации хлоридов в пресинаптических окончаниях и, следовательно, изменения их реверсивного потенциала активация рецептора ГАМК фактически приводит к оттоку хлоридов и, как следствие, к деполяризации. Это явление называется первичной афферентной деполяризацией (ПАД). [15] [16] Вызванный ГАМК деполяризационный потенциал афферентных аксонов был продемонстрирован у многих животных, от кошек до насекомых. Интересно, что, несмотря на деполяризованный потенциал, активация рецептора ГАМК вдоль аксона по-прежнему приводит к снижению высвобождения нейромедиатора и, следовательно, по-прежнему является тормозной.
Есть четыре гипотезы, которые предлагают механизмы этого парадокса:
- Деполяризованная мембрана вызывает инактивацию потенциалзависимых натриевых каналов на терминалях и, следовательно, предотвращается распространение потенциала действия. [13] [17] [18]
- Открытые каналы рецепторов ГАМК действуют как шунт , посредством чего ток рассеивается, а не распространяется к терминалам. [13] [17] [18] [19] [20] [21] [22] [23] [24]
- Деполяризованная мембрана вызывает инактивацию потенциалзависимых кальциевых каналов , предотвращая приток кальция в синапс (что необходимо для нейротрансмиссии). [13] [18] [20] [21] [25]
- Деполяризация на терминалях генерирует антидромный спайк (т.е. потенциал действия, генерируемый в аксоне и направляющийся к соме), который предотвращает распространение ортодромных спайков (т.е. потенциала действия, движущегося от сомы клетки к терминалям аксона). [19]
История открытия пресинаптического торможения
[ редактировать ]1933: Грассер и Грэм наблюдали деполяризацию, возникающую в окончаниях сенсорных аксонов. [26]
1938: Барон и Мэтьюз наблюдали деполяризацию, возникающую в окончаниях сенсорных аксонов и вентральном корешке. [27]
1957: Франк и Фуортес ввели термин «пресинаптическое торможение». [28]
1961: Экклс, Экклс и Магни определили, что потенциал дорсального корешка (DRP) возникает в результате деполяризации в окончаниях сенсорных аксонов. [29]
Ссылки
[ редактировать ]- ^ Олесон, Эрик Б. (26 января 2012 г.). «Эндоканнабиноиды формируют аккумбальное кодирование сигнально-мотивированного поведения посредством активации рецептора CB1 в вентральной покрышке» . Нейрон . 73 (2): 360–373. дои : 10.1016/j.neuron.2011.11.018 . ПМК 3269037 . ПМИД 22284189 .
- ^ Макганн JP (июль 2013 г.). «Пресинаптическое торможение обонятельных сенсорных нейронов: новые механизмы и потенциальные функции» . Химические чувства . 38 (6): 459–474. doi : 10.1093/chemse/bjt018 . ПМЦ 3685425 . ПМИД 23761680 .
- ^ Комитато А, Бардони Р. (январь 2021 г.). «Пресинаптическое торможение боли и прикосновения в спинном мозге: от рецепторов к цепям» . Международный журнал молекулярных наук . 22 (1): 414. doi : 10.3390/ijms22010414 . ПМЦ 7795800 . ПМИД 33401784 .
- ^ Манске Р.Х. (сентябрь 1932 г.). «Алкалоиды фумаровых растений: II. Dicentra Cucullaria (L.) Bernh» . Канадский журнал исследований . 7 (3): 265–269. Бибкод : 1932CJRes...7..265M . дои : 10.1139/cjr32-078 . ISSN 1923-4287 .
- ^ Робертс Л.А., Бейер С., Комисарук Б.Р. (ноябрь 1986 г.). «Ноцицептивные реакции на измененную ГАМКергическую активность спинного мозга». Науки о жизни . 39 (18): 1667–74. дои : 10.1016/0024-3205(86)90164-5 . ПМИД 3022091 .
- ^ Прайс TJ, Cervero F, Gold MS, Hammond DL, Prescott SA (апрель 2009 г.). «Регуляция хлоридов в путях боли» . Обзоры исследований мозга . 60 (1): 149–170. дои : 10.1016/j.brainresrev.2008.12.015 . ПМЦ 2903433 . ПМИД 19167425 .
- ^ Карон, Гийом; Бильчак, Ядвига Н.; Коте, Мари-Паскаль (2020). «Прямое доказательство снижения пресинаптического торможения, вызываемого мышечными афферентами PBSt группы I после хронической травмы спинного мозга и восстановления после пошаговой тренировки у крыс» . Журнал физиологии . 598 (20): 4621–4642. дои : 10.1113/JP280070 . ISSN 0022-3751 . ПМЦ 7719595 . ПМИД 32721039 .
- ^ Дейдда Г., Бозарт И.Ф., Канкедда Л. (2014). «Модуляция ГАМКергической передачи при развитии и нарушениях нервного развития: исследование физиологии и патологии для получения терапевтических перспектив» . Границы клеточной нейронауки . 8 : 119. дои : 10.3389/fncel.2014.00119 . ПМК 4033255 . ПМИД 24904277 .
- ^ Зейлхофер Х.У., Вильднер Х., Йевен Г.Е. (январь 2012 г.). «Быстрое синаптическое торможение в спинальной сенсорной обработке и контроле боли» . Физиологические обзоры . 92 (1): 193–235. doi : 10.1152/physrev.00043.2010 . ПМК 3590010 . ПМИД 22298656 .
- ^ Ли Э, Ли Джей, Ким Э (май 2017 г.). «Дисбаланс возбуждения/торможения на животных моделях расстройств аутистического спектра» . Биологическая психиатрия . 81 (10): 838–847. doi : 10.1016/j.biopsych.2016.05.011 . ПМИД 27450033 .
- ^ Д'Хюльст К., Кой РФ (август 2007 г.). «Рецептор ГАМКА: новая мишень для лечения хрупкого Х?». Тенденции в нейронауках . 30 (8): 425–431. дои : 10.1016/j.tins.2007.06.003 . ПМИД 17590448 . S2CID 7340813 .
- ^ Бенаррох Э.Э. (февраль 2007 г.). «Гетерогенность рецепторов ГАМКА, функции и последствия эпилепсии». Неврология . 68 (8): 612–614. дои : 10.1212/01.wnl.0000255669.83468.dd . ПМИД 17310035 . S2CID 11101571 .
- ^ Перейти обратно: а б с д Го Д, Ху Дж (декабрь 2014 г.). «Спинальное пресинаптическое торможение при контроле боли» . Нейронаука . 283 : 95–106. doi : 10.1016/j.neuroscience.2014.09.032 . ПМИД 25255936 .
- ^ Кале К.Т., Стейли К.Дж., Нахед Б.В., Гамба Дж., Хеберт С.С., Лифтон Р.П., Маунт ДБ (сентябрь 2008 г.). «Роль катионно-хлоридных котранспортеров в неврологических заболеваниях». Природная клиническая практика. Неврология . 4 (9): 490–503. дои : 10.1038/ncpneuro0883 . ПМИД 18769373 . S2CID 15424963 .
- ^ Прайс TJ, Cervero F, Gold MS, Hammond DL, Prescott SA (апрель 2009 г.). «Регуляция хлоридов в путях боли» . Обзоры исследований мозга . 60 (1): 149–170. дои : 10.1016/j.brainresrev.2008.12.015 . ПМЦ 2903433 . ПМИД 19167425 .
- ^ Уиллис В.Д. (февраль 1999 г.). «Потенциалы дорсальных корешков и рефлексы дорсальных корешков: палка о двух концах». Экспериментальное исследование мозга . 124 (4): 395–421. дои : 10.1007/s002210050637 . ПМИД 10090653 . S2CID 40738560 .
- ^ Перейти обратно: а б Каттаерт Д., Эль Манира А. (июль 1999 г.). «Шунтирование против инактивации: анализ пресинаптических тормозных механизмов в первичных афферентах раков» . Журнал неврологии . 19 (14): 6079–6089. doi : 10.1523/JNEUROSCI.19-14-06079.1999 . ПМК 6783106 . ПМИД 10407044 .
- ^ Перейти обратно: а б с Уиллис В.Д. (1 февраля 2006 г.). «Исследования Джона Экклза пресинаптического торможения спинного мозга». Прогресс нейробиологии . 78 (3–5): 189–214. doi : 10.1016/j.pneurobio.2006.02.007 . ПМИД 16650518 . S2CID 38669996 .
- ^ Перейти обратно: а б Каттаерт Д., Либерсат Ф., Эль Манира А. (февраль 2001 г.). «Пресинаптическое торможение и антидромные всплески в первичных афферентах раков: вычислительный и экспериментальный анализ» . Журнал неврологии . 21 (3): 1007–1021. doi : 10.1523/JNEUROSCI.21-03-01007.2001 . ПМК 6762302 . ПМИД 11157086 .
- ^ Перейти обратно: а б Панек И., Френч А.С., Зейфарт Э.А., Секизава С., Торккели П.Х. (июль 2002 г.). «Периферическое ГАМКергическое торможение механосенсорных афферентов пауков». Европейский журнал неврологии . 16 (1): 96–104. дои : 10.1046/j.1460-9568.2002.02065.x . ПМИД 12153534 . S2CID 20750558 .
- ^ Перейти обратно: а б Французский AS, Панек I, Torkkeli PH (июнь 2006 г.). «Шунт против инактивации: моделирование ГАМКергического ингибирования в механорецепторах пауков предполагает, что достаточно и того, и другого». Неврологические исследования . 55 (2): 189–196. doi : 10.1016/j.neures.2006.03.002 . ПМИД 16616790 . S2CID 2099107 .
- ^ Миллер Р.Дж. (1998). «Пресинаптические рецепторы». Ежегодный обзор фармакологии и токсикологии . 38 : 201–227. doi : 10.1146/annurev.pharmtox.38.1.201 . ПМИД 9597154 .
- ^ Чжан С.Дж., Джексон М.Б. (март 1995 г.). «Свойства ГАМКА-рецептора нервных окончаний задней доли гипофиза крысы». Журнал нейрофизиологии . 73 (3): 1135–1144. дои : 10.1152/jn.1995.73.3.1135 . ПМИД 7608760 .
- ^ Чжан С.Дж., Джексон М.Б. (март 1995 г.). «Активация рецептора ГАМКА и возбудимость нервных окончаний задней доли гипофиза крысы» . Журнал физиологии . 483 (3): 583–595. doi : 10.1113/jphysicalol.1995.sp020608 . ПМЦ 1157804 . ПМИД 7776245 .
- ^ Грэм Б., Редман С. (февраль 1994 г.). «Моделирование потенциалов действия в синаптических бутонах во время пресинаптического торможения». Журнал нейрофизиологии . 71 (2): 538–549. дои : 10.1152/jn.1994.71.2.538 . ПМИД 8176423 .
- ^ Гассер Х.С., Грэм Х.Т. (январь 1933 г.). «Потенциалы, возникающие в спинном мозге при стимуляции дорсальных корешков». Американский журнал физиологии . 103 (2): 303–320. дои : 10.1152/ajplegacy.1933.103.2.303 .
- ^ Бэррон Д.Х., Мэтьюз Б.Х. (апрель 1938 г.). «Интерпретация потенциальных изменений в спинном мозге» . Журнал физиологии . 92 (3): 276–321. дои : 10.1113/jphysicalol.1938.sp003603 . ПМК 1395290 . ПМИД 16994975 .
- ^ Фрэнк К., Фуортес М.Г. (1957). «Пресинаптическое и постсинаптическое торможение мосинаптических рефлексов». Труды Федерации . 16 :39–40.
- ^ Экклс Дж.К., Экклс Р.М., Магни Ф. (ноябрь 1961 г.). «Центральное тормозящее действие, обусловленное пресинаптической деполяризацией, вызываемой афферентными залпами мышц» . Журнал физиологии . 159 (1): 147–166. doi : 10.1113/jphysicalol.1961.sp006798 . ПМЦ 1359583 . ПМИД 13889050 .